
Overexpression of Both Human Sodium Iodide Symporter (NIS) and BRG1-Bromodomain Synergistically Enhances Radioiodine Sensitivity by Stabilizing p53 through NPM1 Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
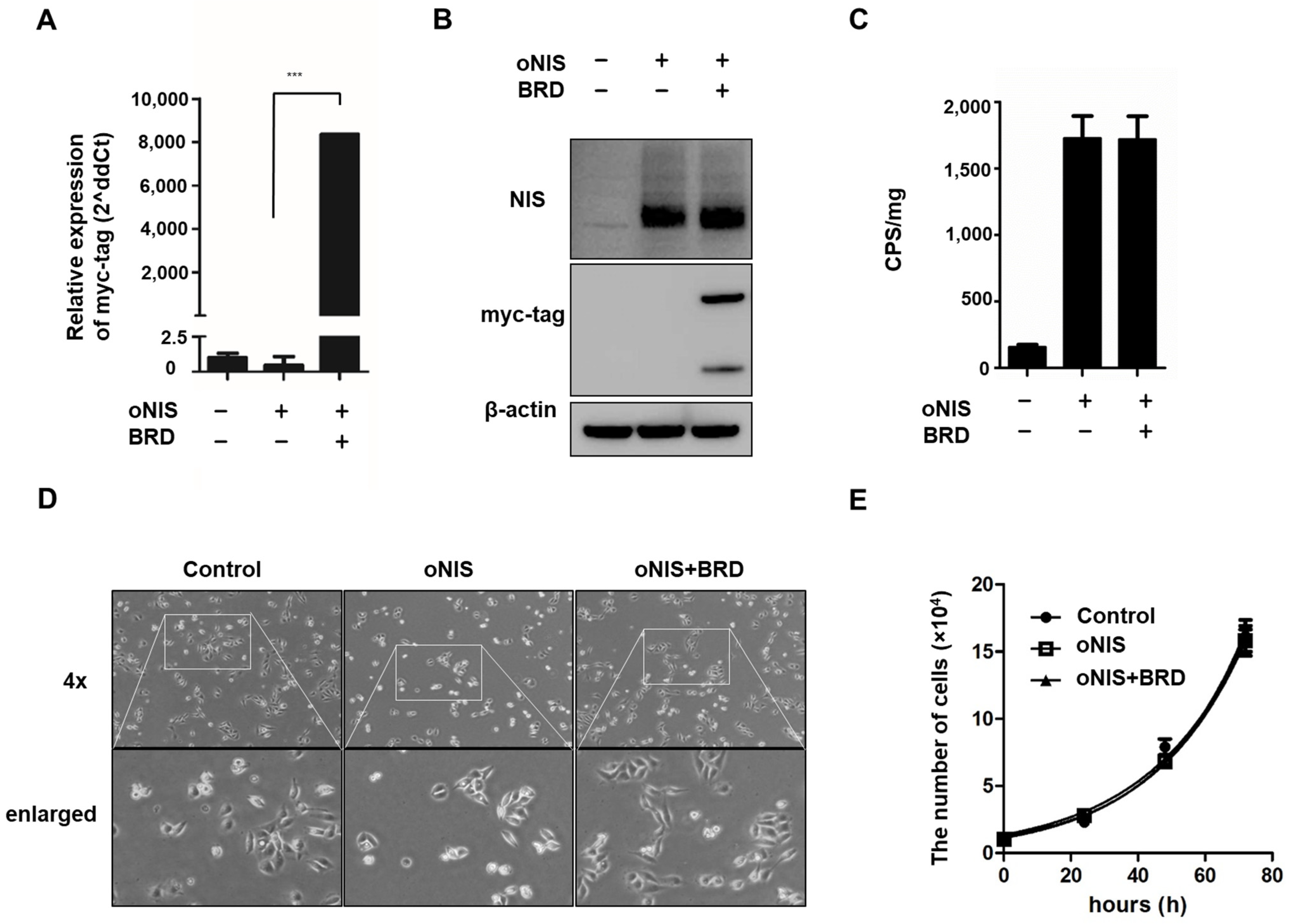
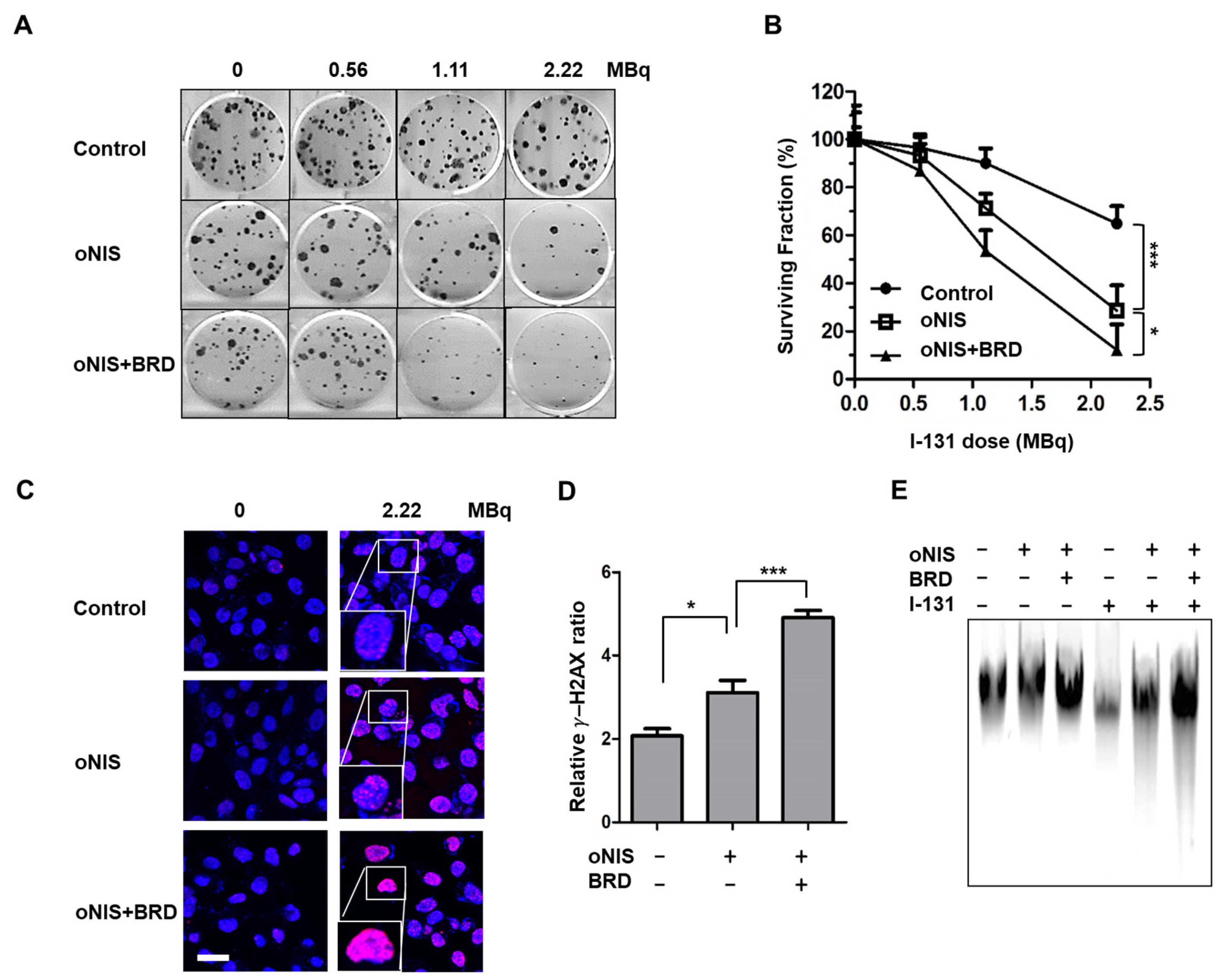
2.1. Overexpression of oNIS and BRG1-BRD Enhances the Effect of I-131 by Increasing DNA DSB Levels
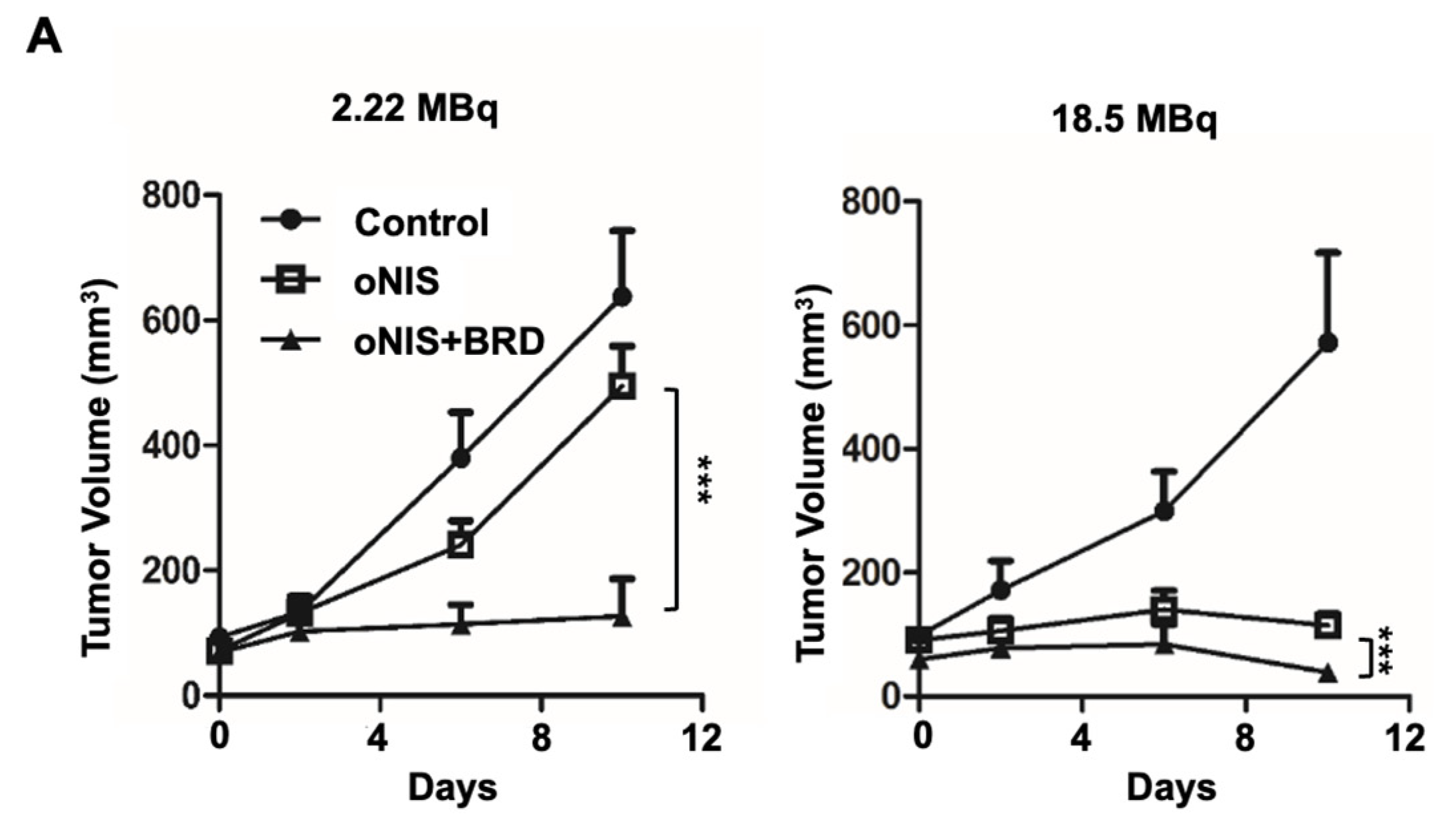
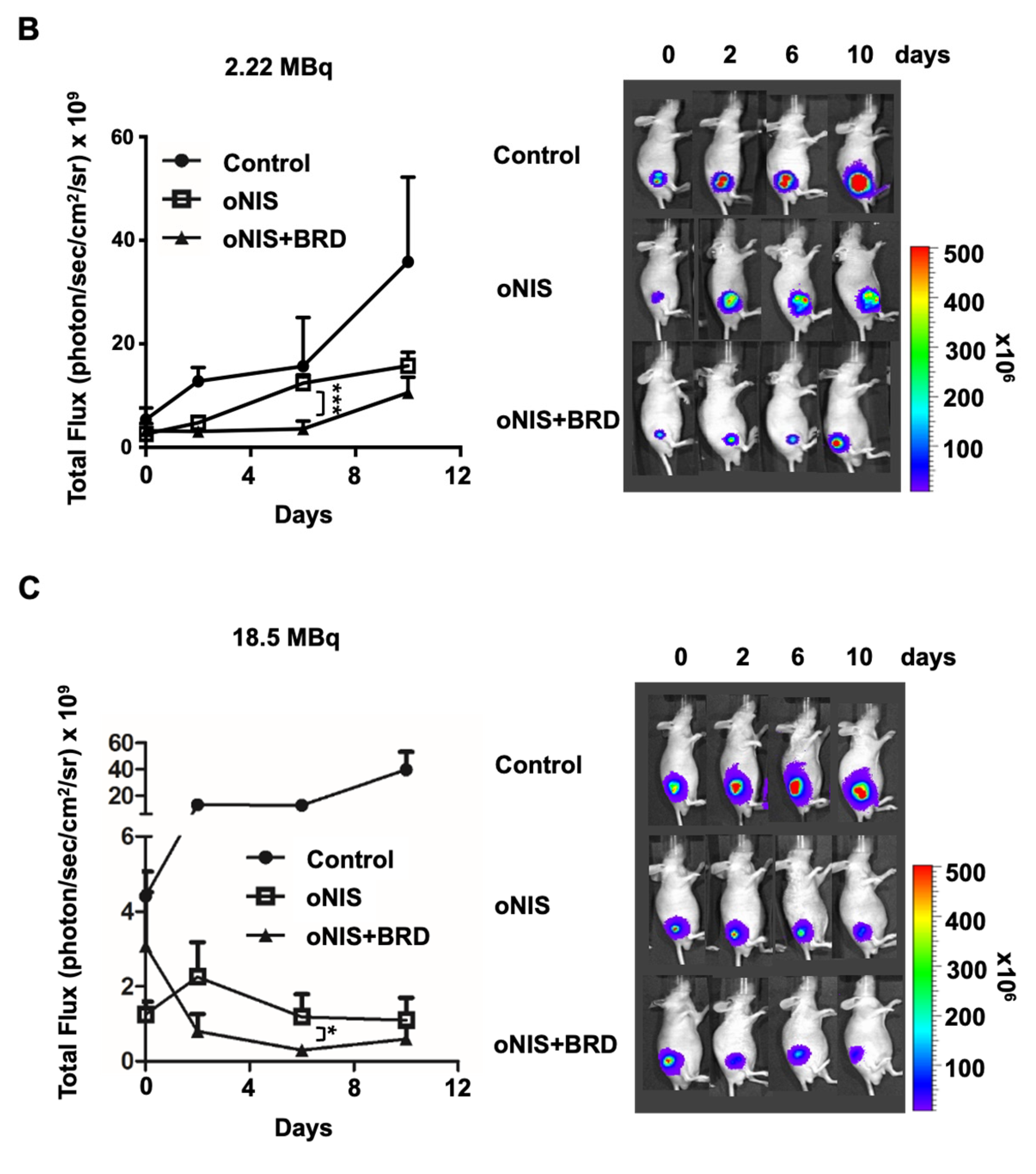
2.2. Overexpression of oNIS and BRG1-BRD Inhibits Tumor Growth In Vivo through I-131-Enhanced Damage
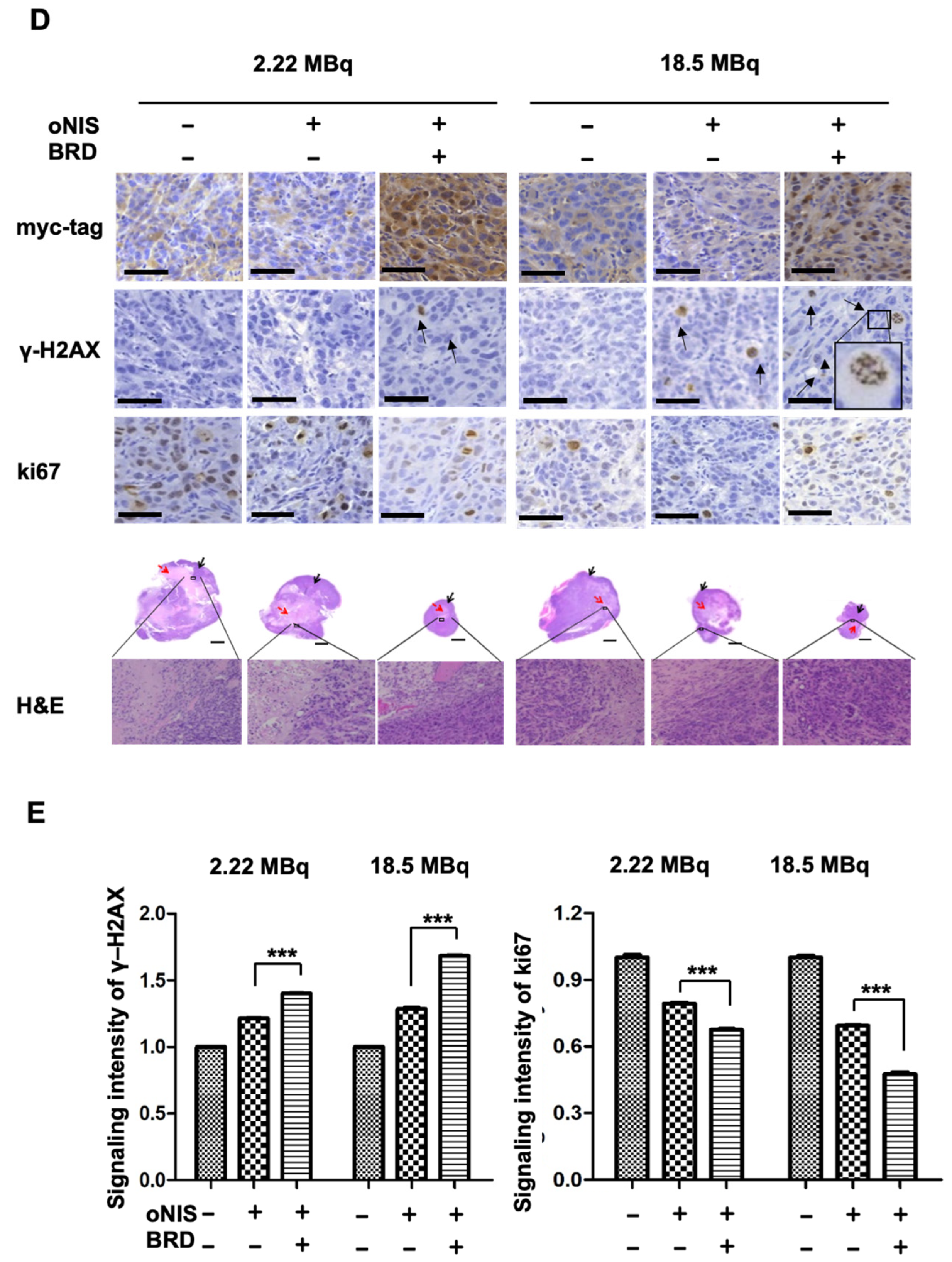
2.3. Immunostained Tumor Sections Show an Increase in DNA DSBs and Fewer Proliferating Cells after I-131 Treatment in the Presence of oNIS+BRD
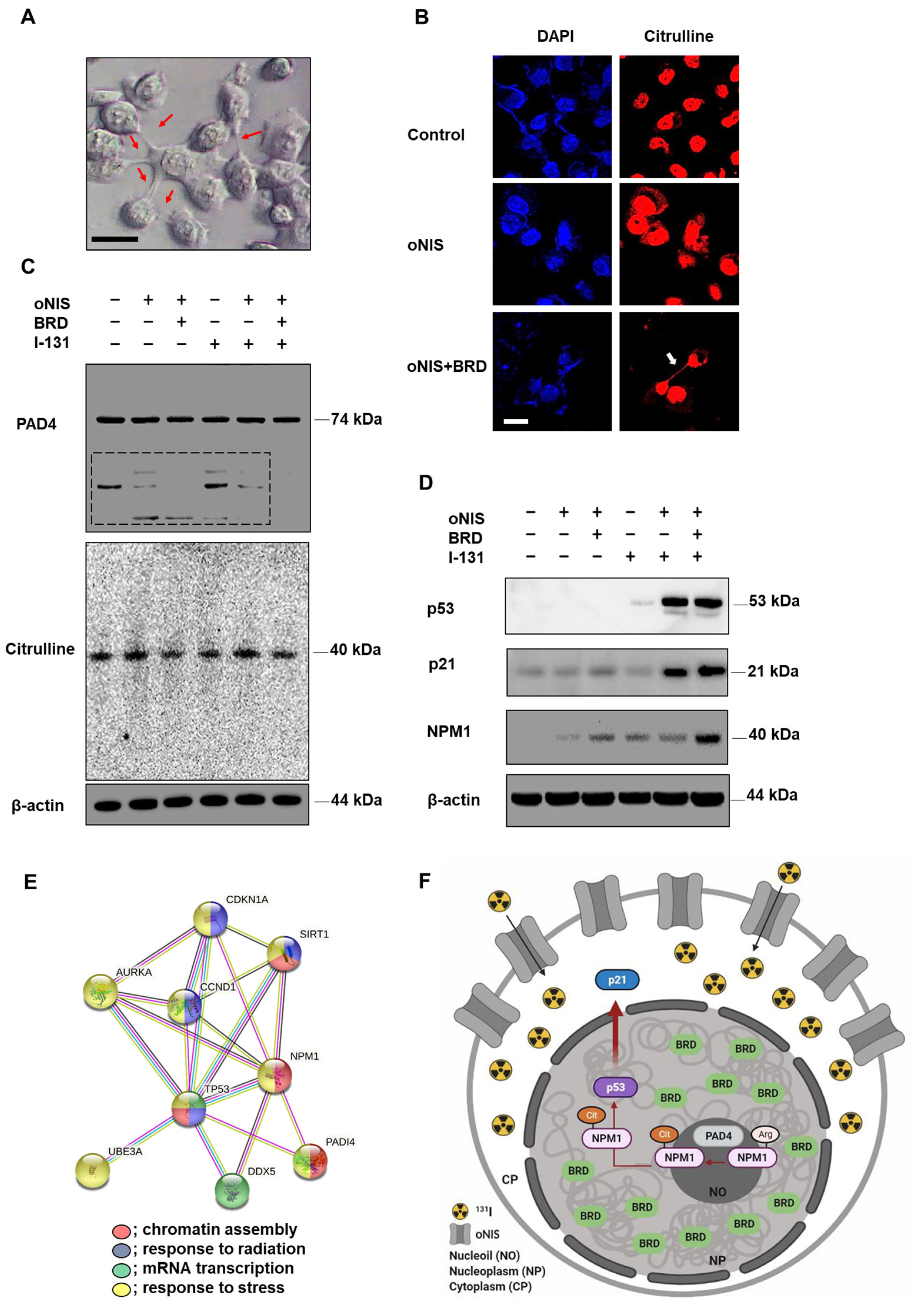
2.4. oNIS and BRG1-BRD Overexpression Induces Citrullination while maintaining the Total Citrulline Level and Increasing p21 and NPM1 Expression after I-131 Treatment
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Retroviral Transduction
4.3. Quantitative Real-Time Reverse Transcription–Polymerase Chain Reaction (Real-Time RT–qPCR)
4.4. Iodine Uptake Assay
4.5. Doubling Time
4.6. Clonogenic Assay
4.7. Immunocytochemistry (ICC) for γ-H2AX Foci and Citrulline Staining
4.8. DNA Fragmentation Assay
4.9. Western Blotting
4.10. Tumor Xenografts in Nude Mice
4.11. Therapeutic Effect of I-131 In Vivo
4.12. Bioluminescence Imaging
4.13. Immunohistochemistry
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, Y. Internal radiation therapy: A neglected aspect of nuclear medicine in the molecular era. J. Biomed. Res. 2015, 29, 345–355. [Google Scholar] [PubMed]
- Erdi, Y.E.; Macapinlac, H.; Larson, S.M.; Erdi, A.K.; Yeung, H.; Furhang, E.E.; Humm, J.L. Radiation Dose Assessment for I-131 Therapy of Thyroid Cancer Using I-124 PET Imaging. Clin. Positron Imaging 1999, 2, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Dehkordi, F.J.; Rasuli, B.; Mahmoud-Pashazadeh, A. An Evaluation of Deviation from the International Atomic Energy Agency-International Commission on Radiological Protection Proposed Equation for Calculation of Radiation Dose Rate Emanating from the Patients with Differentiated Thyroid Cancer Undergoing Radioiodine (I-131) Therapy. World J. Nucl. Med. 2017, 16, 150–155. [Google Scholar] [PubMed]
- Jia, Q.; Meng, Z.; Tan, J.; Zhang, G.; He, Y.; Sun, H.; Upadhyaya, A. Retrospective imaging study on the diagnosis of pathological false positive iodine-131 scans in patients with thyroid cancer. Exp. Ther. Med. 2015, 10, 1995–2001. [Google Scholar] [CrossRef]
- Chung, J.K. Sodium iodide symporter: Its role in nuclear medicine. J. Nucl. Med. 2002, 43, 1188–1200. [Google Scholar]
- Dai, G.; Levy, O.; Carrasco, N. Cloning and characterization of the thyroid iodide transporter. Nature 1996, 379, 458–460. [Google Scholar] [CrossRef]
- Smanik, P.A.; Liu, Q.; Furminger, T.L.; Ryu, K.; Xing, S.; Mazzaferri, E.L.; Jhiang, S.M. Cloning of the human sodium lodide symporter. Biochem. Biophys. Res. Commun. 1996, 226, 339–345. [Google Scholar] [CrossRef]
- Chung, J.K.; Kang, J.H. Translational research using the sodium/iodide symporter in imaging and therapy. Eur. J. Nucl. Med. Mol. Imaging 2004, 31, 799–802. [Google Scholar] [CrossRef]
- Riesco-Eizaguirre, G.; Santisteban, P. A perspective view of sodium iodide symporter research and its clinical implications. Eur. J. Endocrinol. 2006, 155, 495–512. [Google Scholar] [CrossRef]
- Spitzweg, C.; Nelson, P.J.; Wagner, E.; Bartenstein, P.; Weber, W.A.; Schwaiger, M.; Morris, J.C. The sodium iodide symporter (NIS): Novel applications for radionuclide imaging and treatment. Endocr. Relat. Cancer 2021, 28, T193–T213. [Google Scholar] [CrossRef]
- Spitzweg, C.; Dietz, A.B.; O’Connor, M.K.; Bergert, E.R.; Tindall, D.J.; Young, C.Y.; Morris, J.C. In Vivo sodium iodide symporter gene therapy of prostate cancer. Gene Ther. 2001, 8, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Penheiter, A.R.; Russell, S.J.; Carlson, S.K. The sodium iodide symporter (NIS) as an imaging reporter for gene, viral, and cell-based therapies. Curr. Gene Ther. 2012, 12, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Spellerberg, R.; Benli-Hoppe, T.; Kitzberger, C.; Berger, S.; Schmohl, K.A.; Schwenk, N.; Yen, H.Y.; Zach, C.; Schilling, F.; Weber, W.A.; et al. Selective sodium iodide symporter (NIS) genetherapy of glioblastoma mediatedby EGFR-targeted lipopolyplexes. Mol. Ther. Oncolytics 2021, 23, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Youn, H.; Na, J.; Hong, K.J.; Kang, K.W.; Lee, D.S.; Chung, J.K. Codon-optimized human sodium iodide symporter (opt-hNIS) as a sensitive reporter and efficient therapeutic gene. Theranostics 2015, 5, 86–96. [Google Scholar] [CrossRef]
- Rawson, R.W.; Rall, J.E.; Peacock, W. Limitations and indications in the treatment of cancer of the thyroid with radioactive iodine. J. Clin. Endocrinol. Metab. 1951, 11, 1128–1142. [Google Scholar] [CrossRef]
- Mallick, U.; Harmer, C.; Yap, B.; Wadsley, J.; Clarke, S.; Moss, L.; Nicol, A.; Clark, P.M.; Farnell, K.; McCready, R.; et al. Ablation with low-dose radioiodine and thyrotropin alfa in thyroid cancer. N. Engl. J. Med. 2012, 366, 1674–1685. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US); Committee on Thyroid Screening Related to I-131 Exposure; National Research Council (US); Committee on Exposure of the American People to I-131 from the Nevada Atomic Bomb Tests; Commission on Life Sciences; Division on Earth and Life Studies. Exposure of the American People to Iodine-131 from Nevada Nuclear-Bomb Tests: Review of the National Cancer Institute Report and Public Health Implications; National Academy of Sciences (US): Washington, DC, USA, 1999. [Google Scholar]
- Muschel, R.J.; Soto, D.E.; McKenna, W.G.; Bernhard, E.J. Radiosensitization and apoptosis. Oncogene 1998, 17, 3359–3363. [Google Scholar] [CrossRef]
- Willers, H.; Azzoli, C.G.; Santivasi, W.L.; Xia, F. Basic mechanisms of therapeutic resistance to radiation and chemotherapy in lung cancer. Cancer J. 2013, 19, 200–207. [Google Scholar] [CrossRef]
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to improve radiotherapy with targeted drugs. Nat. Rev. Cancer 2011, 11, 239–253. [Google Scholar] [CrossRef]
- Lobrich, M.; Jeggo, P.A. The impact of a negligent G2/M checkpoint on genomic instability and cancer induction. Nat. Rev. Cancer 2007, 7, 861–869. [Google Scholar] [CrossRef]
- Lewanski, C.R.; Gullick, W.J. Radiotherapy and cellular signalling. Lancet Oncol. 2001, 2, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Price, B.D.; D’Andrea, A.D. Chromatin remodeling at DNA double-strand breaks. Cell 2013, 152, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Allard, S.; Masson, J.Y.; Cote, J. Chromatin remodeling and the maintenance of genome integrity. Biochim. Biophys. Acta 2004, 1677, 158–164. [Google Scholar] [CrossRef] [PubMed]
- van Attikum, H.; Gasser, S.M. Crosstalk between histone modifications during the DNA damage response. Trends Cell Biol. 2009, 19, 207–217. [Google Scholar] [CrossRef]
- Reisman, D.N.; Sciarrotta, J.; Bouldin, T.W.; Weissman, B.E.; Funkhouser, W.K. The expression of the SWI/SNF ATPase subunits BRG1 and BRM in normal human tissues. Appl. Immunohistochem. Mol. Morphol. 2005, 13, 66–74. [Google Scholar] [CrossRef]
- Morrison, E.A.; Sanchez, J.C.; Ronan, J.L.; Farrell, D.P.; Varzavand, K.; Johnson, J.K.; Gu, B.X.; Crabtree, G.R.; Musselman, C.A. DNA binding drives the association of BRG1/hBRM bromodomains with nucleosomes. Nat. Commun. 2017, 8, 16080. [Google Scholar] [CrossRef]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. GammaH2AX and cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Park, J.H.; Kim, S.J.; Kwon, S.J.; Kwon, J. A cooperative activation loop among SWI/SNF, gamma-H2AX and H3 acetylation for DNA double-strand break repair. EMBO J. 2010, 29, 1434–1445. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lee, S.K.; Na, J.; Lee, S.A.; Lee, H.S.; Park, J.H.; Chung, J.K.; Youn, H.; Kwon, J. Targeting BRG1 chromatin remodeler via its bromodomain for enhanced tumor cell radiosensitivity in vitro and in vivo. Mol. Cancer Ther. 2015, 14, 597–607. [Google Scholar] [CrossRef]
- Byrd, A.S.; O’Brien, X.M.; Johnson, C.M.; Lavigne, L.M.; Reichner, J.S. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to Candida albicans. J. Immunol. 2013, 190, 4136–4148. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canesi, L. Conservation of Cell Communication Systems in Invertebrate Host-Defence Mechanisms: Possible Role in Immunity and Disease. Biology 2020, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5, 178ra40. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef]
- Ou, Q.; Fang, J.Q.; Zhang, Z.S.; Chi, Z.; Fang, J.; Xu, D.Y.; Lu, K.Z.; Qian, M.Q.; Zhang, D.Y.; Guo, J.P.; et al. TcpC inhibits neutrophil extracellular trap formation by enhancing ubiquitination mediated degradation of peptidylarginine deiminase 4. Nat. Commun. 2021, 12, 3481. [Google Scholar] [CrossRef]
- Fragkos, M.; Jurvansuu, J.; Beard, P. H2AX is required for cell cycle arrest via the p53/p21 pathway. Mol. Cell Biol. 2009, 29, 2828–2840. [Google Scholar] [CrossRef]
- Tanikawa, C.; Ueda, K.; Nakagawa, H.; Yoshida, N.; Nakamura, Y.; Matsuda, K. Regulation of protein Citrullination through p53/PADI4 network in DNA damage response. Cancer Res. 2009, 69, 8761–8769. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, L.; Zhao, P.; Li, T. Role of citrullination modification catalyzed by peptidylarginine deiminase 4 in gene transcriptional regulation. Acta Biochim. Biophys. Sin. 2017, 49, 567–572. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Mumtaz, M.; Lin, L.S.; Hui, K.C.; Mohd Khir, A.S. Radioiodine I-131 for the therapy of graves’ disease. Malays. J. Med. Sci. 2009, 16, 25–33. [Google Scholar] [PubMed]
- Namwongprom, S.; Dejkhamron, P.; Unachak, K. Success rate of radioactive iodine treatment for children and adolescent with hyperthyroidism. J. Endocrinol. Investig. 2021, 44, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Obici, S.; Yaqub, A. Radioiodine Therapy-Induced Conversion of Toxic Adenoma to Graves’ Disease. Cureus 2020, 12, e8683. [Google Scholar] [CrossRef] [PubMed]
- Iwano, S.; Ito, S.; Kamiya, S.; Ito, R.; Kato, K.; Naganawa, S. Unexpected radioactive iodine accumulation on whole-body scan after I-131 ablation therapy for differentiated thyroid cancer. Nagoya J. Med. Sci. 2020, 82, 205–215. [Google Scholar]
- Shi, S.; Zhang, M.; Guo, R.; Zhang, M.; Hu, J.; Xi, Y.; Miao, Y.; Li, B. 131I therapy mediated by sodium/iodide symporter combined with kringle 5 has a synergistic therapeutic effect on glioma. Oncol. Rep. 2016, 35, 691–698. [Google Scholar] [CrossRef]
- Kim, S.; Youn, H.; Song, M.G.; Kang, J.H.; Chung, H.K.; Lee, D.S.; Chung, J.K. Complementary treatment of siTERT for improving the antitumor effect of TERT-specific I-131 therapy. Cancer Gene Ther. 2012, 19, 263–270. [Google Scholar] [CrossRef]
- Jimenez, C.; Erwin, W.; Chasen, B. Targeted Radionuclide Therapy for Patients with Metastatic Pheochromocytoma and Paraganglioma: From Low-Specific-Activity to High-Specific-Activity Iodine-131 Metaiodobenzylguanidine. Cancers 2019, 11, 1018. [Google Scholar] [CrossRef]
- Hahner, S.; Kreissl, M.C.; Fassnacht, M.; Haenscheid, H.; Knoedler, P.; Lang, K.; Buck, A.K.; Reiners, C.; Allolio, B.; Schirbel, A. [131I]iodometomidate for targeted radionuclide therapy of advanced adrenocortical carcinoma. J. Clin. Endocrinol. Metab. 2012, 97, 914–922. [Google Scholar] [CrossRef]
- Klutz, K.; Willhauck, M.J.; Wunderlich, N.; Zach, C.; Anton, M.; Senekowitsch-Schmidtke, R.; Goke, B.; Spitzweg, C. Sodium iodide symporter (NIS)-mediated radionuclide ((131)I, (188)Re) therapy of liver cancer after transcriptionally targeted intratumoral in vivo NIS gene delivery. Hum. Gene Ther. 2011, 22, 1403–1412. [Google Scholar] [CrossRef]
- Ersahin, D.; Doddamane, I.; Cheng, D. Targeted radionuclide therapy. Cancers 2011, 3, 3838–3855. [Google Scholar] [CrossRef]
- Gardette, M.; Papon, J.; Bonnet, M.; Desbois, N.; Labarre, P.; Wu, T.D.; Miot-Noirault, E.; Madelmont, J.C.; Guerquin-Kern, J.L.; Chezal, J.M.; et al. Evaluation of new iodinated acridine derivatives for targeted radionuclide therapy of melanoma using 125I, an Auger electron emitter. Investig. New Drugs 2011, 29, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.S. Radioiodine therapy for hyperthyroidism. N. Engl. J. Med. 2011, 364, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Pant, G.S.; Sharma, S.K.; Bal, C.S.; Kumar, R.; Rath, G.K. Radiation dose to family members of hyperthyroidism and thyroid cancer patients treated with 131I. Radiat. Prot. Dosim. 2006, 118, 22–27. [Google Scholar] [CrossRef]
- Aashiq, M.; Silverman, D.A.; Náara, S.; Takahashi, H.; Amit, M. Radioiodine-Refractory Thyroid Cancer: Molecular Basis of Redifferentiation Therapies, Management, and Novel Therapies. Cancers 2019, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, N.; Jeyasekharan, A.D.; Bernal, J.A.; Venkitaraman, A.R. HP1-beta mobilization promotes chromatin changes that initiate the DNA damage response. Nature 2008, 453, 682–686. [Google Scholar] [CrossRef]
- Santos Apolonio, J.; Lima de Souza Goncalves, V.; Cordeiro Santos, M.L.; Silva Luz, M.; Silva Souza, J.V.; Rocha Pinheiro, S.L.; de Souza, W.R.; Sande Loureiro, M.; de Melo, F.F. Oncolytic virus therapy in cancer: A current review. World J. Virol. 2021, 10, 229–255. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef]
- Dhalluin, C.; Carlson, J.E.; Zeng, L.; He, C.; Aggarwal, A.K.; Zhou, M.M. Structure and ligand of a histone acetyltransferase bromodomain. Nature 1999, 399, 491–496. [Google Scholar] [CrossRef]
- Li, Y.; Liu, B.; Fukudome, E.Y.; Lu, J.; Chong, W.; Jin, G.; Liu, Z.; Velmahos, G.C.; Demoya, M.; King, D.R.; et al. Identification of citrullinated histone H3 as a potential serum protein biomarker in a lethal model of lipopolysaccharide-induced shock. Surgery 2011, 150, 442–451. [Google Scholar] [CrossRef]
- Zhao, J.; Jiang, P.; Guo, S.; Schrodi, S.J.; He, D. Apoptosis, Autophagy, NETosis, Necroptosis, and Pyroptosis Mediated Programmed Cell Death as Targets for Innovative Therapy in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 809806. [Google Scholar] [CrossRef]
- Hamam, H.J.; Palaniyar, N. Histone Deacetylase Inhibitors Dose-Dependently Switch Neutrophil Death from NETosis to Apoptosis. Biomolecules 2019, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, L.; Chang, N.; Hou, L.; Zhou, X.; Yang, L.; Li, L. Neutrophils undergo switch of apoptosis to NETosis during murine fatty liver injury via S1P receptor 2 signaling. Cell Death Discov. 2020, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Azzouz, D.; Khan, M.A.; Sweezey, N.; Palaniyar, N. Two-in-one: UV radiation simultaneously induces apoptosis and NETosis. Cell Death Discov. 2018, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- Azzouz, D.; Palaniyar, N. ApoNETosis: Discovery of a novel form of neutrophil death with concomitant apoptosis and NETosis. Cell Death Discov. 2018, 9, 839. [Google Scholar] [CrossRef]
- Leshner, M.; Wang, S.; Lewis, C.; Zheng, H.; Chen, X.A.; Santy, L.; Wang, Y. PAD4 mediated histone hypercitrullination induces heterochromatin decondensation and chromatin unfolding to form neutrophil extracellular trap-like structures. Front. Immunol. 2012, 3, 307. [Google Scholar] [CrossRef]
- Li, P.; Wang, D.; Yao, H.; Doret, P.; Hao, G.; Shen, Q.; Qiu, H.; Zhang, X.; Wang, Y.; Chen, G.; et al. Coordination of PAD4 and HDAC2 in the regulation of p53-target gene expression. Oncogene 2010, 29, 3153–3162. [Google Scholar] [CrossRef]
- Hagiwara, T.; Nakashima, K.; Hirano, H.; Senshu, T.; Yamada, M. Deimination of arginine residues in nucleophosmin/B23 and histones in HL-60 granulocytes. Biochem. Biophys. Res. Commun. 2002, 290, 979–983. [Google Scholar] [CrossRef]
- Poletto, M.; Lirussi, L.; Wilson, D.M., 3rd; Tell, G. Nucleophosmin modulates stability, activity, and nucleolar accumulation of base excision repair proteins. Mol. Biol. Cell 2014, 25, 1641–1652. [Google Scholar] [CrossRef]
- Xu, R.; Yu, S.; Zhu, D.; Huang, X.; Xu, Y.; Lao, Y.; Tian, Y.; Zhang, J.; Tang, Z.; Zhang, Z.; et al. hCINAP regulates the DNA-damage response and mediates the resistance of acute myelocytic leukemia cells to therapy. Nat. Commun. 2019, 10, 3812. [Google Scholar] [CrossRef]
- Ziv, O.; Zeisel, A.; Mirlas-Neisberg, N.; Swain, U.; Nevo, R.; Ben-Chetrit, N.; Martelli, M.P.; Rossi, R.; Schiesser, S.; Canman, C.E.; et al. Identification of novel DNA-damage tolerance genes reveals regulation of translesion DNA synthesis by nucleophosmin. Nat. Commun. 2014, 5, 5437. [Google Scholar] [CrossRef]
- Traver, G.; Sekhar, K.R.; Crooks, P.A.; Keeney, D.S.; Freeman, M.L. Targeting NPM1 in irradiated cells inhibits NPM1 binding to RAD51, RAD51 foci formation and radiosensitizes NSCLC. Cancer Lett. 2021, 500, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, N.; Gieringer, R.; Grus, F.; Brieger, J. Phosphoproteome Profiling Reveals Multifunctional Protein NPM1 as part of the Irradiation Response of Tumor Cells. Transl. Oncol. 2019, 12, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, V.; Kishore, A.H.; Febitha, K.K.; Kundu, T.K. Human histone chaperone nucleophosmin enhances acetylation-dependent chromatin transcription. Mol. Cell Biol. 2005, 25, 7534–7545. [Google Scholar] [CrossRef] [PubMed]
- Tamada, H.; Van Thuan, N.; Reed, P.; Nelson, D.; Katoku-Kikyo, N.; Wudel, J.; Wakayama, T.; Kikyo, N. Chromatin decondensation and nuclear reprogramming by nucleoplasmin. Mol. Cell Biol. 2006, 26, 1259–1271. [Google Scholar] [CrossRef]
- Sung, Y.K.; Kim, S.W. Recent advances in the development of gene delivery systems. Biomater. Res. 2019, 23, 8. [Google Scholar] [CrossRef]
- Institute of Laboratory Animal Resources (US); Committee on Care, Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals; US Department of Health and Human Services, Public Health Service, National Institutes of Health: Washington, DC, USA, 1986.






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Na, J.; Lee, C.-H.; Chung, J.-K.; Youn, H. Overexpression of Both Human Sodium Iodide Symporter (NIS) and BRG1-Bromodomain Synergistically Enhances Radioiodine Sensitivity by Stabilizing p53 through NPM1 Expression. Int. J. Mol. Sci. 2023, 24, 2761. https://doi.org/10.3390/ijms24032761
Na J, Lee C-H, Chung J-K, Youn H. Overexpression of Both Human Sodium Iodide Symporter (NIS) and BRG1-Bromodomain Synergistically Enhances Radioiodine Sensitivity by Stabilizing p53 through NPM1 Expression. International Journal of Molecular Sciences. 2023; 24(3):2761. https://doi.org/10.3390/ijms24032761
Chicago/Turabian StyleNa, Juri, Chul-Hee Lee, June-Key Chung, and Hyewon Youn. 2023. "Overexpression of Both Human Sodium Iodide Symporter (NIS) and BRG1-Bromodomain Synergistically Enhances Radioiodine Sensitivity by Stabilizing p53 through NPM1 Expression" International Journal of Molecular Sciences 24, no. 3: 2761. https://doi.org/10.3390/ijms24032761
APA StyleNa, J., Lee, C.-H., Chung, J.-K., & Youn, H. (2023). Overexpression of Both Human Sodium Iodide Symporter (NIS) and BRG1-Bromodomain Synergistically Enhances Radioiodine Sensitivity by Stabilizing p53 through NPM1 Expression. International Journal of Molecular Sciences, 24(3), 2761. https://doi.org/10.3390/ijms24032761