
Melatonin Mitigates iNOS-Related Effects of HEMA and Camphorquinone in Human Dental Pulp Cells: Relevance for Postoperative Sensitivity Mechanism in Type 2 Diabetes

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
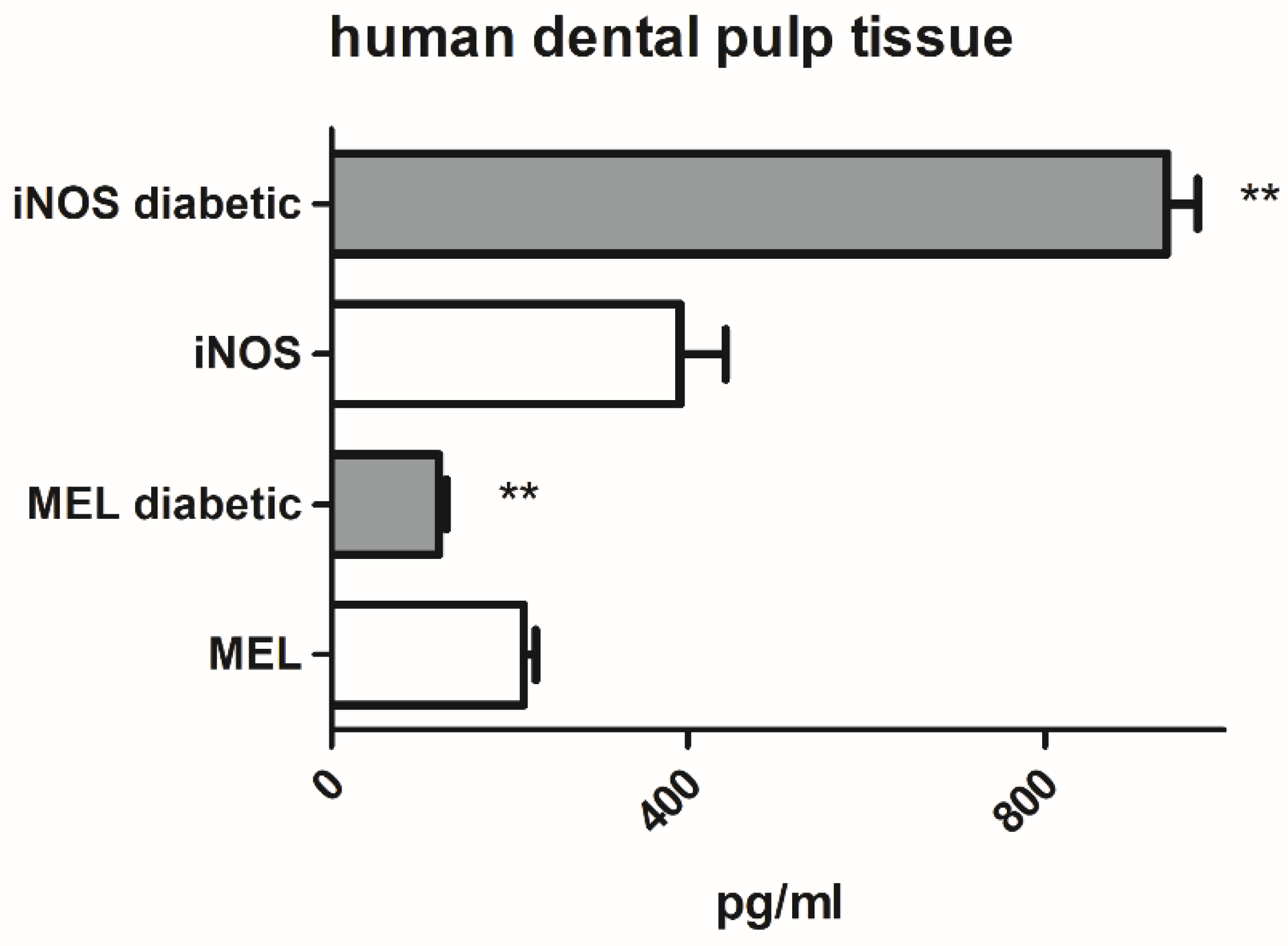
2.1. Melatonin and iNOS Content in Dental Pulp Tissue from Non-Diabetic and Type 2 Diabetic Patients and Incidence of Postoperative Sensitivity after Placement of Direct Composite Restorations in Type 2 Diabetic Patients
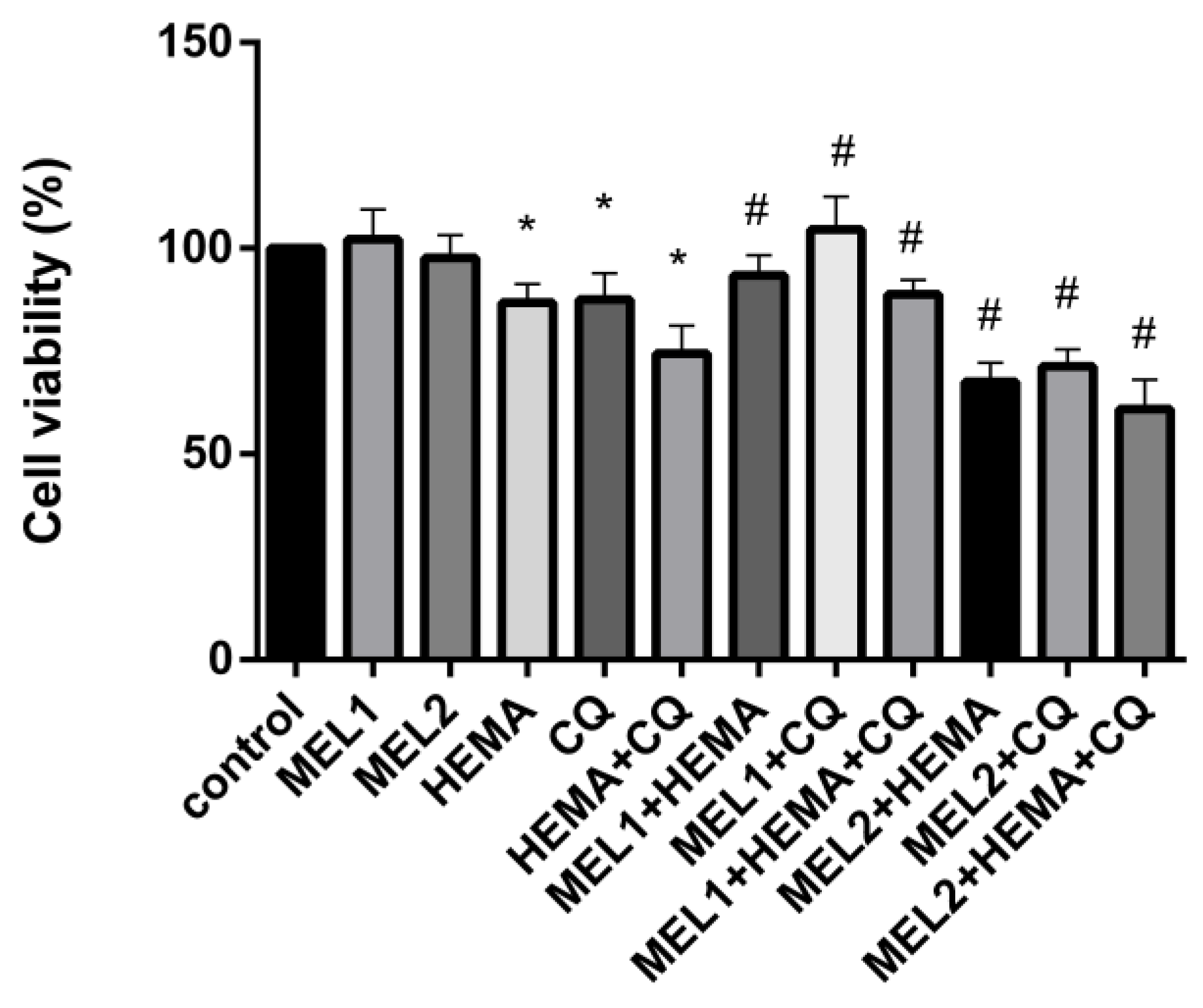
2.2. Melatonin Effects on Viability of hDPCs Treated with HEMA and/or CQ
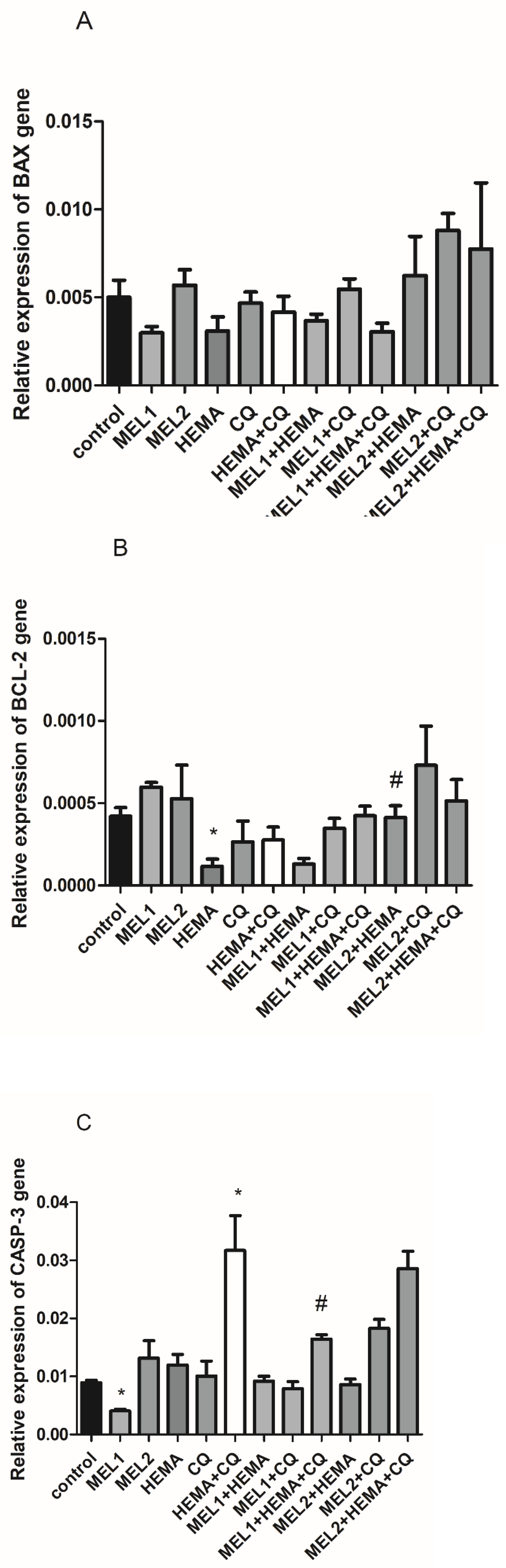
2.3. Melatonin Effects on BAX, BCL-2, and CASP3Gene Expression Levels in hDPCs Treated with HEMA and/or CQ
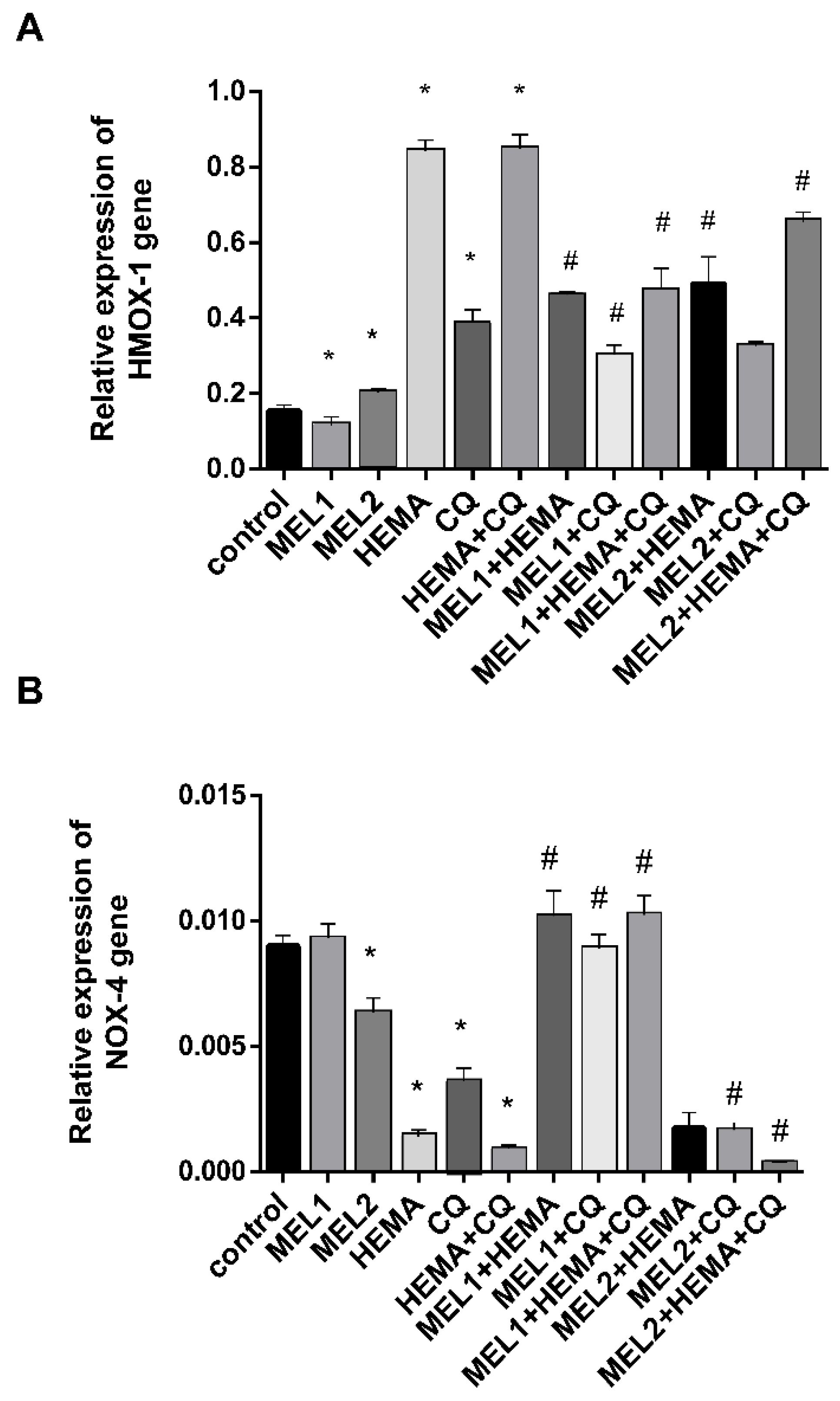
2.4. Melatonin Effects on HMOX1 and NOX4 Gene Expression Levels in hDPCs Treated with HEMA and/or CQ
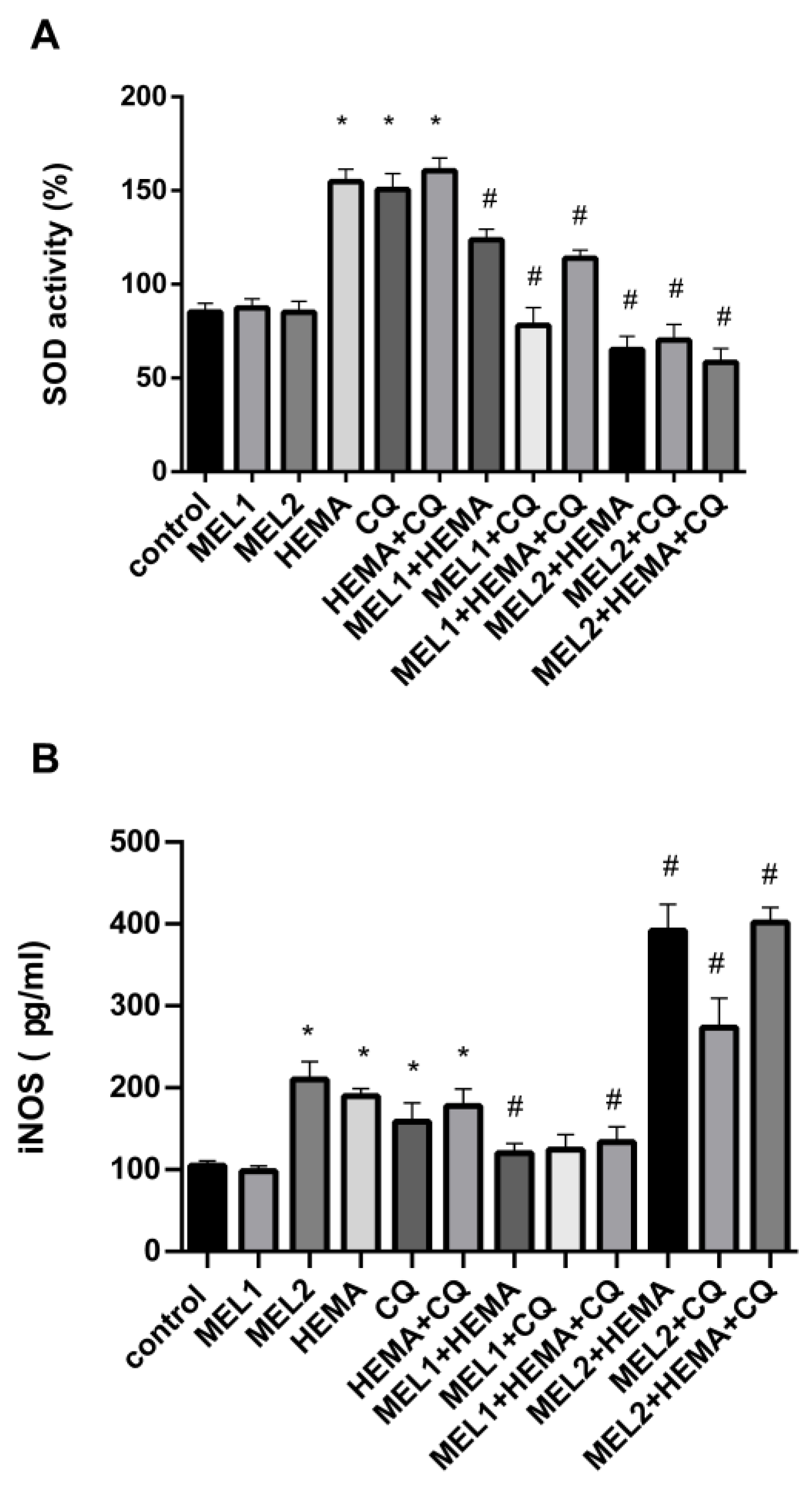
2.5. Melatonin Effects on SOD Activity and iNOS Protein Levels in hDPCs Treated with HEMA and/or CQ
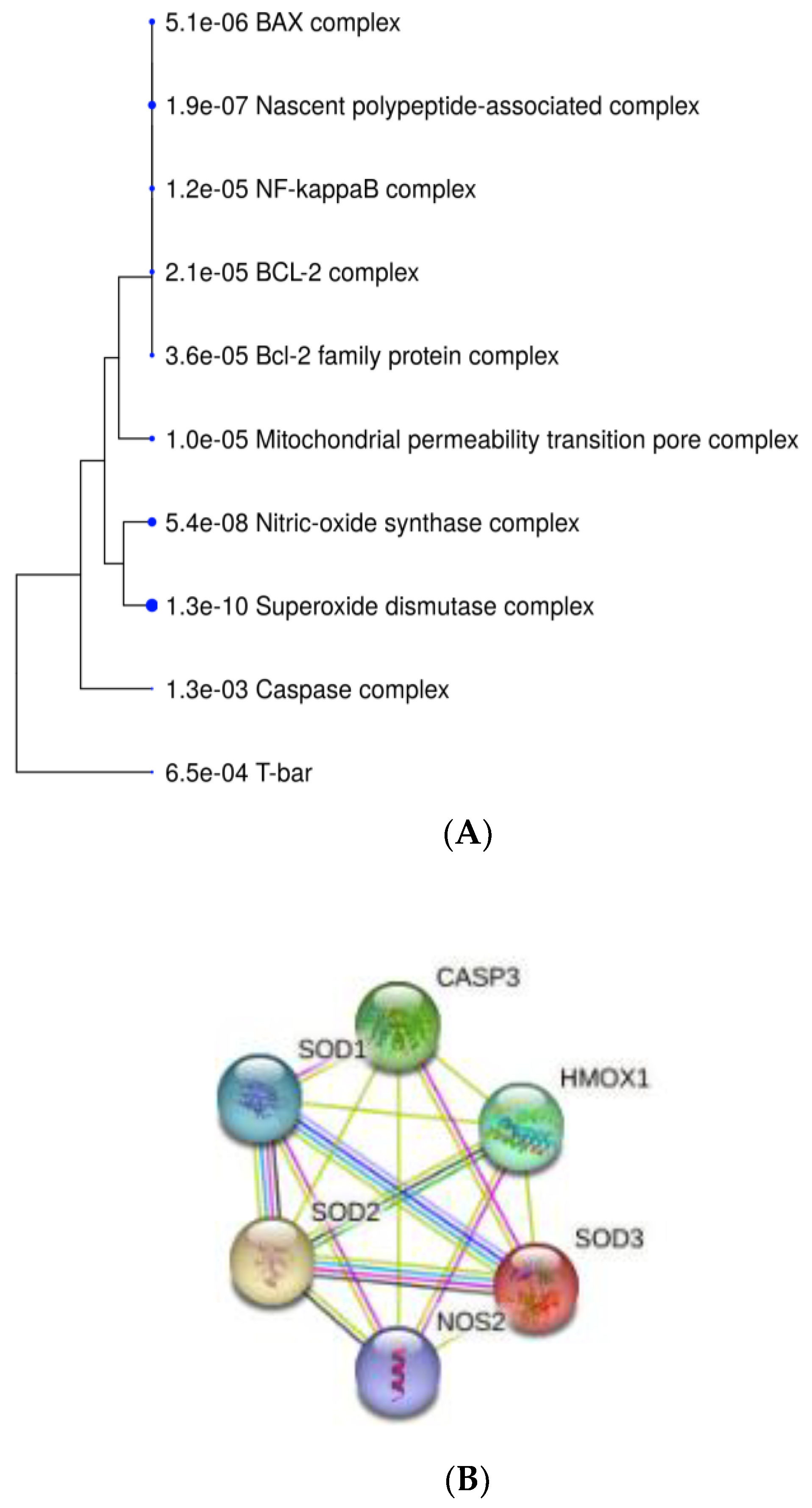
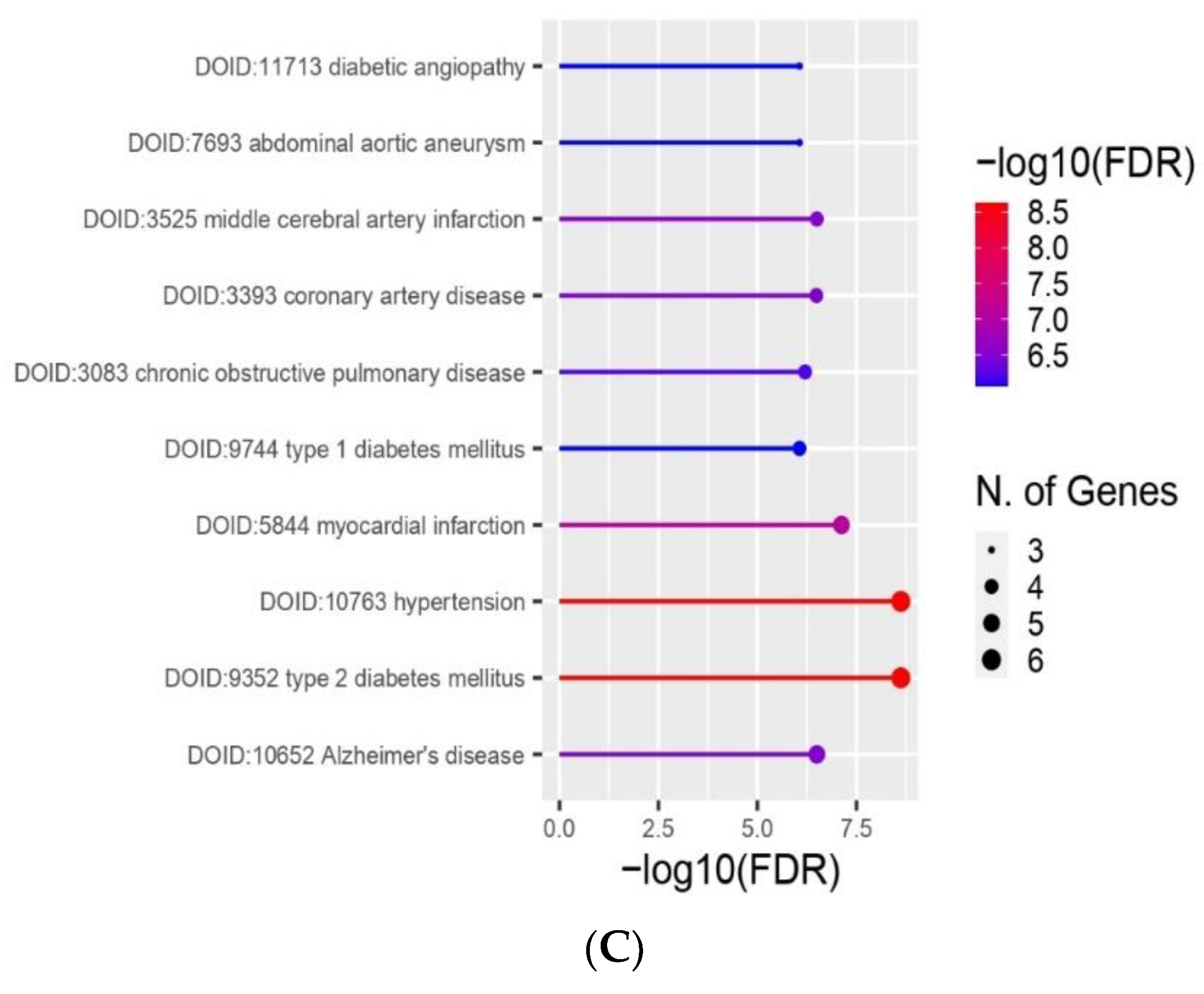
2.6. Enrichment Analyses and Gene Protein–Protein Interaction Network
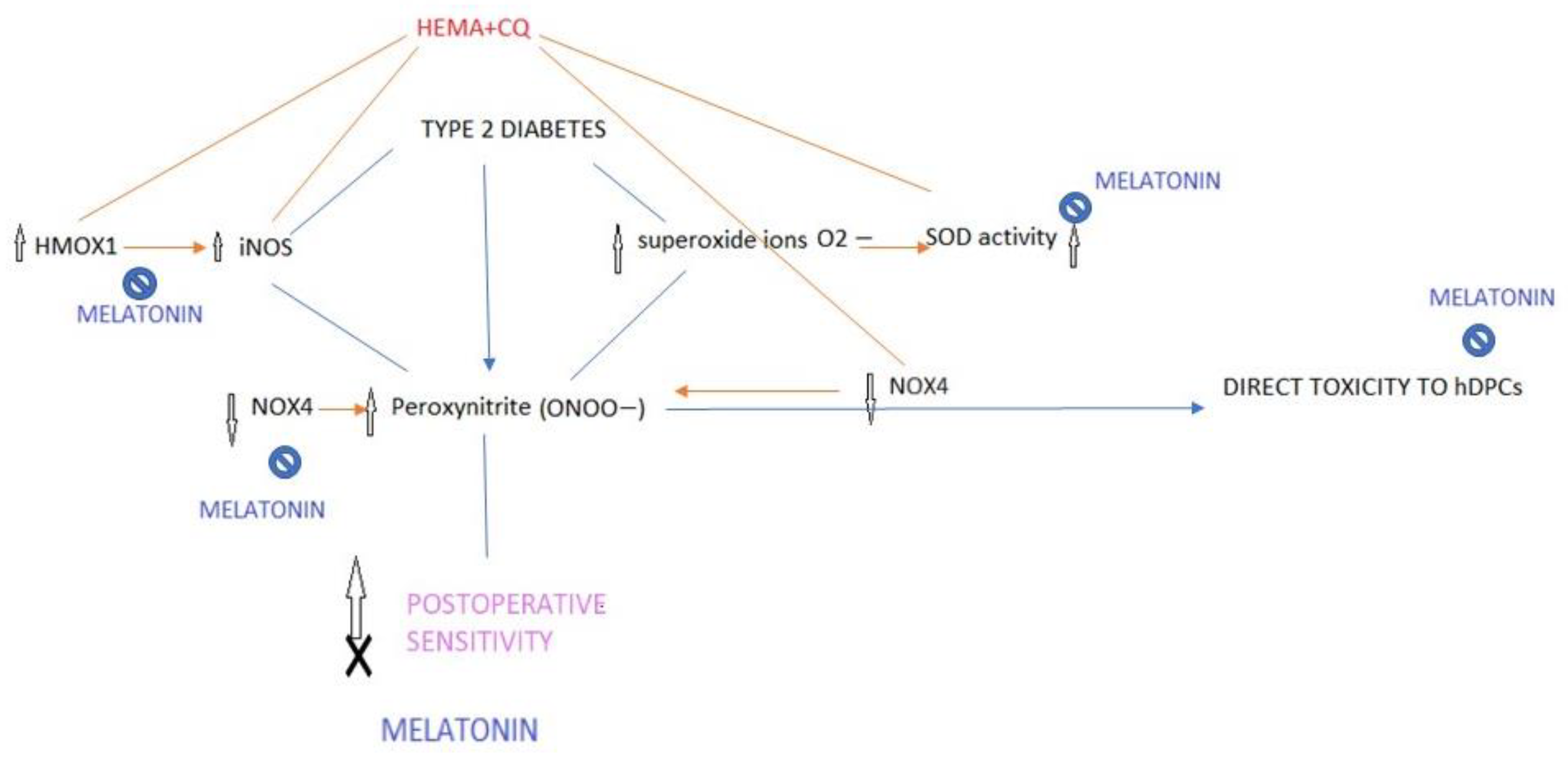
3. Discussion
4. Materials and Methods
4.1. Human Dental Pulp Tissue Collection
4.2. Cell Culture
4.3. Retrospective Chart Review for Detection of Postoperative Hypersensitivity
4.4. MTT Assay
4.5. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.6. Spectrophotometric Analysis
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Bioinformatic Analyses
4.9. Data Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Landuyt, K.L.; Nawrot, T.; Geebelen, B.; De Munck, J.; Snauwaert, J.; Yoshihara, K.; Scheers, H.; Godderis, L.; Hoet, P.; Van Meerbeek, B. How much do resin-based dental materials release? A meta-analytical approach. Dent. Mater. 2011, 27, 723–747. [Google Scholar] [CrossRef] [PubMed]
- Putzeys, E.; Duca, R.C.; Coppens, L.; Vanoirbeek, J.; Godderis, L.; Van Meerbeek, B.; Van Landuyt, K.L. In-vitro transdentin-al diffusion of monomers from adhesives. J. Dent. 2018, 75, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Bouillaguet, S. Biological Risks of Resin-based Materials to the Detin-Pulp Complex. Crit. Rev. Oral Biol. Med. 2004, 15, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Blasiak, J.; Synowiec, E.; Tarnawska, J.; Czarny, P.; Poplawski, T.; Reiter, R.J. Dental methacrylates may exert genotoxic ef-fects via the oxidative induction of DNA double strand breaks and the inhibition of their repair. Mol. Biol. Rep. 2012, 39, 7487–7496. [Google Scholar] [CrossRef]
- Sun, F.; Mao, P.; Wang, C.; Shi, C.; Nie, R.; Han, N.; Han, X. Cytotoxic Effects of One-step Self-etching Dental Adhesives on Human Periodontal Ligament Fibroblasts In Vitro. J. Adhes. Dent. 2016, 18, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Lin, L.D.; Wu, M.T.; Chan, C.P.; Chang, H.H.; Lee, M.S.; Sun, T.Y.; Jeng, P.Y.; Yeung, S.Y.; Lin, H.J.; et al. Ef-fects of Camphorquinone on Cytotoxicity, Cell Cycle Regulation and Prostaglandin E2 Production of Dental Pulp Cells: Role of ROS, ATM/Chk2, MEK/ERK and Hemeoxygenase-1. PLoS ONE 2015, 10, e0143663. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Tian, R.; Yang, Z.; Peng, Y.Y.; Lu, N. Quercetin Suppressed NADPH Oxidase-Derived Oxidative Stress via Heme Oxygenase-1 Induction in Macrophages. Arch. Biochem. Biophys. 2019, 671, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Huang, F.; Li, C.; Qu, H.; Gao, Z.; Leng, S.; Li, D.; He, H. Involvement of NOS/NO in the development of chronic dental inflammatory pain in rats. Brain Res. Rev. 2009, 59, 324–332. [Google Scholar] [CrossRef]
- Janes, K.; Neumann, W.L.; Salvemini, D. Anti-superoxide and anti-peroxynitrite strategies in pain suppression. Biochim. Biophys. Acta. 2012, 1822, 815–821. [Google Scholar] [CrossRef]
- Vareniuk, I.; Pavlov, I.A.; Obrosova, I.G. Inducible nitric oxide synthase gene deficiency counteracts multiple manifestations of peripheral neuropathy in a streptozotocin-induced mouse model of diabetes. Diabetologia 2008, 51, 2126–2133. [Google Scholar] [CrossRef]
- Rahim-Williams, B.; Tomar, S.; Blanchard, S.; Riley, J.L. 3rd. Influences of adult-onset diabetes on orofacial pain and related health behaviors. J. Public Health Dent. 2010, 70, 85–92. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Hardeland, R.; Lopez-Burillo, S.; Mayo, J.C.; Sainz, R.M.; Reiter, R.J. Melatonin: A hormone, a tissue factor, an autocoid, a paracoid, and an antioxidant vitamin. J. Pineal Res. 2003, 34, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Paterniti, I.; Mazzon, E.; Bramanti, P.; Cuzzocrea, S. Melatonin reduces hyperalgesia associated with inflammation. J. Pineal Res. 2010, 49, 321–331. [Google Scholar] [CrossRef] [PubMed]
- le Grand, S.M.; Patumraj, S.; Phansuwan-Pujito, P.; Srikiatkhachorn, A. Melatonin inhibits cortical spreading depression-evoked trigeminal nociception. Neuroreport 2006, 17, 1709–1713. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, A.; DJukić, L.; Toljić, B.; Milašin, J.; DŽeletović, B.; Brković, B.; Roganović, J. Melatonin levels in human dia-betic dental pulp tissue and its effects on dental pulp cells under hyperglycaemic conditions. Int. Endod. J. 2018, 51, 1149–1158. [Google Scholar] [CrossRef]
- Dennery, P.A.; Wong, H.E.; Sridhar, K.J.; Rodgers, P.A.; Sim, J.E.; Spitz, D.R. Differences in basal and hyperoxia-associated HO expression in oxidant-resistant hamster fibroblasts. Am. J. Physiol. Lung Cell Mol. Physiol. 1996, 271, L672–L679. [Google Scholar] [CrossRef]
- Suttner, D.M.; Dennery, P.A. Reversal of HO-1 related cytoprotection with increased expression is due to reactive iron. FASEB J. 1999, 13, 1800–1809. [Google Scholar] [CrossRef]
- Siitonen, T.; Siitonen, P.; Kinnula, V.; Mäntymaa, P.; Savolainen, E.R.; Koistinen, P. Contribution of the induction of heme oxygenase-1 to etoposide-induced apoptosis in acute myeloblastic leukemia. Haematologica 2003, 88, 112–113. [Google Scholar]
- Jia, L.; Wang, Y.; Wang, Y.; Ma, Y.; Shen, J.; Fu, Z.; Wu, Y.; Su, S.; Zhang, Y.; Cai, Z.; et al. Heme Oxygenase-1 in Macrophages Drives Septic Cardiac Dysfunction via Suppressing Lysosomal Degradation of Inducible Nitric Oxide Synthase. Circ. Res. 2018, 122, 1532–1544. [Google Scholar] [CrossRef]
- Basuroy, S.; Tcheranova, D.; Bhattacharya, S.; Leffler, C.W.; Parfenova, H. Nox4 NADPH oxidase-derived reactive oxygen species, via endogenous carbon monoxide, promote survival of brain endothelial cells during TNF-α-induced apoptosis. Am. J. Physiol. Cell Physiol. 2011, 300, C256–C265. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Deffert, C.; Donati, Y.; Basset, O.; Hinz, B.; Preynat-Seauve, O.; Guichard, C.; Arbiser, J.L.; Banfi, B.; Pache, J.C.; et al. A key role for NOX4 in epithelial cell death during development of lung fibrosis. Antioxid. Redox Signal. 2011, 15, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.X.; Hafstad, A.D.; Beretta, M.; Zhang, M.; Molenaar, C.; Kopec, J.; Fotinou, D.; Murray, T.V.; Cobb, A.M.; Martin, D.; et al. Targeted redox inhibition of protein phosphatase 1 by Nox4 regulates eIF2α-mediated stress signaling. EMBO J. 2016, 35, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.; Park, S.H.; Lin, L.M.; Chiappelli, F.; Huang, G.T. Substance P induces interleukin-8 secretion from human dental pulp cells. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2003, 96, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time uantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat Genet 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Binder, J.X.; Pletscher-Frankild, S.; Tsafou, K.; Stolte, C.; O’Donoghue, S.I.; Schneider, R.; Jensen, L.J. COMPARTMENTS: Unification and visualization of protein subcellular localization evidence. Database (Oxford) 2014, 2014, bau012. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2019, 36, 2628–2629. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ||
|---|---|---|
| Non-Diabetic | Type 2 Diabetic | |
| Number of patients Age (mean ± SEM) Number of fillings Fillings with postoperative sensitivity n (% of total) | 32 55.2 ± 3.3 44 5 (11.4%) | 32 58.6 ± 3.0 87 25 (28.7%) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilić, J.; Milosavljević, A.; Lazarević, M.; Milošević Marković, M.; Milašin, J.; Vučetić, M.; Chaurasia, A.; Miletić, V.; Roganović, J. Melatonin Mitigates iNOS-Related Effects of HEMA and Camphorquinone in Human Dental Pulp Cells: Relevance for Postoperative Sensitivity Mechanism in Type 2 Diabetes. Int. J. Mol. Sci. 2023, 24, 2562. https://doi.org/10.3390/ijms24032562
Ilić J, Milosavljević A, Lazarević M, Milošević Marković M, Milašin J, Vučetić M, Chaurasia A, Miletić V, Roganović J. Melatonin Mitigates iNOS-Related Effects of HEMA and Camphorquinone in Human Dental Pulp Cells: Relevance for Postoperative Sensitivity Mechanism in Type 2 Diabetes. International Journal of Molecular Sciences. 2023; 24(3):2562. https://doi.org/10.3390/ijms24032562
Chicago/Turabian StyleIlić, Jugoslav, Aleksandra Milosavljević, Miloš Lazarević, Maja Milošević Marković, Jelena Milašin, Milan Vučetić, Akhilanand Chaurasia, Vesna Miletić, and Jelena Roganović. 2023. "Melatonin Mitigates iNOS-Related Effects of HEMA and Camphorquinone in Human Dental Pulp Cells: Relevance for Postoperative Sensitivity Mechanism in Type 2 Diabetes" International Journal of Molecular Sciences 24, no. 3: 2562. https://doi.org/10.3390/ijms24032562
APA StyleIlić, J., Milosavljević, A., Lazarević, M., Milošević Marković, M., Milašin, J., Vučetić, M., Chaurasia, A., Miletić, V., & Roganović, J. (2023). Melatonin Mitigates iNOS-Related Effects of HEMA and Camphorquinone in Human Dental Pulp Cells: Relevance for Postoperative Sensitivity Mechanism in Type 2 Diabetes. International Journal of Molecular Sciences, 24(3), 2562. https://doi.org/10.3390/ijms24032562