
Interaction of High-Molecular Weight Fucoidan from Laminaria hyperborea with Natural Functions of the Retinal Pigment Epithelium




Abstract
1. Introduction
2. Results
2.1. Effects on Mid-Term Morphology and Viability
2.2. Effects on Wound Healing
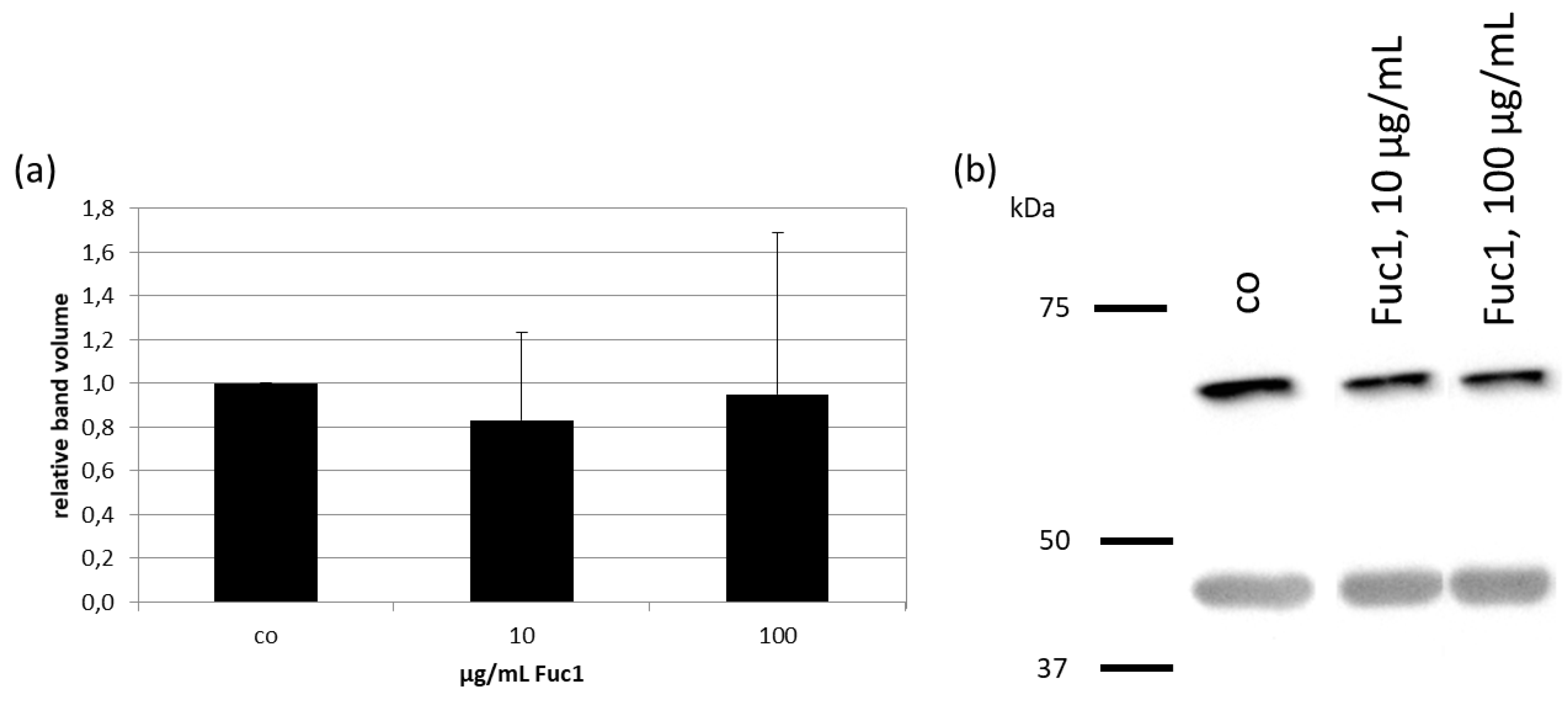
2.3. Effects on RPE65 Expression
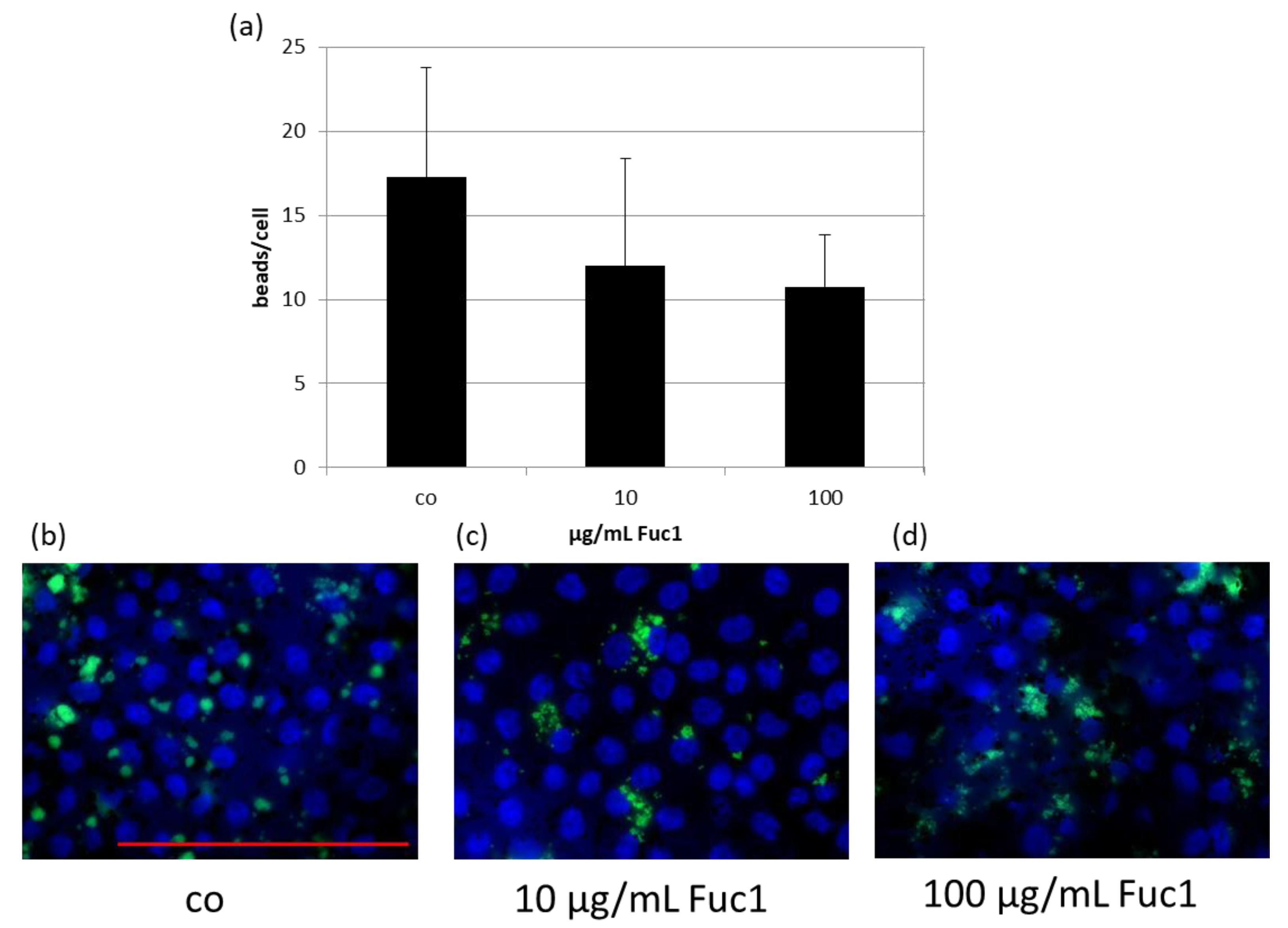
2.4. Effects on Phagocytosis
2.5. Effects on VEGF Secretion
2.6. Effects on Gene Expression
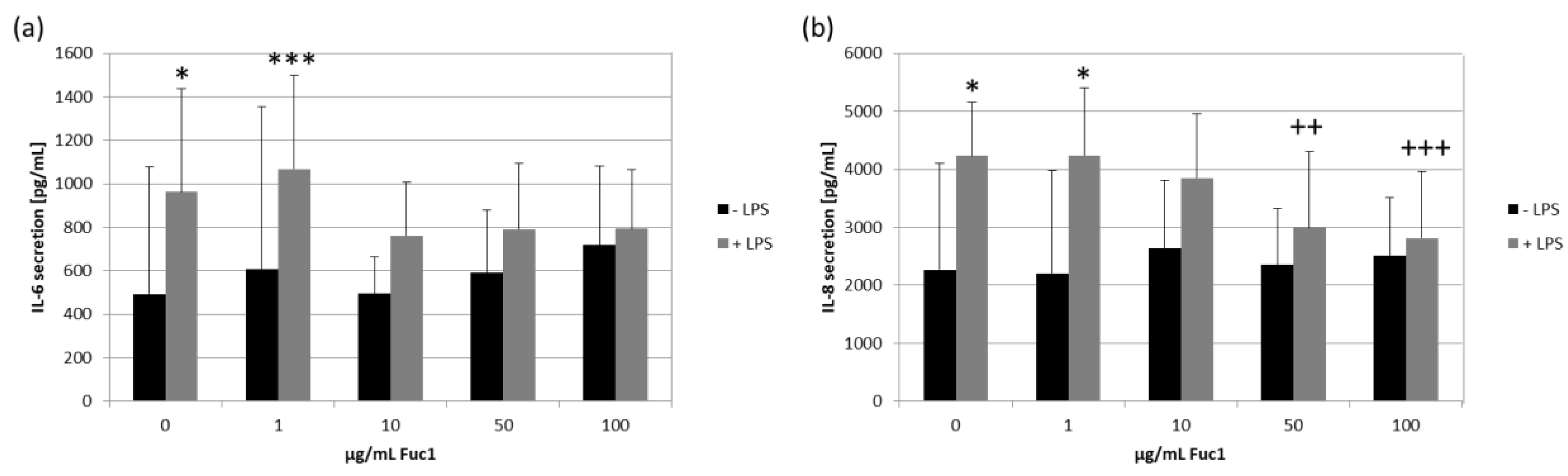
2.7. Secretion of Proinflammatory Cytokines
2.8. Characterization through Vibrational Spectroscopy—FTIR and Raman
3. Discussion
4. Material and Methods
4.1. Cell Culture and Fucoidan
4.2. Methyl Thiazolyl Tetrazolium Assay
4.3. Scratch Assay
4.4. Western Blot
4.5. Phagocytosis Assay
4.6. RPE/Choroid Organ Cultures
4.7. Real-Time PCR
4.8. Inflammation Assays
4.9. Vibrational Spectroscopy—FTIR and Raman
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Assay ID | RQ | p-Value |
|---|---|---|---|
| 18S-RNA | Hs99999901_s1 | 0.903 | 0.868 |
| ABCA1 | Ss04955209_m1 | 1.799 | 0.394 |
| ABCA4 | Ss06884373_m1 | 0.69 | 0.507 |
| ACE | Ss04953780_g1 | 1.276 | 0.78 |
| ACTB | Ss03376081_u1 | - | - |
| ALB | Ss03378640_u1 | 1.062 | 0.949 |
| ANXA5 | Ss06880508_m1 | 1.196 | 0.405 |
| APOE | Ss03394681_m1 | 0.855 | 0.381 |
| B2M | Ss03391154_m1 | 1.34 | 0.538 |
| BDNF | Ss03822335_s1 | 1.132 | 0.731 |
| C2 | Ss03389255_m1 | 1.076 | 0.636 |
| C3 | Ss03391255_m1 | 2.036 | 0.566 |
| C5 | Ss03391586_m1 | 1.069 | 0.832 |
| C9 | Ss03388866_m1 | 1.13 | 0.868 |
| CCL11 | Ss03377222_u1 | 0.615 | 0.089 |
| CCL19 | Ss04246386_m1 | 0.868 | 0.83 |
| CCL2 | Ss03394377_m1 | 1.182 | 0.839 |
| CD46 | Ss03392461_u1 | 1.056 | 0.449 |
| CD55 | Ss03392383_m1 | 0.935 | 0.93 |
| CD59 | Ss03394252_m1 | 1.063 | 0.49 |
| CFB | Ss03389385_g1 | 1.028 | 0.956 |
| CFH | Ss03391439_m1 | 0.9 | 0.795 |
| CFI | Ss06935384_m1 | 0.872 | 0.684 |
| CLU | Ss03391130_m1 | 1.013 | 0.928 |
| COL14A1 | Ss06865093_m1 | 1.268 | 0.801 |
| CP | Ss04330806_m1 | 0.842 | 0.697 |
| CRP | Ss03390889_m1 | 0.615 | 0.089 |
| CRYA1 | Ss06837084_m1 | 0.596 | 0.544 |
| CRYAB | Ss06921086_m1 | 1.197 | 0.701 |
| CST3 | Ss03388477_m1 | 0.867 | 0.576 |
| CTSD | Ss03379762_u1 | 1.068 | 0.735 |
| CX3CR1 | Ss06883230_m1 | 0.615 | 0.089 |
| CXCL12 | Ss03391855_m1 | 0.81 | 0.892 |
| DICER1 | Ss04248150_m1 | 1.129 | 0.352 |
| EFEMP1 | Ss04326723_m1 | 0.924 | 0.853 |
| ELN | Ss04955056_m1 | 0.401 | 0.241 |
| ERCC6 | Ss06934989_m1 | 1.345 | 0.408 |
| FANCG | Ss06877694_g1 | 1.075 | 0.766 |
| FASLG | Ss03381579_u1 | 0.564 | 0.54 |
| FBLN5 | Ss04805024_m1 | 0.977 | 0.947 |
| FLT1 | Ss03375679_u1 | 0.939 | 0.832 |
| FN1 | Ss03373673_m1 | 0.208 | 0.416 |
| GAPDH | Ss03375629_u1 | - | - |
| GFAP | Ss03373547_m1 | 0.378 | 0.445 |
| GUSB | Ss03387751_u1 | - | - |
| HIF1A | Ss03390447_m1 | 1.344 | 0.348 |
| HMOX1 | Ss03378516_u1 | 1.029 | 0.941 |
| HO2 | Ss03376410_u1 | 0.936 | 0.758 |
| HTRA1 | Ss06876775_m1 | 0.214 | 0.43 |
| ICAM-1 | Ss03392385_m1 | 1.135 | 0.83 |
| ICA | Ss04955454_m1 | 0.727 | 0.47 |
| IGF1 | Ss03394499_m1 | 0.847 | 0.84 |
| IL6 | Ss07308316_g1 | 0.367 | 0.477 |
| IL8 | Ss03392437_m1 | 2.768 | 0.438 |
| KDR | Ss03375683_u1 | 0.722 | 0.734 |
| LEP | Ss03392404_m1 | 0.615 | 0.089 |
| MAPK1 | Ss04248225_m1 | 1.275 | 0.579 |
| MAPK3 | Ss06887442_m1 | 13.062 | 0.389 |
| MMP2 | Ss03394318_m1 | 0.968 | 0.884 |
| MMP9 | Ss03392100_m1 | 0.47 | 0.503 |
| NFE2L2 | Ss06886076_m1 | 1.169 | 0.214 |
| NKAP | Ss04322419_m1 | 1.138 | 0.603 |
| NOS1 | Ss06838170_m1 | 0.685 | 0.159 |
| NOS2 | Ss03374608_u1 | 0.753 | 0.412 |
| NOS3 | Ss03383840_u1 | 0.885 | 0.627 |
| NOX4 | Ss06909549_m1 | 0.84 | 0.545 |
| PES1 | Ss04327334_m1 | 8.057 | 0.412 |
| PGK1 | Ss03389144_m1 | 1.328 | 0.118 |
| PLG | Ss03380545_u1 | 0.865 | 0.815 |
| PTGS2 | Ss03394694_m1 | 1.261 | 0.792 |
| RHO | Ss03394397_m1 | 0.893 | 0.962 |
| RLBP1 | Ss06905574_m1 | 0.676 | 0.568 |
| RPE65 | Ss06891920_m1 | 0.822 | 0.781 |
| SAG | Ss03382439_u1 | 0.772 | 0.881 |
| SCARB1 | Ss03391104_m1 | 1.035 | 0.757 |
| SERPINE1 | Ss03392656_u1 | 0.179 | 0.436 |
| SERPINF1 | Ss03385090_u1 | 0.687 | 0.481 |
| SERPING1 | Ss03387977_u1 | 0.862 | 0.698 |
| SOD2 | Ss03374828_m1 | 1.306 | 0.214 |
| SPARC | Ss03392006_m1 | 0.95 | 0.87 |
| TGFB1 | Ss04955543_m1 | 0.974 | 0.943 |
| TIMP1 | Ss03381944_u1 | 1.144 | 0.664 |
| TIMP-3 | Ss03375447_u1 | 0.851 | 0.719 |
| TLR3 | Ss03388862_m1 | 1.651 | 0.572 |
| TLR4 | Ss04956023_s1 | 0.632 | 0.653 |
| UBC | Ss03374343_g1 | 1.038 | 0.993 |
| VCAM1 | Ss03390912_m1 | 1.445 | 0.693 |
| VEGFA | Ss03393990_m1 | 1.226 | 0.418 |
| VIM | Ss04330801_gH | 0.245 | 0.496 |
| VLDLR | Ss03374049_m1 | 1.131 | 0.434 |
| VTN | Ss03382603_u1 | 0.772 | 0.506 |
| VWF | Ss04322692_m1 | 0.954 | 0.926 |
References
- Kloareg, B.; Demarty, M.; Mabeau, S. Polyanionic characteristics of purified sulphated homofucans from brown algae. Int. J. Biol. Macromol. 1986, 8, 380–386. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Stortz, C.A. A Comprehensive and Comparative Analysis of the Fucoidan Compositional Data Across the Phaeophyceae. Front. Plant Sci. 2020, 11, 556312. [Google Scholar] [CrossRef] [PubMed]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan Structure and Its Impact on Glucose Metabolism: Implications for Diabetes and Cancer Therapy. Mar. Drugs 2021, 19, 30. [Google Scholar] [CrossRef] [PubMed]
- Kopplin, G.; Rokstad, A.M.; Mélida, H.; Bulone, V.; Skjåk-Bræk, G.; Aachmann, F.L. Structural Characterization of Fucoidan from Laminaria hyperborea: Assessment of Coagulation and Inflammatory Properties and Their Structure–Function Relationship. ACS Appl. Bio Mater. 2018, 1, 1880–1892. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.-Y.; Kim, G.-Y.; Choi, I.-W.; Kim, N.D.; Nam, T.-J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef]
- Dörschmann, P.; Klettner, A. Fucoidans as Potential Therapeutics for Age-Related Macular Degeneration-Current Evidence from In Vitro Research. Int. J. Mol. Sci. 2020, 21, 9272. [Google Scholar] [CrossRef]
- Klettner, A. Fucoidan as a Potential Therapeutic for Major Blinding Diseases--A Hypothesis. Mar. Drugs 2016, 14, 31. [Google Scholar] [CrossRef]
- Dörschmann, P.; Schmitt, C.; Bittkau, K.S.; Neupane, S.; Synowitz, M.; Roider, J.; Alban, S.; Held-Feindt, J.; Klettner, A. Evaluation of a Brown Seaweed Extract from Dictyosiphon foeniculaceus as a Potential Therapeutic Agent for the Treatment of Glioblastoma and Uveal Melanoma. Mar. Drugs 2020, 18, 625. [Google Scholar] [CrossRef]
- Dörschmann, P.; Bittkau, K.S.; Neupane, S.; Roider, J.; Alban, S.; Klettner, A. Effects of Fucoidans from Five Different Brown Algae on Oxidative Stress and VEGF Interference in Ocular Cells. Mar. Drugs 2019, 17, 258. [Google Scholar] [CrossRef] [PubMed]
- Bittkau, K.S.; Dörschmann, P.; Blümel, M.; Tasdemir, D.; Roider, J.; Klettner, A.; Alban, S. Comparison of the Effects of Fucoidans on the Cell Viability of Tumor and Non-Tumor Cell Lines. Mar. Drugs 2019, 17, 441. [Google Scholar] [CrossRef] [PubMed]
- Dörschmann, P.; Kopplin, G.; Roider, J.; Klettner, A. Effects of Sulfated Fucans from Laminaria hyperborea Regarding VEGF Secretion, Cell Viability, and Oxidative Stress and Correlation with Molecular Weight. Mar. Drugs 2019, 17, 548. [Google Scholar] [CrossRef] [PubMed]
- Dörschmann, P.; Mikkelsen, M.D.; Thi, T.N.; Roider, J.; Meyer, A.S.; Klettner, A. Effects of a Newly Developed Enzyme-Assisted Extraction Method on the Biological Activities of Fucoidans in Ocular Cells. Mar. Drugs 2020, 18, 282. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, K.; Neupane, S.; Bittkau, K.S.; Galarza Pérez, M.; Dörschmann, P.; Roider, J.; Alban, S.; Klettner, A. Effects of Crude Fucus distichus Subspecies evanescens Fucoidan Extract on Retinal Pigment Epithelium Cells-Implications for Use in Age-Related Macular Degeneration. Mar. Drugs 2019, 17, 538. [Google Scholar] [CrossRef] [PubMed]
- Bittkau, K.S.; Neupane, S.; Alban, S. Initial evaluation of six different brown algae species as source for crude bioactive fucoidans. Algal Research 2020, 45, 101759. [Google Scholar] [CrossRef]
- Miller, J.W. Age-related macular degeneration revisited--piecing the puzzle: The LXIX Edward Jackson memorial lecture. Am. J. Ophthalmol. 2013, 155, 1–35.e13. [Google Scholar] [CrossRef]
- Klettner, A. Altersabhängige Makuladegeneration (AMD). Biologie und Behandlung. In Medizinische Monatsschrift für Pharmazeuten; Deutscher Apotheker Verlag Stuttgart 2015; Volume 38, pp. 258–264, quiz 265-6.
- Schmidt-Erfurth, U.; Chong, V.; Loewenstein, A.; Larsen, M.; Souied, E.; Schlingemann, R.; Eldem, B.; Monés, J.; Richard, G.; Bandello, F. Guidelines for the management of neovascular age-related macular degeneration by the European Society of Retina Specialists (EURETINA). Br. J. Ophthalmol. 2014, 98, 1144–1167. [Google Scholar] [CrossRef]
- Dithmer, M.; Fuchs, S.; Shi, Y.; Schmidt, H.; Richert, E.; Roider, J.; Klettner, A. Fucoidan reduces secretion and expression of vascular endothelial growth factor in the retinal pigment epithelium and reduces angiogenesis in vitro. PLoS One 2014, 9, e89150. [Google Scholar] [CrossRef]
- Dörschmann, P.; Apitz, S.; Hellige, I.; Neupane, S.; Alban, S.; Kopplin, G.; Ptak, S.; Fretté, X.; Roider, J.; Zille, M.; et al. Evaluation of the Effects of Fucoidans from Fucus Species and Laminaria hyperborea against Oxidative Stress and Iron-Dependent Cell Death. Mar. Drugs 2021, 19, 557. [Google Scholar] [CrossRef]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Aspects Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [PubMed]
- Strunnikova, N.V.; Maminishkis, A.; Barb, J.J.; Wang, F.; Zhi, C.; Sergeev, Y.; Chen, W.; Edwards, A.O.; Stambolian, D.; Abecasis, G.; et al. Transcriptome analysis and molecular signature of human retinal pigment epithelium. Hum. Mol. Genet. 2010, 19, 2468–2486. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Roider, J. Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 8387. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, L.; Lucius, R.; Roider, J.; Klettner, A. Interaction of inflammatorily activated retinal pigment epithelium with retinal microglia and neuronal cells. Exp. Eye Res. 2020, 199, 108167. [Google Scholar] [CrossRef]
- Ptak, S.H.; Sanchez, L.; Fretté, X.; Kurouski, D. Complementarity of Raman and Infrared spectroscopy for rapid characterization of fucoidan extracts. Plant Methods 2021, 17, 130. [Google Scholar] [CrossRef]
- Synytsya, A.; Kim, W.-J.; Kim, S.-M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Il Park, Y. Structure and antitumour activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Pereira, L.; Amado, A.M.; Critchley, A.T.; van de Velde, F.; Ribeiro-Claro, P.J.A. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FT-Raman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef]
- Pielesz, A.; Biniaś, W.; Paluch, J. Mild acid hydrolysis of fucoidan: Characterization by electrophoresis and FT-Raman spectroscopy. Carbohydr. Res. 2011, 346, 1937–1944. [Google Scholar] [CrossRef]
- Pielesz, A.; Biniaś, W. Cellulose acetate membrane electrophoresis and FTIR spectroscopy as methods of identifying a fucoidan in Fucusvesiculosus Linnaeus. Carbohydr. Res. 2010, 345, 2676–2682. [Google Scholar] [CrossRef]
- Sekkal, M.; Legrand, P.; Huvenne, J.P.; Verdus, M.C. The use of FTIR microspectrometry as a new tool for the identification in situ of polygalactanes in red seaweeds. J. Mol. Struct. 1993, 294, 227–230. [Google Scholar] [CrossRef]
- Kiefer, W. Recent Advances in linear and nonlinear Raman spectroscopy I. J. Raman Spectrosc. 2007, 38, 1538–1553. [Google Scholar] [CrossRef]
- Mathlouthi, M.; Koenig, J.L. Vibrational spectra of carbohydrates. Adv. Carbohydr. Chem. Biochem. 1986, 44, 7–89. [Google Scholar] [CrossRef] [PubMed]
- Wiercigroch, E.; Szafraniec, E.; Czamara, K.; Pacia, M.Z.; Majzner, K.; Kochan, K.; Kaczor, A.; Baranska, M.; Malek, K. Raman and infrared spectroscopy of carbohydrates: A review. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2017, 185, 317–335. [Google Scholar] [CrossRef]
- Inoue, Y.; Nagasawa, K. Selective N-desulfation of heparin with dimethyl sulfoxide containing water or methanol. Carbohydr. Res. 1976, 46, 87–95. [Google Scholar] [CrossRef]
- Buzgar, N. The Raman Study of Certain Sulfates, 2nd ed.; Physics: College Park, MD, USA, 2009. [Google Scholar]
- Ben Mabrouk, K.; Kauffmann, T.H.; Aroui, H.; Fontana, M.D. Raman study of cation effect on sulfate vibration modes in solid state and in aqueous solutions. J. Raman Spectrosc. 2013, 44, 1603–1608. [Google Scholar] [CrossRef]
- Adebajo, M.O.; Frost, R.L.; Kloprogge, J.T.; Kokot, S. Raman spectroscopic investigation of acetylation of raw cotton. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2006, 64, 448–453. [Google Scholar] [CrossRef]
- Smith, E.; Dent, G. Modern Raman spectroscopy: A practical approach, 2nd ed.; Wiley: Hoboken, NJ, USA, 2019; ISBN 978-1-119-44055-0. [Google Scholar]
- Espina, A.; Sanchez-Cortes, S.; Jurašeková, Z. Vibrational Study (Raman, SERS, and IR) of Plant Gallnut Polyphenols Related to the Fabrication of Iron Gall Inks. Molecules 2022, 27, 279. [Google Scholar] [CrossRef]
- Chen, M.; Rajapakse, D.; Fraczek, M.; Luo, C.; Forrester, J.V.; Xu, H. Retinal pigment epithelial cell multinucleation in the aging eye—A mechanism to repair damage and maintain homoeostasis. Aging Cell 2016, 15, 436–445. [Google Scholar] [CrossRef]
- Cai, X.; Conley, S.M.; Naash, M.I. RPE65: Role in the visual cycle, human retinal disease, and gene therapy. Ophthalmic Genet. 2009, 30, 57–62. [Google Scholar] [CrossRef]
- Hamel, C.P.; Tsilou, E.; Harris, E.; Pfeffer, B.A.; Hooks, J.J.; Detrick, B.; Redmond, T.M. A developmentally regulated microsomal protein specific for the pigment epithelium of the vertebrate retina. J. Neurosci. Res. 1993, 34, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, D.; Feng, W.; Duncan, J.L.; Yasumura, D.; D’Cruz, P.M.; Chappelow, A.; Matthes, M.T.; Kay, M.A.; LaVail, M.M. Correction of the retinal dystrophy phenotype of the RCS rat by viral gene transfer of Mertk. Proc. Natl. Acad. Sci. USA 2001, 98, 12584–12589. [Google Scholar] [CrossRef]
- Pan, T.-J.; Li, L.-X.; Zhang, J.-W.; Yang, Z.-S.; Shi, D.-M.; Yang, Y.-K.; Wu, W.-Z. Antimetastatic Effect of Fucoidan-Sargassum against Liver Cancer Cell Invadopodia Formation via Targeting Integrin αVβ3 and Mediating αVβ3/Src/E2F1 Signaling. J. Cancer 2019, 10, 4777–4792. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Westhues, D.; Lassen, J.; Bartsch, S.; Roider, J. Regulation of constitutive vascular endothelial growth factor secretion in retinal pigment epithelium/choroid organ cultures: P38, nuclear factor κB, and the vascular endothelial growth factor receptor-2/phosphatidylinositol 3 kinase pathway. Mol. Vis. 2013, 19, 281–291. [Google Scholar] [PubMed]
- Ustyuzhanina, N.E.; Bilan, M.I.; Ushakova, N.A.; Usov, A.I.; Kiselevskiy, M.V.; Nifantiev, N.E. Fucoidans: Pro- or antiangiogenic agents? Glycobiology 2014, 24, 1265–1274. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- Rofagha, S.; Bhisitkul, R.B.; Boyer, D.S.; Sadda, S.R.; Zhang, K. Seven-year outcomes in ranibizumab-treated patients in ANCHOR, MARINA, and HORIZON: A multicenter cohort study (SEVEN-UP). Ophthalmology 2013, 120, 2292–2299. [Google Scholar] [CrossRef]
- Musiał-Kopiejka, M.; Polanowska, K.; Dobrowolski, D.; Krysik, K.; Wylęgała, E.; Grabarek, B.O.; Lyssek-Boroń, A. The Effectiveness of Brolucizumab and Aflibercept in Patients with Neovascular Age-Related Macular Degeneration. Int. J. Environ. Res. Public Health 2022, 19, 2303. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Faustova, N.M.; Obluchinskaya, E.D.; Kosman, V.M.; Vuorela, H.; Makarov, V.G. Pharmacokinetic and Tissue Distribution of Fucoidan from Fucus vesiculosus after Oral Administration to Rats. Mar. Drugs 2018, 16, 132. [Google Scholar] [CrossRef]
- Gabbia, D.; Dall’Acqua, S.; Di Gangi, I.M.; Bogialli, S.; Caputi, V.; Albertoni, L.; Marsilio, I.; Paccagnella, N.; Carrara, M.; Giron, M.C.; et al. The Phytocomplex from Fucus vesiculosus and Ascophyllum nodosum Controls Postprandial Plasma Glucose Levels: An In Vitro and In Vivo Study in a Mouse Model of NASH. Mar. Drugs 2017, 15, 41. [Google Scholar] [CrossRef]
- Shan, X.; Wang, X.; Jiang, H.; Cai, C.; Hao, J.; Yu, G. Fucoidan from Ascophyllumnodosum Suppresses Postprandial Hyperglycemia by Inhibiting Na+/Glucose Cotransporter 1 Activity. Mar. Drugs 2020, 18, 485. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-Y.; Lin, T.-Y.; Lu, M.-K.; Leng, P.-J.; Tsao, S.-M.; Wu, Y.-C. Fucoidan induces Toll-like receptor 4-regulated reactive oxygen species and promotes endoplasmic reticulum stress-mediated apoptosis in lung cancer. Sci. Rep. 2017, 7, 44990. [Google Scholar] [CrossRef]
- Dutot, M.; Grassin-Delyle, S.; Salvator, H.; Brollo, M.; Rat, P.; Fagon, R.; Naline, E.; Devillier, P. A marine-sourced fucoidan solution inhibits Toll-like-receptor-3-induced cytokine release by human bronchial epithelial cells. Int. J. Biol. Macromol. 2019, 130, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Yan, C.; Ai, C.; Wen, C.; Guo, X.; Song, S. Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately. Foods 2022, 11, 2381. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Roider, J. Comparison of bevacizumab, ranibizumab, and pegaptanib in vitro: Efficiency and possible additional pathways. Invest. Ophthalmol. Vis. Sci. 2008, 49, 4523–4527. [Google Scholar] [CrossRef] [PubMed]
- Wiencke, A.K.; Kiilgaard, J.F.; Nicolini, J.; Bundgaard, M.; Röpke, C.; La Cour, M. Growth of cultured porcine retinal pigment epithelial cells. Acta Ophthalmol. Scand. 2003, 81, 170–176. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.; Worzella, T.J.; Minor, L. Cell Viability Assays; Markossian, S., Grossman, A., Brimacombe, K., Arkin, M., Auld, D., Austin, C., Baell, J., Chung, T.D.Y., Coussens, N.P., Dahlin, J.L., et al., Eds.; Assay Guidance Manual; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. Available online: https://www.ncbi.nlm.nih.gov/books/NBK144065/ (accessed on 1 December 2022).
- Klettner, A.; Brinkmann, A.; Winkelmann, K.; Käckenmeister, T.; Hildebrandt, J.; Roider, J. Effect of long-term inflammation on viability and function of RPE cells. Exp. Eye Res. 2020, 200, 108214. [Google Scholar] [CrossRef]
- Dithmer, M.; Kirsch, A.-M.; Richert, E.; Fuchs, S.; Wang, F.; Schmidt, H.; Coupland, S.E.; Roider, J.; Klettner, A. Fucoidan Does Not Exert Anti-Tumorigenic Effects on Uveal Melanoma Cell Lines. Mar. Drugs 2017, 15, 193. [Google Scholar] [CrossRef]
- Klettner, A.; Möhle, F.; Lucius, R.; Roider, J. Quantifying FITC-labeled latex beads opsonized with photoreceptor outer segment fragments: An easy and inexpensive method of investigating phagocytosis in retinal pigment epithelium cells. Ophthalmic Res. 2011, 46, 88–91. [Google Scholar] [CrossRef]
- Miura, Y.; Klettner, A.; Noelle, B.; Hasselbach, H.; Roider, J. Change of morphological and functional characteristics of retinal pigment epithelium cells during cultivation of retinal pigment epithelium-choroid perfusion tissue culture. Ophthalmic Res. 2010, 43, 122–133. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








| Wavenumber [cm−1] | Assignment | Activity |
|---|---|---|
| 576 | δ(pyranoid ring C-C-O) | Infrared |
| 586 | δs(pyranoid ring C-C-O) | Raman |
| 680 | δ(pyranoid ring C-O-C) | Infrared |
| 824 (shoulder) | δ(2C-O-S) | Raman |
| 838 | ν(α-pyranoid ring C-C) + ν(2C-O-S) | Infrared |
| 839 | δ(4C-O-S) | Raman |
| 911 | τ(CH2) + δ(2C-O-S) | Infrared |
| 961 | ν(C-O in C-O-C) + δ(C-O-H) | Infrared |
| 994 | νs(C-O-S) + ν(glycosidic C-O) | Raman |
| 1030 | δ(C-O-H) + ν(C-C) + ν(C-O-S) | Infrared |
| 1066 | νs(O=S=O) + ν(2C-O-S) | Raman |
| 1078 | ν(C-O) + ν(C-C) + δ(C-O-H) | Infrared |
| 1128 | ν(C-O-C) + ν(C-O-S) | Infrared |
| 1162 | ν(C-O) | Infrared |
| ~970–1200 | ν(pyranoid ring C-C) + ν(C-O) | Raman |
| 1217 | ν(S=O) | Infrared |
| 1257 (shoulder) | ν(S=O) | Infrared |
| 1273 | νas(O=S=O) | Raman |
| 1336 | ν(C-O) + δ(C-O-H) | Raman |
| 1387 | δs(CH3) | Infrared |
| 1454 | δs(CH3) | Raman |
| 1636 | ν(C=O) + ν(C=C) | Infrared |
| ~1650–1800 | ν(C=O) | Raman |
| 2947 | ν(C-H) | Raman |
| 3409 | ν(O-H) | Infrared |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dörschmann, P.; Kopplin, G.; Roider, J.; Klettner, A. Interaction of High-Molecular Weight Fucoidan from Laminaria hyperborea with Natural Functions of the Retinal Pigment Epithelium. Int. J. Mol. Sci. 2023, 24, 2232. https://doi.org/10.3390/ijms24032232
Dörschmann P, Kopplin G, Roider J, Klettner A. Interaction of High-Molecular Weight Fucoidan from Laminaria hyperborea with Natural Functions of the Retinal Pigment Epithelium. International Journal of Molecular Sciences. 2023; 24(3):2232. https://doi.org/10.3390/ijms24032232
Chicago/Turabian StyleDörschmann, Philipp, Georg Kopplin, Johann Roider, and Alexa Klettner. 2023. "Interaction of High-Molecular Weight Fucoidan from Laminaria hyperborea with Natural Functions of the Retinal Pigment Epithelium" International Journal of Molecular Sciences 24, no. 3: 2232. https://doi.org/10.3390/ijms24032232
APA StyleDörschmann, P., Kopplin, G., Roider, J., & Klettner, A. (2023). Interaction of High-Molecular Weight Fucoidan from Laminaria hyperborea with Natural Functions of the Retinal Pigment Epithelium. International Journal of Molecular Sciences, 24(3), 2232. https://doi.org/10.3390/ijms24032232