
Cap ‘n’ Collar C and Aryl Hydrocarbon Receptor Nuclear Translocator Facilitate the Expression of Glutathione S-Transferases Conferring Adaptation to Tannic Acid and Quercetin in Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae)

 and
and
Abstract
1. Introduction
2. Results
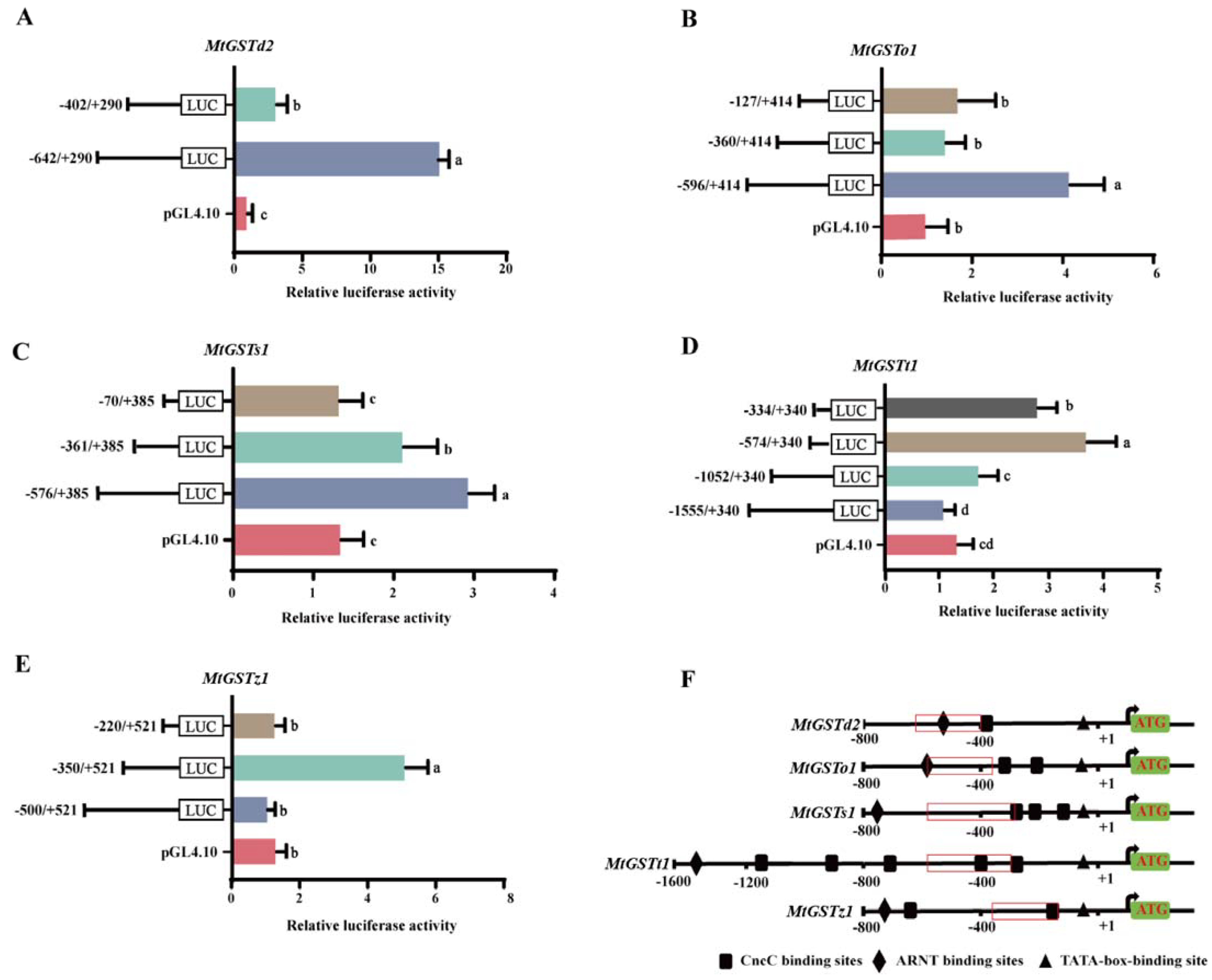
2.1. MtGST Promoter Activity Analysis
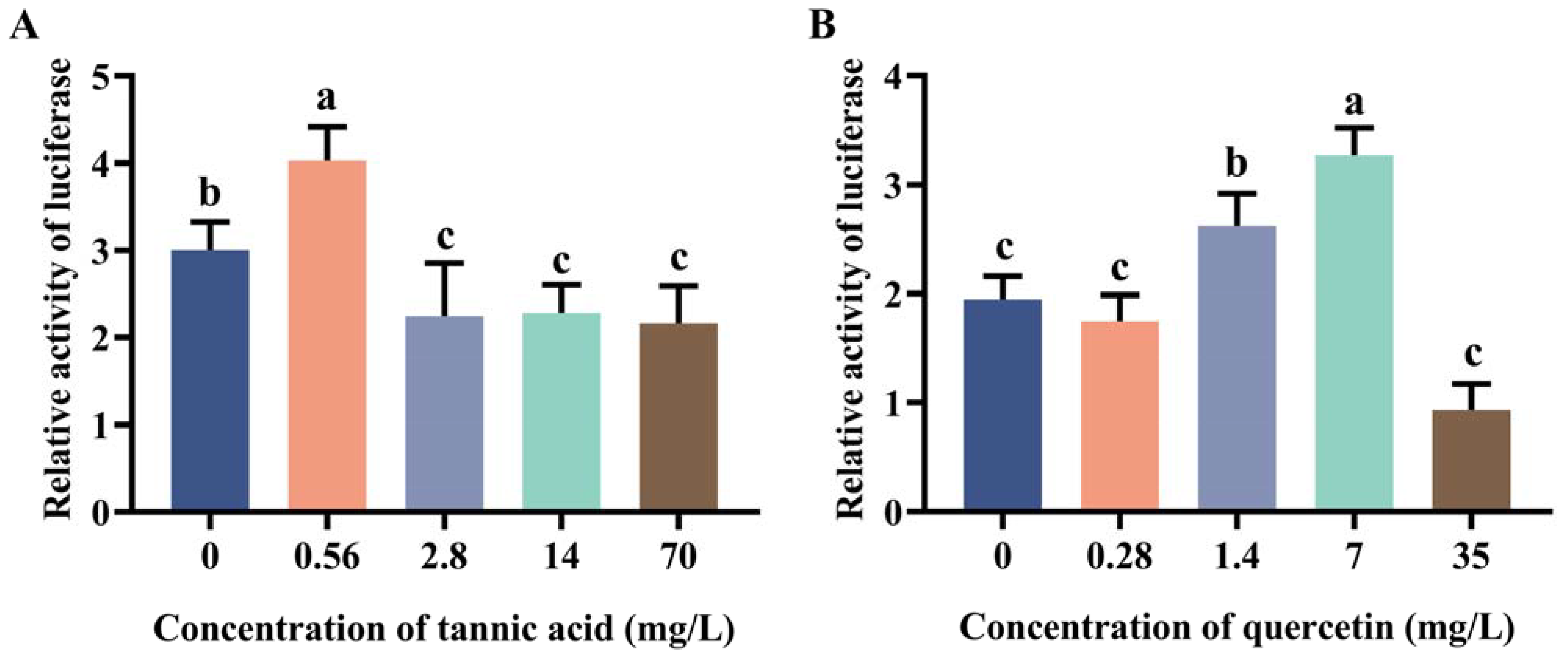
2.2. Induction Effect of Tannic Acid and Quercetin on the Core Region of Two Promoters
2.3. Cloning and Phylogenetic Analysis of MtCncC
2.4. Cloning and Phylogenetic Analysis of MtARNT
2.5. Analysis of the Transcriptional Activity of MtGST Promoters Regulated by MtCncC/MtARNT
3. Discussion
4. Materials and Methods
4.1. Insect Rearing and Cell Culture
4.2. Cloning and Sequencing 5’ Loss Fragments of GST Promoters
4.3. Promoter Activity Analysis by Luciferase Reporter Assays
4.4. Cloning the Sequences of MtCncC and MtARNT Genes
4.5. Phylogenetic Analysis of MtCncC and MtARNT
4.6. Cotransfection of MtCncC and MtARNT with MtGSTt1 (−574/+340) or MtGSTz1 (−350/+521) Promoter
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krieger, R.I.; Feeny, P.P.; Wilkinson, C.F. Detoxication enzymes in the guts of caterpillars: An evolutionary answer to plant defenses? Science 1971, 172, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Schuler, M.A. The role of cytochrome P450 monooxygenases in plant-insect interactions. Plant Physiol. 1996, 112, 1411–1419. [Google Scholar] [CrossRef]
- Tang, F.; Tu, H.; Shang, Q.; Gao, X.; Liang, P. Molecular Cloning and Characterization of Five Glutathione S-Transferase Genes and Promoters from Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae) and Their Response to Tannic Acid Stress. Insects 2020, 11, 339. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, F.; Xu, M.; Shen, T. Exploring miRNA-mRNA regulatory modules responding to tannic acid stress in Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae) via small RNA sequencing. Bull. Entomol. Res. 2022, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Zhang, X.; Liu, Y.; Gao, X.; Liu, N. In vitro inhibition of glutathione S-transferases by several insecticides and allelochemicals in two moth species. Int. J. Pest Manag. 2014, 60, 33–38. [Google Scholar] [CrossRef]
- Cheng, H.; Tang, F.; Li, W.; Xu, M. Tannic acid induction of a glutathione S-transferase in Micromelalopha troglodyta (Lepidoptera: Notodontidae) larvae. J. Entomol. Sci. 2015, 50, 350–362. [Google Scholar] [CrossRef]
- Despres, L.; David, J.P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef]
- Koirala, B.K.S.; Moural, T.; Zhu, F. Functional and Structural Diversity of Insect Glutathione S-transferases in Xenobiotic Adaptation. Int. J. Biol. Sci. 2022, 18, 5713–5723. [Google Scholar] [CrossRef]
- Lee, K. Glutathione S-transferase activities in phytophagous insects: Induction and inhibition by plant phototoxins and phenols. Insect Biochem. 1991, 21, 353–361. [Google Scholar] [CrossRef]
- Gao, X.W.; Dong, X.L.; Zheng, B.Z.; Chen, Q. Glutathiones S-transferase (GSTs) of cotton bollworm: Induction of pesticides and plant secondary substances and metabolism of GSTs to pesticides. Acta Entomol. Sin. 1997, 40, 122–127. [Google Scholar]
- Francis, F.; Vanhaelen, N.; Haubruge, E. Glutathione S-transferases in the adaptation to plant secondary metabolites in the Myzus persicae aphid. Arch. Insect Biochem. Physiol. 2005, 58, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Sun, L.; Zhao, H.; Wang, Z.; Zou, L.; Cao, C. Functional identification and characterization of GST genes in the Asian gypsy moth in response to poplar secondary metabolites. Pestic. Biochem. Physiol. 2021, 176, 104860. [Google Scholar] [CrossRef]
- AlJabr, A.M.; Hussain, A.; Rizwan-Ul-Haq, M.; Al-Ayedh, H. Toxicity of Plant Secondary Metabolites Modulating Detoxification Genes Expression for Natural Red Palm Weevil Pesticide Development. Molecules 2017, 22, 169. [Google Scholar] [CrossRef]
- Palli, S.R. CncC/Maf-mediated xenobiotic response pathway in insects. Arch. Insect Biochem. Physiol. 2020, 104, e21674. [Google Scholar] [CrossRef]
- Li, X.; Deng, Z.; Chen, X. Regulation of insect P450s in response to phytochemicals. Curr. Opin. Insect Sci. 2021, 43, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Song, Y.; Zeng, R. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr. Opin. Insect Sci. 2021, 43, 103–107. [Google Scholar] [CrossRef]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Misra, J.R.; Horner, M.A.; Lam, G.; Thummel, C.S. Transcriptional regulation of xenobiotic detoxification in Drosophila. Genes. Dev. 2011, 25, 1796–1806. [Google Scholar] [CrossRef]
- Deng, H.; Kerppola, T.K. Regulation of Drosophila metamorphosis by xenobiotic response regulators. PLoS Genet. 2013, 9, e1003263. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lu, M.; Zhang, N.; Zou, X.; Mo, M.; Zheng, S. Nuclear factor erythroid-derived 2-related factor 2 activates glutathione S-transferase expression in the midgut of Spodoptera litura (Lepidoptera: Noctuidae) in response to phytochemicals and insecticides. Insect Mol. Biol. 2018, 27, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Kalsi, M.; Palli, S.R. Transcription factor cap n collar C regulates multiple cytochrome P450 genes conferring adaptation to potato plant allelochemicals and resistance to imidacloprid in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2017, 83, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kewley, R.J.; Whitelaw, M.L.; Chapman-Smith, A. The mammalian basic helix-loop-helix/PAS family of transcriptional regulators. Int. J. Biochem. Cell. Biol. 2004, 36, 189–204. [Google Scholar] [CrossRef]
- Mulero-Navarro, S.; Fernandez-Salguero, P.M. New Trends in Aryl Hydrocarbon Receptor Biology. Front. Cell. Dev. Biol. 2016, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Kann, S.; Huang, M.Y.; Estes, C.; Reichard, J.F.; Sartor, M.A.; Xia, Y.; Puga, A. Arsenite-induced aryl hydrocarbon receptor nuclear translocation results in additive induction of phase I genes and synergistic induction of phase II genes. Mol. Pharmacol. 2005, 68, 336–346. [Google Scholar] [CrossRef]
- Hu, B.; Huang, H.; Wei, Q.; Ren, M.; Mburu, D.K.; Tian, X.; Su, J. Transcription factors CncC/Maf and AhR/ARNT coordinately regulate the expression of multiple GSTs conferring resistance to chlorpyrifos and cypermethrin in Spodoptera exigua. Pest Manag. Sci. 2019, 75, 2009–2019. [Google Scholar] [CrossRef]
- Zhang, X.; Jie, D.; Liu, J.; Zhang, J.; Zhang, T.; Zhang, J.; Ma, E. Aryl hydrocarbon receptor regulates the expression of LmGSTd7 and is associated with chlorpyrifos susceptibility in Locusta migratoria. Pest Manag. Sci. 2019, 75, 2916–2924. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef]
- Samra, A.I.; Kamita, S.G.; Yao, H.W.; Cornel, A.J.; Hammock, B.D. Cloning and characterization of two glutathione S-transferases from pyrethroid-resistant Culex pipiens. Pest Manag. Sci. 2012, 68, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Liu, J.Y.; Rashid, M.; Wang, D.; Zhang, Y.L. Cantharidin Impedes Activity of Glutathione S-Transferase in the Midgut of Helicoverpa armigera Hubner. Int. J. Mol. Sci. 2013, 14, 5482–5500. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Jia, M.; Liu, T.; Xuan, T.; Yan, Z.K.; Guo, Y.; Ma, E.; Zhang, J. Identification and characterisation of ten glutathione S-transferase genes from oriental migratory locust, Locusta migratoria manilensis (Meyen). Pest Manag. Sci. 2011, 67, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Luo, Q.H.; Wang, J.M.; Li, X.F.; Yang, Y. Functional characterization and analysis of the Arabidopsis UGT71C5 promoter region. Genet. Mol. Res. 2015, 14, 19173–19183. [Google Scholar] [CrossRef] [PubMed]
- Kluge, J.; Terfehr, D.; Kuck, U. Inducible promoters and functional genomic approaches for the genetic engineering of filamentous fungi. Appl. Microbiol. Biotechnol. 2018, 102, 6357–6372. [Google Scholar] [CrossRef]
- Concha, C.; Edman, R.M.; Belikoff, E.J.; Schiemann, A.H.; Carey, B.; Scott, M.J. Organization and expression of the Australian sheep blowfly (Lucilia cuprina) hsp23, hsp24, hsp70 and hsp83 genes. Insect Mol. Biol. 2012, 21, 169–180. [Google Scholar] [CrossRef]
- Pellegrineschi, A.; Reynolds, M.; Pacheco, M.; Brito, R.M.; Almeraya, R.; Yamaguchi-Shinozaki, K.; Hoisington, D. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 2004, 47, 493–500. [Google Scholar] [CrossRef]
- Cao, M.Y.; Kuang, X.X.; Li, H.Q.; Lei, X.J.; Xiao, W.F.; Dong, Z.Q.; Zhang, J.; Hu, N.; Chen, T.T.; Lu, C.; et al. Screening and optimization of an efficient Bombyx mori nucleopolyhedrovirus inducible promoter. J. Biotechnol. 2016, 231, 72–80. [Google Scholar] [CrossRef]
- Huang, X.; Li, J.; Song, S.; Wang, L.; Lin, Z.; Ouyang, Z.; Yu, R. Hormesis effect of hydrogen peroxide on the promoter activity of neuropeptide receptor PAC1-R. J. Food Biochem. 2019, 43, e12877. [Google Scholar] [CrossRef]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Nakata, K.; Tanaka, Y.; Nakano, T.; Adachi, T.; Tanaka, H.; Kaminuma, T.; Ishikawa, T. Nuclear receptor-mediated transcriptional regulation in Phase I, II, and III xenobiotic metabolizing systems. Drug Metab. Pharm. 2006, 21, 437–457. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; O’Connor, T.; Katsuoka, F.; Engel, J.D.; Yamamoto, M. Integration and diversity of the regulatory network composed of Maf and CNC families of transcription factors. Gene 2002, 294, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Yamamoto, M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv. Enzyme. Regul. 2006, 46, 113–140. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, J.; Wang, H.; Mao, T.; Li, J.; Cheng, X.; Hu, J.; Xue, B.; Li, B. Cloning and Functional Analysis of CncC and Keap1 Genes in Silkworm. J. Agric. Food Chem. 2018, 66, 2630–2636. [Google Scholar] [CrossRef] [PubMed]
- Sykiotis, G.P.; Bohmann, D. Keap1/Nrf2 signaling regulates oxidative stress tolerance and lifespan in Drosophila. Dev. Cell 2008, 14, 76–85. [Google Scholar] [CrossRef]
- Wan, H.; Liu, Y.; Li, M.; Zhu, S.; Li, X.; Pittendrigh, B.R.; Qiu, X. Nrf2/Maf-binding-site-containing functional Cyp6a2 allele is associated with DDT resistance in Drosophila melanogaster. Pest Manag. Sci. 2014, 70, 1048–1058. [Google Scholar] [CrossRef]
- Kalsi, M.; Palli, S.R. Transcription factors, CncC and Maf, regulate expression of CYP6BQ genes responsible for deltamethrin resistance in Tribolium castaneum. Insect Biochem. Mol. Biol. 2015, 65, 47–56. [Google Scholar] [CrossRef]
- Vorrink, S.U.; Domann, F.E. Regulatory crosstalk and interference between the xenobiotic and hypoxia sensing pathways at the AhR-ARNT-HIF1alpha signaling node. Chem. Biol. Interact. 2014, 218, 82–88. [Google Scholar] [CrossRef]
- Jin, H.; Ji, C.; Ren, F.; Aniagu, S.; Tong, J.; Jiang, Y.; Chen, T. AHR-mediated oxidative stress contributes to the cardiac developmental toxicity of trichloroethylene in zebrafish embryos. J. Hazard. Mater. 2020, 385, 121521. [Google Scholar] [CrossRef]
- Wang, L.; He, X.; Szklarz, G.D.; Bi, Y.; Rojanasakul, Y.; Ma, Q. The aryl hydrocarbon receptor interacts with nuclear factor erythroid 2-related factor 2 to mediate induction of NAD(P)H:quinoneoxidoreductase 1 by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Arch. Biochem. Biophys. 2013, 537, 31–38. [Google Scholar] [CrossRef]
- Rushmore, T.H.; King, R.G.; Paulson, K.E.; Pickett, C.B. Regulation of glutathione S-transferase Ya subunit gene expression: Identification of a unique xenobiotic-responsive element controlling inducible expression by planar aromatic compounds. Proc. Natl. Acad. Sci. USA 1990, 87, 3826–3830. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, C. Antioxidant Functions of the Aryl Hydrocarbon Receptor. Stem Cells Int. 2016, 2016, 7943495. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Chen, X.; Pan, Y.; Zheng, Z.; Wei, X.; Xi, J.; Zhang, J.; Gao, X.; Shang, Q. Transcription factor aryl hydrocarbon receptor/aryl hydrocarbon receptor nuclear translocator is involved in regulation of the xenobiotic tolerance-related cytochrome P450 CYP6DA2 in Aphis gossypii Glover. Insect Mol. Biol. 2017, 26, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Peng, T.; Xu, P.; Zeng, X.; Tian, F.; Song, J.; Shang, Q. Transcription Factors AhR/ARNT Regulate the Expression of CYP6CY3 and CYP6CY4 Switch Conferring Nicotine Adaptation. Int. J. Mol. Sci. 2019, 20, 4521, Published 12 September 2019. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, R.; Liu, C.; Gao, Y.; Deng, X.; Wan, H.; Li, J. Functional characterization of the transcription factors AhR and ARNT in Nilaparvata lugens. Pestic. Biochem. Physiol. 2021, 176, 104875. [Google Scholar] [CrossRef]
- Kohle, C.; Bock, K.W. Coordinate regulation of Phase I and II xenobiotic metabolisms by the Ah receptor and Nrf2. Biochem. Pharmacol. 2007, 73, 1853–1862. [Google Scholar] [CrossRef]
- Li, F.; Ma, K.; Chen, X.; Zhou, J.J.; Gao, X. The regulation of three new members of the cytochrome P450 CYP6 family and their promoters in the cotton aphid Aphis gossypii by plant allelochemicals. Pest Manag. Sci. 2019, 75, 152–159. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Forward Primer Sequences (5’-3′) | Reverse Primer Sequences (5’-3′) | Experiments |
|---|---|---|---|
| Pro-MtGSTd2 (−642/+290) | CTCGAGAGATTACTATAGGGCACG | GCTAGCAGGTAGTACAGGTCG | 5’ loss fragment MtGST promoters |
| Pro-MtGSTd2 (−402/+290) | CTCGAGCTTAGCTGGTTGC | GCTAGCAGGTAGTACAGGTCG | |
| Pro-MtGSTo1 (−596/+414) | CTCGAGATTACTATAGGGCACGC | GCTAGCGGTTTGTAAATGTTTT | |
| Pro-MtGSTo1 (−360/+414) | CTCGAGTAACAATTGGCAC | GCTAGCGAATACACGCAGTTT | |
| Pro-MtGSTo1 (−127/+414) | CTCGAGTCCGACTTTGTGAA | GCTAGCGAATACACGCAGTTT | |
| Pro-MtGSTs1 (−576/+385) | CTCGAGCGACGAAGGCTT | GCTAGCTCAGTAACAACGAC | |
| Pro-MtGSTs1 (−361/+385) | CTCGAGCCTTCCAGTAGTTTG | GCTAGCTCAGTAACAACGAC | |
| Pro-MtGSTs1 (−70/+385) | CTCGAGCATCGTTTCTAGAGT | GCTAGCTCAGTAACAACGAC | |
| Pro-MtGSTt1 (−1552/+340) | CTCGAGTGCCTGCAGGTC | GCTAGCGATTTCTTCACAGAGTG | |
| Pro-MtGSTt1 (−1055/+340) | CTCGAGTGTCCCGTCACA | GCTAGCGATTTCTTCACAGAGTG | |
| Pro-MtGSTt1 (−574/+340) | CTCGAGTTGGACTATAGCCTTC | GCTAGCGATTTCTTCACAGAGTG | |
| Pro-MtGSTt1 (−334/+340) | CTCGAGCATGCTATGCCC | GCTAGCGATTTCTTCACAGAGTG | |
| Pro-MtGSTz1 (−500/+521) | CTCGAGGGCACGCGTG | GCTAGCAGCGATAGATAAGCG | |
| Pro-MtGSTz1 (−350/+521) | CTCGAGGACGTTGGCATT | GCTAGCAGCGATAGATAAGCG | |
| Pro-MtGSTz1 (−220/+521) | CTCGAGTCTTATTTGGAAACG | GCTAGCAGCGATAGATAAGCG | |
| ARNT | ATGAGTTTATTGACTGATGTCTGCCT | TCACCGGCGCCCGCCGCC | Gene cloning |
| CncC | ATGCTGCACCCGGCCAT | TCACTGATCGTAGTGCTTCGCTT | |
| pAC-V5-ARNT | GCGGCCGCATGAGTTTATTGACTGAT | GGCGCGCCCCGGCGCCCGCCGCC | pAC-V5 vector construct |
| pAC-V5-CncC | GCGGCCGCATGCTGCACCCGGCCAT | GGCGCGCCCTGATCGTAGTGCTTCGCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Shi, X.; Zhou, Y.; Tang, F.; Gao, X.; Liang, P. Cap ‘n’ Collar C and Aryl Hydrocarbon Receptor Nuclear Translocator Facilitate the Expression of Glutathione S-Transferases Conferring Adaptation to Tannic Acid and Quercetin in Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae). Int. J. Mol. Sci. 2023, 24, 2190. https://doi.org/10.3390/ijms24032190
Wang Z, Shi X, Zhou Y, Tang F, Gao X, Liang P. Cap ‘n’ Collar C and Aryl Hydrocarbon Receptor Nuclear Translocator Facilitate the Expression of Glutathione S-Transferases Conferring Adaptation to Tannic Acid and Quercetin in Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae). International Journal of Molecular Sciences. 2023; 24(3):2190. https://doi.org/10.3390/ijms24032190
Chicago/Turabian StyleWang, Zhiqiang, Xiaoling Shi, Yujingyun Zhou, Fang Tang, Xiwu Gao, and Pei Liang. 2023. "Cap ‘n’ Collar C and Aryl Hydrocarbon Receptor Nuclear Translocator Facilitate the Expression of Glutathione S-Transferases Conferring Adaptation to Tannic Acid and Quercetin in Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae)" International Journal of Molecular Sciences 24, no. 3: 2190. https://doi.org/10.3390/ijms24032190
APA StyleWang, Z., Shi, X., Zhou, Y., Tang, F., Gao, X., & Liang, P. (2023). Cap ‘n’ Collar C and Aryl Hydrocarbon Receptor Nuclear Translocator Facilitate the Expression of Glutathione S-Transferases Conferring Adaptation to Tannic Acid and Quercetin in Micromelalopha troglodyta (Graeser) (Lepidoptera: Notodontidae). International Journal of Molecular Sciences, 24(3), 2190. https://doi.org/10.3390/ijms24032190