


Sodium Nitroprusside Improves Bamboo Resistance under Mn and Cr Toxicity with Stimulation of Antioxidants Activity, Relative Water Content, and Metal Translocation and Accumulation

 ,
,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Mn and Cr Accumulation in Root, Stem, and Leaves in Bamboo Species
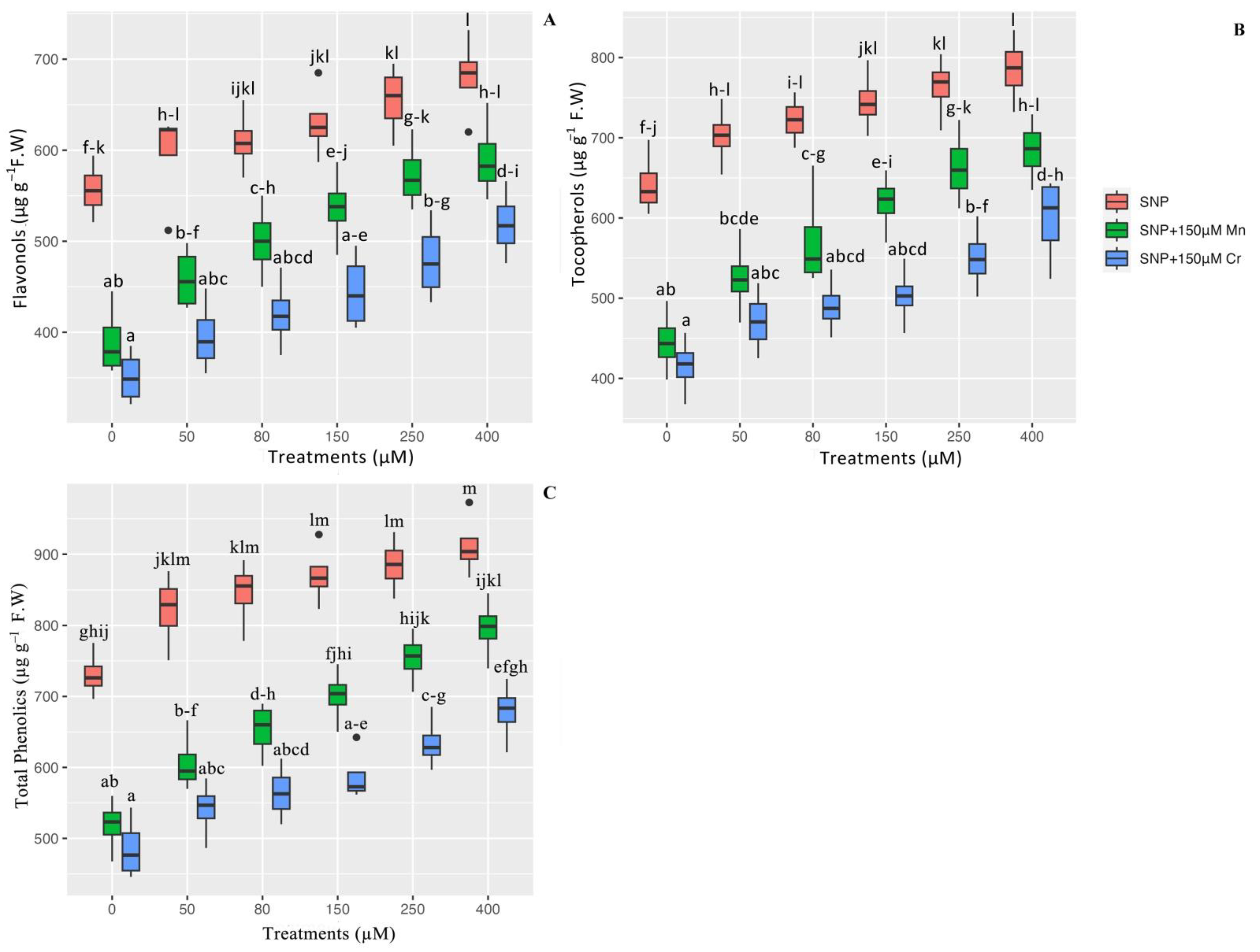
2.2. Tocopherols, Flavonols, and Total Phenolics in Plants under Mn and Cr
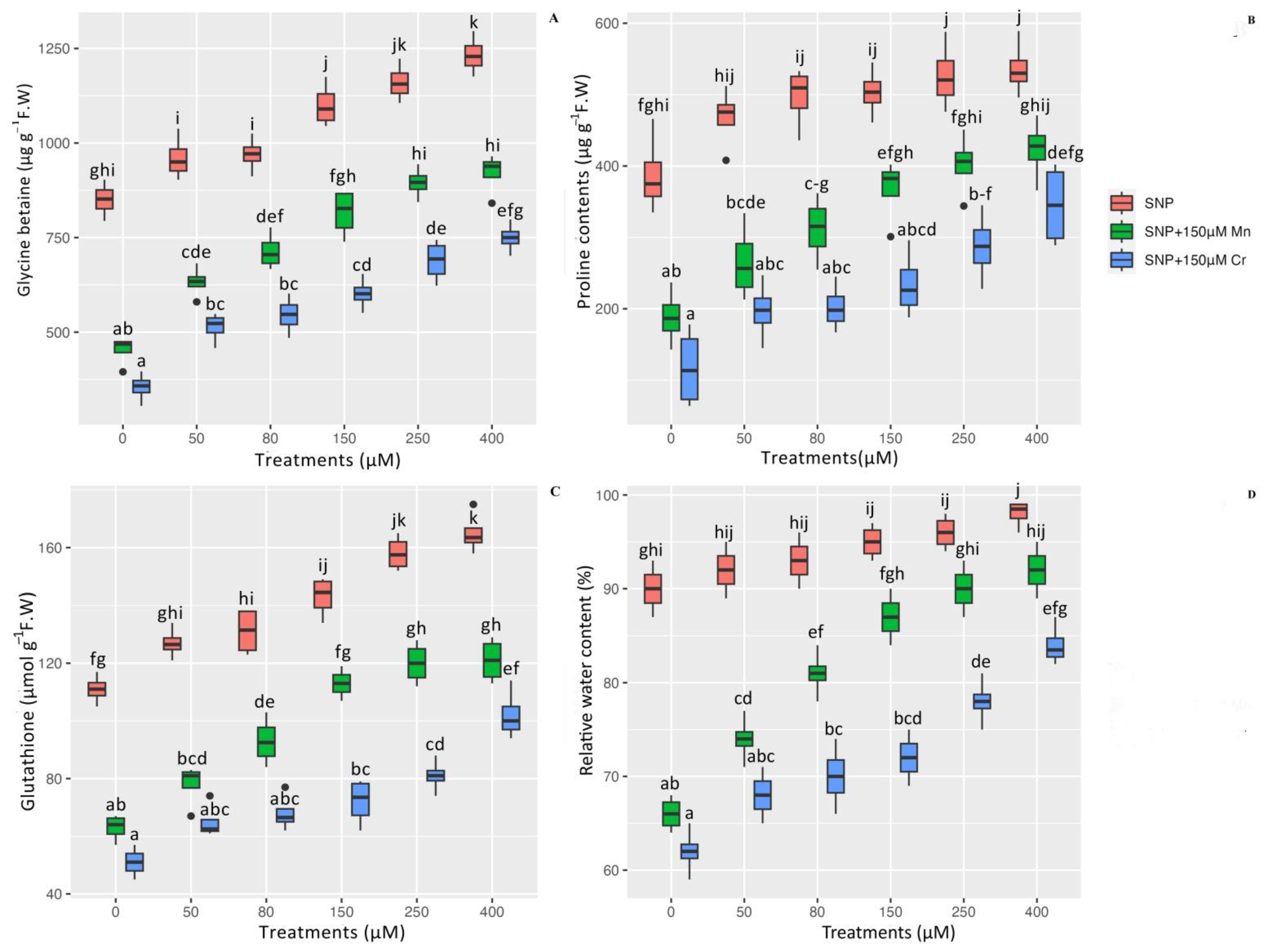
2.3. Proline Contents (Pro), Glycine Betaine (GB), Glutathione (GSH), and Relative Water Content (RWC) in Pleioblastus pygmaea L. under Mn and Cr
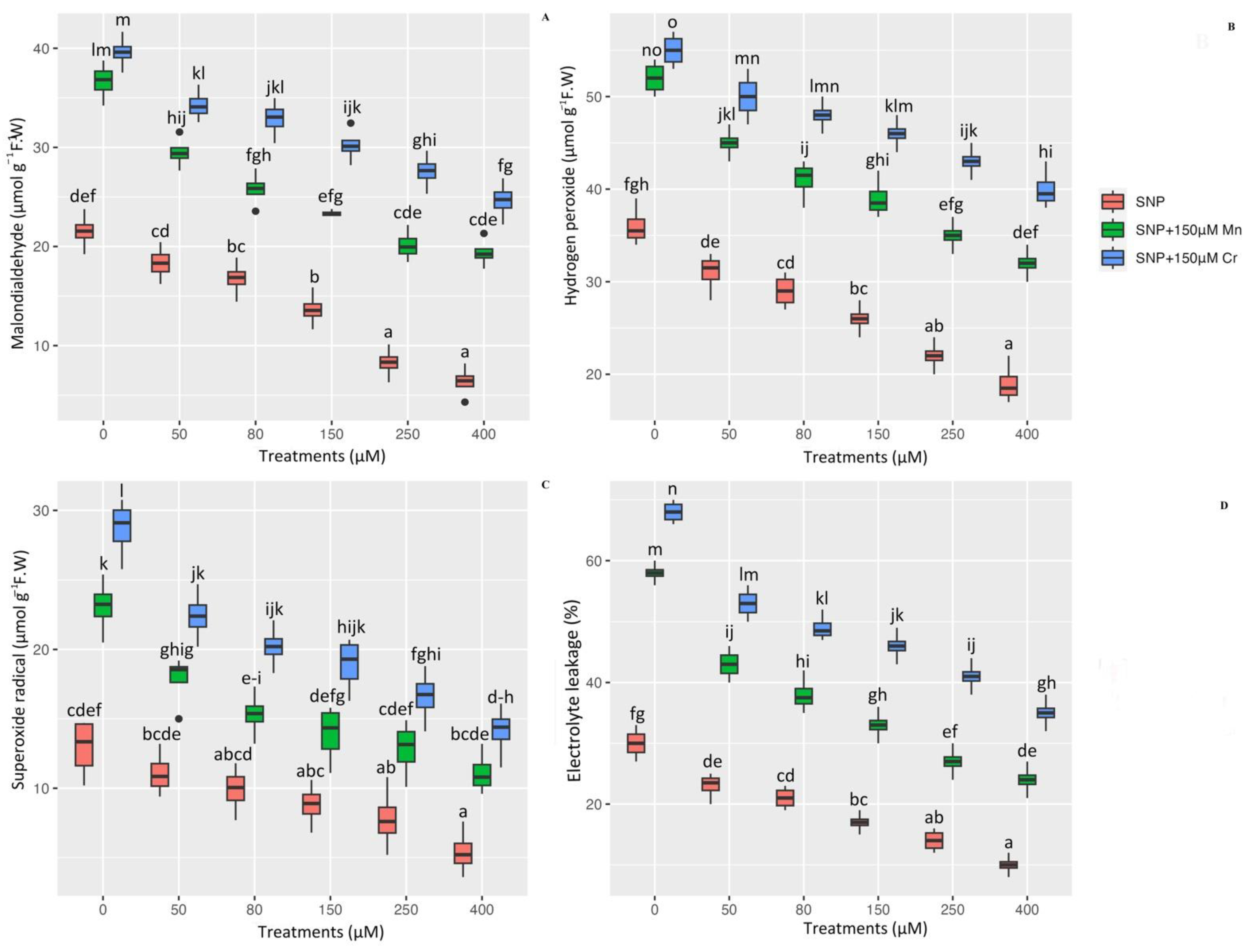
2.4. ROS Component and Amelioration Lipid Peroxidation in Pleioblastus pygmaea L. under Mn and Cr Stress
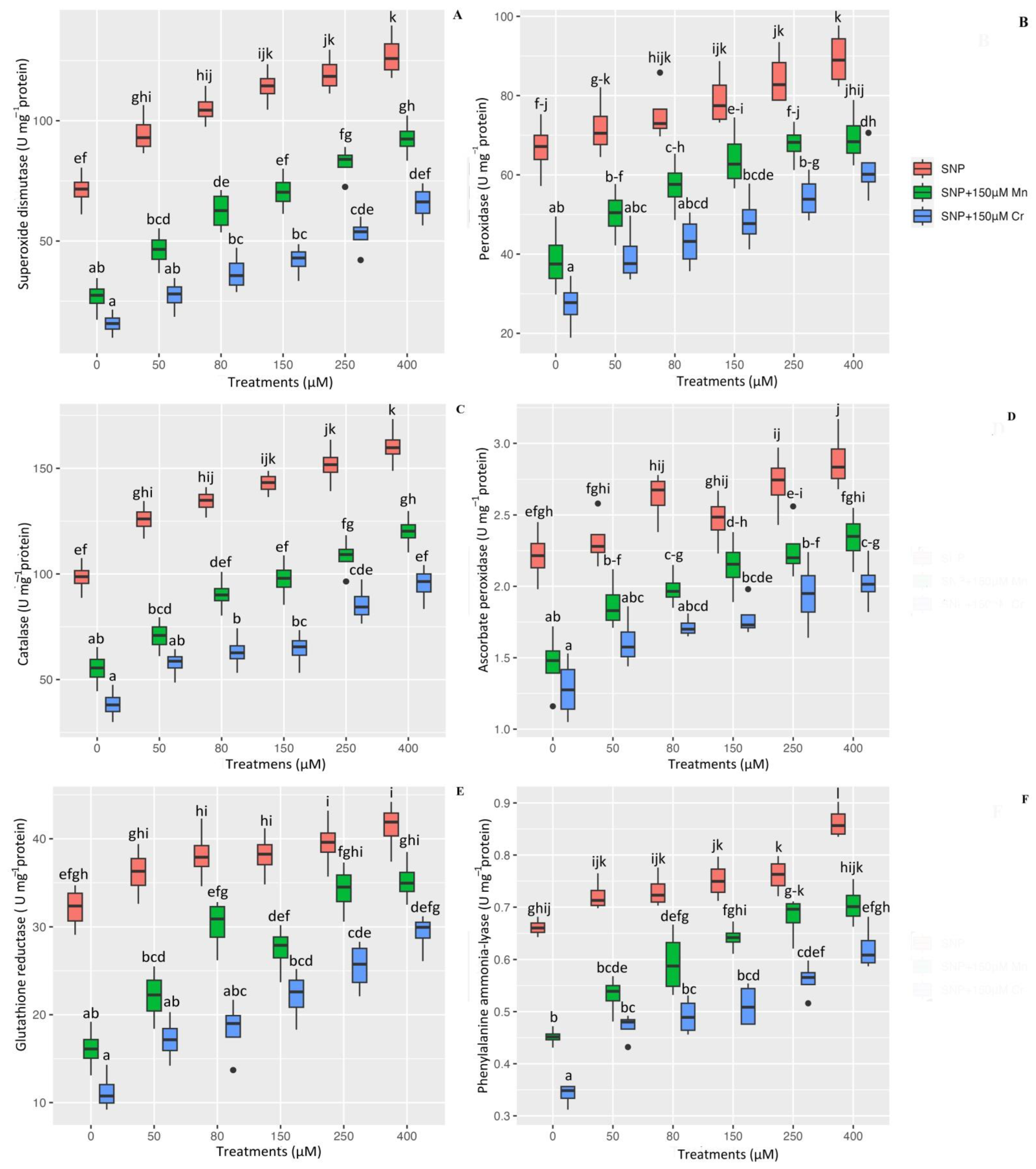
2.5. Antioxidant Enzyme Activity in Plants Exposed to Mn and Cr Toxicity
2.6. Plant Photosynthetic Pigments, including Chlorophyll a, Chlorophyll b, Total Chlorophyll and Carotenoids Exposed to Mn and Cr
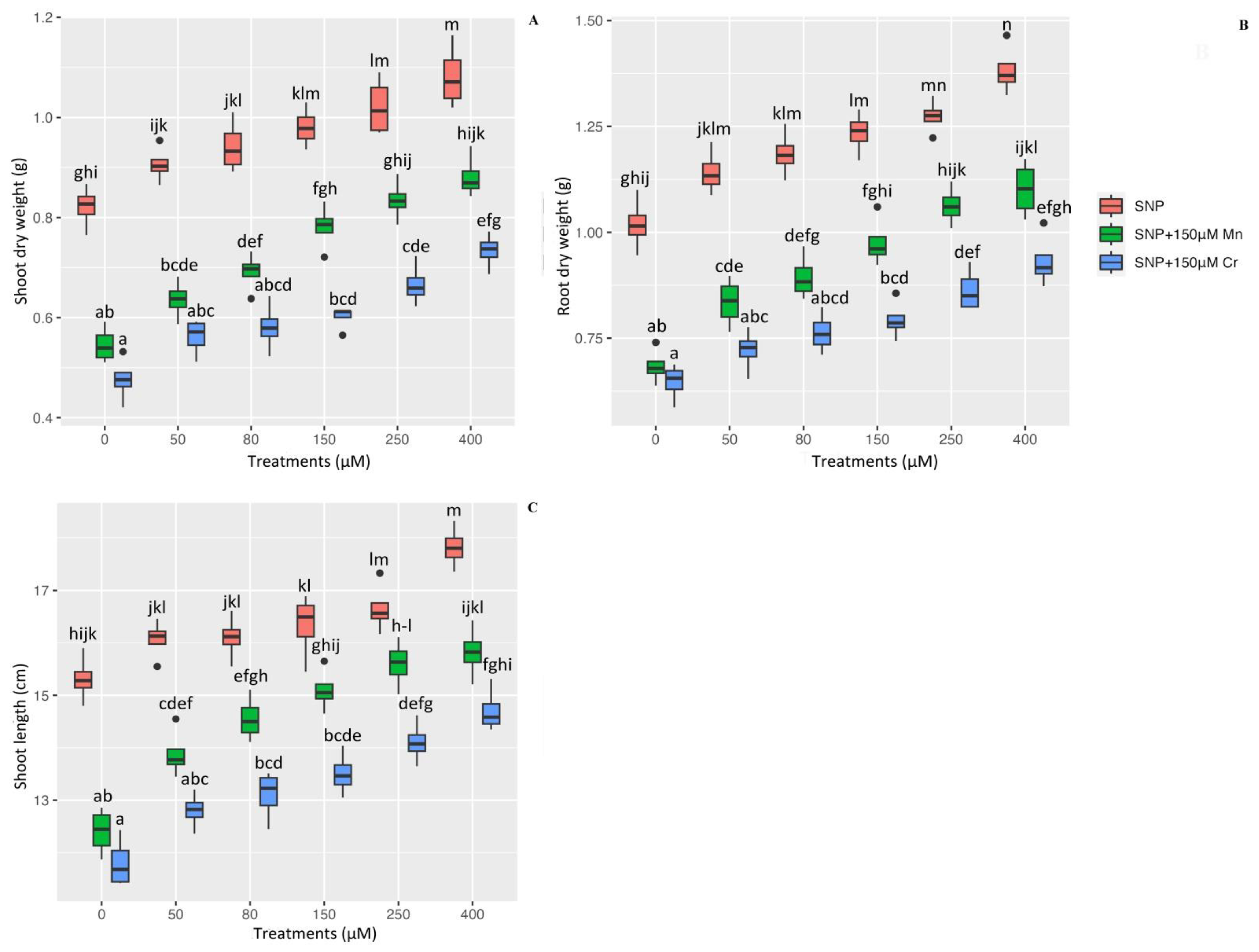
2.7. Plant Growth, and Plant Biomass on the Dry Weight of Shoot and Root as Well as Plant Shoot Length under Mn and Cr
2.8. Effects of Nitric Oxide on Plant Tolerance Indices (TI) in Shoot and Root, as Well as Bio Accumulation Factor (BAF) and Plant Translocation Factor (TF) in Pleioblastus pygmaea L. Exposed to Mn and Cr Toxicity
3. Discussion
4. Materials and Methods
4.1. Experiment Materials and Vitro Plant Tissue Culture
4.2. Estimation of Metal Content and Nitric Oxide Accumulation in Roots, Stems, and Leaves
4.3. Determination of Tocopherols, Flavonols, and Total Phenolics as Non-Antioxidant Activity
4.4. Relative Water Content (RWC), Proline Content (Pro), Glutathione (GSH), and Glycine Betaine (GB)
4.5. Lipid Peroxidation (MDA), Hydrogen Peroxide (H2O2), Electrolyte Leakage (EL), and Superoxide Radical (O2•−)
4.6. Antioxidant Activity
4.7. Photosynthetic Pigments including Chlorophyll a, Chlorophyll b, Total Chlorophyll, and Carotenoids Content
Chlorophyll a = 12.25A663 − 2.79A647
Chlorophyll b = 21.50A647 − 5.10A663
Carotenoid = (1000A470 − 1.82Chl a − 95.15 Chl b)/225
4.8. The Calculation of Tolerance Index (TI) in shoot and Root, The Translocation Factor (TF) in Leaves and Stem, as Well as Bioaccumulation Factor (BAF) in Root, Stem, and Leaves
4.9. Root Dry Weight, Shoot Dry Weight, and Shoot Length
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ekmekci, Y.; Bohms, A.; Thomson, J.A.; Mundree, S.G. Photochemical and Antioxidant Responses in the Leaves of Xerophyta viscosa Baker and Digitaria sanguinalis L. under Water Deficit. Z. Naturforsch. C J. Biosci. 2005, 60, 435–443. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.-G.; Yun, B.-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Huang, Y.L.; Yang, S.; Long, G.X.; Zhao, Z.K.; Li, X.F.; Gu, M.H. Manganese Toxicity in Sugarcane Plantlets Grown on Acidic Soils of Southern China. PLoS ONE 2016, 11, e0148956. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-Ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Fernando, D.R.; Lynch, J.P. Manganese phytotoxicity: New light on an old problem. Ann. Bot. 2015, 116, 313–319. [Google Scholar] [CrossRef]
- Kumar, R.; Alamelu, D.; Acharya, R.; Rai, A.K. Determination of concentrations of chromium and other elements in soil and plant samples from leather tanning area by Instrumental Neutron Activation Analysis. J. Radioanal. Nucl. Chem. 2014, 300, 213–218. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Gao, Y.; Xia, J. Chromium Contamination Accident in China: Viewing Environment Policy of China. Environ. Sci. Technol. 2011, 45, 8605–8606. [Google Scholar] [CrossRef]
- State Ministry of Environmental Protection (MEP). Chromium Slays Pollution Control and Environmental Protection Technical Specifications. Rep. State Environ. China 2007. [Google Scholar]
- Domingos, P.; Prado, A.M.; Wong, A.; Gehring, C.; Feijo, J.A. Nitric Oxide: A Multitasked Signaling Gas in Plants. Mol. Plant 2015, 8, 506–520. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Liu, L.; Guo, Y.; Ren, H. Alleviating effect of exogenous nitric oxide in cucumber seedling against chilling stress. Afr. J. Biotechnol. 2011, 10, 4380–4386. [Google Scholar]
- Hao, G.-P.; Zhang, J.-H. The role of nitric oxide as a bioactive signaling molecule in plants under abiotic stress. In Nitric oxide in Plant Physiology; Hayat, S., Mori, M., Pichtel, J., Ahmad, A., Eds.; Wiley: Weinheim, Germany, 2010; pp. 115–138. [Google Scholar] [CrossRef]
- Jaspers, P.; Kangasjärvi, J. Reactive oxygen species in abiotic stress signaling. Physiol. Plant. 2010, 138, 405–413. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef]
- Vranová, E.; Atichartpongkul, S.; Villarroel, R.; Van Montagu, M.; Inzé, D.; Van Camp, W. Comprehensive analysis of gene expression in Nicotiana tabacum leaves acclimated to oxidative stress. Proc. Natl. Acad. Sci. USA 2002, 99, 10870–10875. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, J.; Wei, S.; Hu, D.; Liu, Y.; Feng, L.; Li, C.; Qi, N.; Wang, C.; Liao, W. Nitric Oxide Enhanced Salt Stress Tolerance in Tomato Seedlings, Involving Phytohormone Equilibrium and Photosynthesis. Int. J. Mol. Sci. 2022, 23, 4539. [Google Scholar] [CrossRef]
- Hasanuzzam, M.; Hoss, M.A.; Fujita, M. Physiological and Biochemical Mechanisms of Nitric Oxide Induced Abiotic Stress Tolerance in Plants. Am. J. Plant Physiol. 2010, 5, 295–324. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Gill, S.S.; Fujita, M. Physiological role of nitric oxide in plants grown under adverse environmental conditions. In Plant Acclimation to Environmental Stress; Tuteja, N., Gill, S.S., Eds.; Springer: New York, NY, USA, 2013; pp. 169–322. [Google Scholar] [CrossRef]
- Gupta, K.J.; Igamberdiev, A.U.; Manjunatha, G.; Segu, S.; Moran, J.F.; Neelawarne, B.; Bauwe, H.; Kaiser, W.M. The emerging roles of nitric oxide (NO) in plant mitochondria. Plant Sci. 2011, 181, 520–526. [Google Scholar] [CrossRef]
- De Michele, R.; Vurro, E.; Rigo, C.; Costa, A.; Elviri, L.; Di Valentin, M.; Careri, M.; Zottini, M.; Sanita di Toppi, L.; Lo Schiavo, F. Nitric Oxide Is Involved in Cadmium-Induced Programmed Cell Death in Arabidopsis Suspension Cultures. Plant Physiol. 2009, 150, 217–228. [Google Scholar] [CrossRef]
- Lipton, S.A.; Choi, Y.-B.; Pan, Z.-H.; Lei, S.Z.; Chen, H.-S.V.; Sucher, N.J.; Loscalzo, J.; Singel, D.J.; Stamler, J.S. A redox-based mechanism for the neuroprotective and neurodestructive effects of nitric oxide and related nitroso-compounds. Nature 1993, 364, 626–632. [Google Scholar] [CrossRef]
- Corpas, F.J.; Leterrier, M.; Valderrama, R.; Airaki, M.; Chaki, M.; Palma, J.M.; Barroso, J.B. Nitric oxide imbalance provokes a nitrosative response in plants under abiotic stress. Plant Sci. 2011, 181, 604–611. [Google Scholar] [CrossRef]
- Planchet, E.; Kaiser, W.M. Nitric Oxide Production in Plants. Plant Signal. Behav. 2006, 1, 46–51. [Google Scholar] [CrossRef]
- Planchet, E. Nitric oxide (NO) detection by DAF fluorescence and chemiluminescence: A comparison using abiotic and biotic NO sources. J. Exp. Bot. 2006, 57, 3043–3055. [Google Scholar] [CrossRef]
- Souri, Z.; Karimi, N.; Farooq, M.A.; Sandalio, L.M. Nitric oxide improves tolerance to arsenic stress in Isatis cappadocica desv. Shoots by enhancing antioxidant defenses. Chemosphere 2019, 239, 124523. [Google Scholar] [CrossRef]
- Souri, Z.; Karimi, N.; Farooq, M.A.; Lobato, A.K.S. Improved physiological defense responses by application of sodium nitroprusside in Isatis cappadocica Desv. under cadmium stress. Physiol. Plant. 2021, 173, 100–115. [Google Scholar] [CrossRef]
- Jabeen, Z.; Fayyaz, H.A.; Irshad, F.; Hussain, N.; Hassan, M.N.; Li, J.; Rehman, S.; Haider, W.; Yasmin, H.; Mumtaz, S.; et al. Sodium nitroprusside application improves morphological and physiological attributes of soybean (Glycine max L.) under salinity stress. PLoS ONE 2021, 16, e0248207. [Google Scholar] [CrossRef]
- Emamverdian, Y.; Ding, Y.; Xie, Y. Phytoremediation potential of bamboo plant in China. Ecol. Environ. Conserv. 2018, 24, 530–539. [Google Scholar]
- Ahmad, Z.; Upadhyay, A.; Ding, Y.; Emamverdian, A.; Shahzad, A. Bamboo: Origin, habitat, distributions and global prospective. In Biotechnological Advances in Bamboo; Ahmad, Z., Ding, Y., Shahzad, A., Eds.; Springer: Singapore, 2021; pp. 1–31. [Google Scholar]
- Emamverdian, A.; Ding, Y.; Ranaei, F.; Ahmad, Z. Application of Bamboo Plants in Nine Aspects. Sci. World J. 2020, 2020, 7284203. [Google Scholar] [CrossRef]
- Huang, Z.; Jin, S.-H.; Guo, H.-D.; Zhong, X.-J.; He, J.; Li, X.; Jiang, M.-Y.; Yu, X.-F.; Long, H.; Ma, M.-D.; et al. Genome-wide identification and characterization of TIFY family genes in Moso Bamboo (Phyllostachys edulis) and expression profiling analysis under dehydration and cold stresses. Peerj 2016, 4, e2620. [Google Scholar] [CrossRef]
- Jin, Q.-Y.; Peng, H.-Z.; Lin, E.-P.; Li, N.; Huang, D.-N.; Xu, Y.-L.; Hua, X.-Q.; Wang, K.-H.; Zhu, T.-J. Identification and characterization of differentially expressed miRNAs between bamboo shoot and rhizome shoot. J. Plant Biol. 2016, 59, 322–335. [Google Scholar] [CrossRef]
- Hogarth, N.; Belcher, B. The contribution of bamboo to household income and rural livelihoods in a poor and mountainous county in Guangxi, China. Int. For. Rev. 2013, 15, 71–81. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Zhang, Y.; Booth, T.; He, X. Changes of carbon stocks in bamboo stands in China during 100 years. For. Ecol. Manag. 2009, 258, 1489–1496. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, Y.; He, T.; You, L.; Shen, X. Assessment of Potential Capability of Water Bamboo Leaves on the Adsorption Removal Efficiency of Cationic Dye from Aqueous Solutions. J. Polym. Environ. 2016, 24, 148–158. [Google Scholar] [CrossRef]
- State Forestry Administration of China. Statistical Yearbook of Forestry; State Forestry Administration of China: Beijing, China, 2012.
- Liu, J.-N.; Zhou, Q.-X.; Sun, T.; Ma, L.Q.; Wang, S. Growth responses of three ornamental plants to Cd and Cd–Pb stress and their metal accumulation characteristics. J. Hazard. Mater. 2008, 151, 261–267. [Google Scholar] [CrossRef]
- Huang, W.; Olson, E.; Wang, S.; Shi, P. The growth and mortality of Pleioblastus pygmaeus under different light availability. Glob. Ecol. Conserv. 2020, 24, e01262. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Ramakrishnan, M.; Ahmad, Z.; Xie, Y. Different Physiological and Biochemical Responses of Bamboo to the Addition of TiO2 NPs under Heavy Metal Toxicity. Forests 2021, 12, 759. [Google Scholar] [CrossRef]
- RunHong, M.; Cheng, J.; Tang, F.; Yue, J.; Li, Z.; Ni, Z. Heavy metals in bamboo shoots from Southeastern China and risk assessment. Food Addit. Contam. Part B 2021, 14, 264–270. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Batish, D.R.; Sharma, V.P.; Sharma, N.; Kohli, R.K. Nitric oxide alleviates arsenic toxicity by reducing oxidative damage in the roots of Oryza sativa (rice). Nitric Oxide 2009, 20, 289–297. [Google Scholar] [CrossRef]
- Ismail, G.S.M. Protective role of nitric oxide against arsenic-induced damages in germinating mung bean seeds. Acta Physiol. Plant. 2012, 34, 1303–1311. [Google Scholar] [CrossRef]
- Verma, K.; Mehta, S.K.; Shekhawat, G.S. Nitric oxide (NO) counteracts cadmium induced cytotoxic processes mediated by reactive oxygen species (ROS) in Brassica juncea: Cross-talk between ROS, NO and antioxidant responses. Biometals 2013, 26, 255–269. [Google Scholar] [CrossRef]
- Hsu, Y.T.; Kao, C.H. Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul. 2004, 42, 227–238. [Google Scholar] [CrossRef]
- Wink, D.A.; Hanbauer, I.; Krishna, M.C.; DeGraff, W.; Gamson, J.; Mitchell, J.B. Nitric oxide protects against cellular damage and cytotoxicity from reactive oxygen species. Proc. Natl. Acad. Sci. USA 1993, 90, 9813–9817. [Google Scholar] [CrossRef]
- Wendehenne, D.; Pugin, A.; Klessig, D.F.; Durner, J. Nitric oxide: Comparative synthesis and signaling in animal and plant cells. Trends Plant Sci. 2001, 6, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Zottini, M.; Formentin, E.; Scattolin, M.; Carimi, F.; Schiavo, F.L.; Terzi, M. Nitric oxide affects plant mitochondrial functionality in vivo. FEBS Lett. 2002, 515, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Ding, F.; Wang, X.; Wei, M. Exogenous nitric oxide protect cucumber roots against oxidative stress induced by salt stress. Plant Physiol. Biochem. 2007, 45, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, S.; Xuan, Y.; Sun, M.; Zhao, L. Protective Effect of Nitric Oxide against Oxidative Damage in Arabidopsis Leaves under Ultraviolet-B Irradiation. J. Plant Biol. 2009, 52, 135–140. [Google Scholar] [CrossRef]
- Sun, F.; Guo, G.; Du, J.; Guo, W.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. Whole-genome discovery of miRNAs and their targets in wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 142. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, Y.; Wang, M.; Li, T.; Zhou, Y.; Wang, X.; Wei, S.; He, G.; Yang, G. Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.). BMC Plant Biol. 2015, 15, 1–17. [Google Scholar] [CrossRef]
- Farnese, F.S.; de Oliveira, J.A.; Gusman, G.S.; Leão, G.A.; Ribeiro, C.; Siman, L.I.; Cambraia, J. Plant Responses to Arsenic: The Role of Nitric Oxide. Water Air Soil Pollut. 2013, 224, 1660. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef]
- Talukdar, D. Arsenic-induced oxidative stress in the common bean legume, Phaseolus vulgaris L. seedlings and its amelioration by exogenous nitric oxide. Physiol. Mol. Biol. Plants 2012, 19, 69–79. [Google Scholar] [CrossRef]
- Márquez-García, B.; Fernández-Recamales, M.; Córdoba, F. Effects of Cadmium on Phenolic Composition and Antioxidant Activities of Erica andevalensis. J. Bot. 2012, 2012, 936950. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd Allah, E.F.; Gucel, S.; Tran, L.S. Nitric Oxide Mitigates Salt Stress by Regulating Levels of Osmolytes and Antioxidant Enzymes in Chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Polish J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Jung, C.; Maeder, V.; Funk, F.; Frey, B.; Sticher, H.; Frossard, E. Release of phenols from Lupinus albus L. roots exposed to Cu and their possible role in Cu detoxification. Plant Soil 2003, 252, 301–312. [Google Scholar] [CrossRef]
- Simontacchi, M.; Galatro, A.; Ramos-Artuso, F.; Santa-María, G.E. Plant Survival in a Changing Environment: The Role of Nitric Oxide in Plant Responses to Abiotic Stress. Front. Plant Sci. 2015, 6, 977. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef]
- Tossi, V.; Amenta, M.; Lamattina, L.; Cassia, R. Retracted: Nitric oxide enhances plant ultraviolet-B protection up-regulating gene expression of the phenylpropanoid biosynthetic pathway. Plant. Cell Environ. 2011, 34, 909–921. [Google Scholar] [CrossRef]
- Dube, B.K.; Sinha, P.; Shukla, K.; Chatterjee, C.; Pandey, V.K.; Rai, A.D. Involvement of Excess Cadmium on Oxidative Stress and Other Physiological Parameters of Eggplant. J. Plant Nutr. 2009, 32, 996–1004. [Google Scholar] [CrossRef]
- Laloi, C.; Apel, K.; Danon, A. Reactive oxygen signalling: The latest news. Curr. Opin. Plant Biol. 2004, 7, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Desikan, R.; Hancock, J.T. Nitric oxide signalling in plants. New Phytol. 2003, 159, 11–35. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Singla-Pareek, S.L.; Sopory, S.K. An overview on the role of methylglyoxal and glyoxalases in plants. Drug Metab. Drug Interactions 2008, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Yadav, S.K. Proline and betaine provide protection to antioxidant and methylglyoxal detoxification systems during cold stress in Camellia sinensis (L.) O. Kuntze. Acta Physiol. Plant. 2008, 31, 261–269. [Google Scholar] [CrossRef]
- Innocenti, G.; Pucciariello, C.; Le Gleuher, M.; Hopkins, J.; de Stefano, M.; Delledonne, M.; Puppo, A.; Baudouin, E.; Frendo, P. Glutathione synthesis is regulated by nitric oxide in Medicago truncatula roots. Planta 2006, 225, 1597–1602. [Google Scholar] [CrossRef]
- El-Shabrawi, H.; Kumar, B.; Kaul, T.; Reddy, M.K.; Singla-Pareek, S.L.; Sopory, S.K. Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice. Protoplasma 2010, 245, 85–96. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop. Sci. 2012, 6, 1314–1323. [Google Scholar]
- Hasanuzzaman, M.; Fujita, M. Selenium Pretreatment Upregulates the Antioxidant Defense and Methylglyoxal Detoxification System and Confers Enhanced Tolerance to Drought Stress in Rapeseed Seedlings. Biol. Trace Element Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.; Sopory, S. Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 2005, 337, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Parvaiz, A.; Satyawati, S. Salt stress and phyto-biochemical responses of plants—A review. Plant Soil Environ. 2008, 54, 89–99. [Google Scholar] [CrossRef]
- Emamverdian, A.; Hasanuzzaman, M.; Ding, Y.; Barker, J.; Mokhberdoran, F.; Liu, G. Zinc Oxide Nanoparticles Improve Pleioblastus pygmaeus Plant Tolerance to Arsenic and Mercury by Stimulating Antioxidant Defense and Reducing the Metal Accumulation and Translocation. Front. Plant Sci. 2022, 13, 841501. [Google Scholar] [CrossRef]
- Rezayian, M.; Ebrahimzadeh, H.; Niknam, V. Nitric Oxide Stimulates Antioxidant System and Osmotic Adjustment in Soybean Under Drought Stress. J. Soil Sci. Plant Nutr. 2020, 20, 1122–1132. [Google Scholar] [CrossRef]
- Altaf, M.M.; Diao, X.-P.; Rehman, A.U.; Imtiaz, M.; Shakoor, A.; Younis, H.; Fu, P.; Ghani, M.U. Effect of Vanadium on Growth, Photosynthesis, Reactive Oxygen Species, Antioxidant Enzymes, and Cell Death of Rice. J. Soil Sci. Plant Nutr. 2020, 20, 2643–2656. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Zhao, H.; Jin, Q.; Wang, Y.; Chu, L.; Li, X.; Xu, Y. Effects of nitric oxide on alleviating cadmium stress in Typha angustifolia. Plant Growth Regul. 2015, 78, 243–251. [Google Scholar] [CrossRef]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide 2012, 27, 210–218. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef]
- Li, C.; Li, T.; Zhang, D.; Jiang, L.; Shao, Y. Exogenous nitric oxide effect on fructan accumulation and FBEs expression in chilling-sensitive and chilling-resistant wheat. Environ. Exp. Bot. 2013, 86, 2–8. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Moad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. In Water Stress and Crop Plants: A Sustainable Approach; Ahmad, P., Ed.; Wiley Blackwell: Hoboken, NJ, USA, 2016; pp. 649–668. [Google Scholar]
- Dong, Y.; Chen, W.; Xu, L.; Kong, J.; Liu, S.; He, Z. Nitric oxide can induce tolerance to oxidative stress of peanut seedlings under cadmium toxicity. Plant Growth Regul. 2015, 79, 19–28. [Google Scholar] [CrossRef]
- Khairy, A.I.H.; Oh, M.J.; Lee, S.M.; Kim, D.S.; Roh, K.S. Nitric oxide overcomes Cd and Cu toxicity in in vitro-grown tobacco plants through increasing contents and activities of rubisco and rubisco activase. Biochim. Open 2016, 2, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Sadeghipour, O.; Aghaei, P. Improving the growth of cowpea (Vigna unguiculata L. Walp.) by magnetized water. J. Biodivers. Environ. Sci. 2013, 3, 37–43. [Google Scholar]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Role of nitric oxide in tolerance of plants to abiotic stress. Protoplasma 2010, 248, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.I.; Latif, H.H.; Hanafy, R.S. Influence of Nitric Oxide Application on Some Biochemical Aspects, Endogenous Hormones, Minerals and Phenolic Compounds of Vicia faba Plant Grown under Arsenic Stress. Gesunde Pflanz. 2016, 68, 99–107. [Google Scholar] [CrossRef]
- Yang, L.; Ji, J.; Harris-Shultz, K.; Wang, H.; Wang, H.; Abd_Allah, E.; Luo, Y.; Hu, X. The Dynamic Changes of the Plasma Membrane Proteins and the Protective Roles of Nitric Oxide in Rice Subjected to Heavy Metal Cadmium Stress. Front. Plant Sci. 2016, 7, 190. [Google Scholar] [CrossRef]
- Seabra, A.B.; Oliveira, H.C. How nitric oxide donors can protect plants in a changing environment: What we know so far and perspectives. AIMS Mol. Sci. 2016, 3, 692–718. [Google Scholar] [CrossRef]
- Leshem, Y.Y.; Haramaty, E. The Characterization and Contrasting Effects of the Nitric Oxide Free Radical in Vegetative Stress and Senescence of Pisum sativum Linn. Foliage. J. Plant Physiol. 1996, 148, 258–263. [Google Scholar] [CrossRef]
- Yang, L.-T.; Qi, Y.-P.; Chen, L.-S.; Sang, W.; Lin, X.-J.; Wu, Y.-L.; Yang, C.-J. Nitric oxide protects sour pummelo (Citrus grandis) seedlings against aluminum-induced inhibition of growth and photosynthesis. Environ. Exp. Bot. 2012, 82, 1–13. [Google Scholar] [CrossRef]
- Singh, P.K.; Indoliya, Y.; Chauhan, A.S.; Singh, S.P.; Singh, A.P.; Dwivedi, S.; Tripathi, R.D.; Chakrabarty, D. Nitric oxide mediated transcriptional modulation enhances plant adaptive responses to arsenic stress. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Kopyra, M.; Gwóźdź, E.A. Nitric oxide stimulates seed germination and counteracts the inhibitory effect of heavy metals and salinity on root growth of Lupinus luteus. Plant Physiol. Biochem. 2003, 41, 1011–1017. [Google Scholar] [CrossRef]
- Dong, Y.; Xu, L.; Wang, Q.; Fan, Z.; Kong, J.; Bai, X. Effects of exogenous nitric oxide on photosynthesis, antioxidative ability, and mineral element contents of perennial ryegrass under copper stress. J. Plant Interact. 2013, 9, 402–411. [Google Scholar] [CrossRef]
- Dong, Y.; Jinc, S.; Liu, S.; Xu, L.; Kong, J. Effects of exogenous nitric oxide on growth of cotton seedlings under NaCl stress. J. Soil Sci. Plant Nutr. 2014, 14, 1–13. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Seraj, Z.; Fujita, M. Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings. Protoplasma 2014, 251, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Fu, G.; Tao, L.; Zhu, C. Roles of nitric oxide in alleviating heavy metal toxicity in plants. Arch. Biochem. Biophys. 2010, 497, 13–20. [Google Scholar] [CrossRef]
- Gill, S.S.; Hasanuzzaman, M.; Nahar, K.; Macovei, A.; Tuteja, N. Importance of nitric oxide in cadmium stress tolerance in crop plants. Plant Physiol. Biochem. 2013, 63, 254–261. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Gupta, D.K.; Archilla, A.; Romero-Puertas, M.C.; del Río, L.A. Reactive oxygen species and nitric oxide in plants under cadmium stress from toxicity to signaling. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 199–215. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Xiong, J.; An, L.; Lu, H.; Zhu, C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 2009, 230, 755–765. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Karimi, N.; Ghaderian, S.M.; Raab, A.; Feldmann, J.; Meharg, A.A. An arsenic-accumulating, hypertolerant brassica, Isatis capadocica. New Phytol. 2009, 184, 41–47. [Google Scholar] [CrossRef]
- Shukla, P.; Singh, A.K. Nitric oxide mitigates arsenic-induced oxidative stress and genotoxicity in Vicia faba L. Environ. Sci. Pollut. Res. 2015, 22, 13881–13891. [Google Scholar] [CrossRef]
- Murphy, M.E.; Noack, E. Nitric oxide assay using hemoglobin method. Methods Enzymol. 1994, 233, 240–250. [Google Scholar] [CrossRef]
- Conde, E.; Cadahía, E.; Garcia-Vallejo, M.C. HPLC analysis of flavonoids and phenolic acids and aldehydes in Eucalyptus spp. Chromatographia 1995, 41, 657–660. [Google Scholar] [CrossRef]
- Akkol, E.K.; Göger, F.; Koşar, M.; Başer, K.H.C. Phenolic composition and biological activities of Salvia halophila and Salvia virgata from Turkey. Food Chem. 2008, 108, 942–949. [Google Scholar] [CrossRef]
- Kayden, H.J.; Chow, C.-K.; Bjornson, L.K. Spectrophotometric method for determination of tocopherol in red blood cells. J. Lipid Res. 1973, 14, 533–540. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A Re-Examination of the Relative Turgidity Technique for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Rao, K.M.; Sresty, T. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Li, C.; Bai, T.; Ma, F.; Han, M. Hypoxia tolerance and adaptation of anaerobic respiration to hypoxia stress in two Malus species. Sci. Hortic. 2010, 124, 274–279. [Google Scholar] [CrossRef]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 186–191. [Google Scholar] [CrossRef]
- Zhang, X. The Measurement and Mechanism of Lipid Peroxidation and SOD, POD and CAT Activities in Biological System. In Research Methodology of Crop Physiology; Agriculture Press: Beijing, China, 1992. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar]
- Foster, J.G.; Hess, J.L. Responses of Superoxide Dismutase and Glutathione Reductase Activities in Cotton Leaf Tissue Exposed to an Atmosphere Enriched in Oxygen. Plant Physiol. 1980, 66, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Berner, M.; Krug, D.; Bihlmaier, C.; Vente, A.; Müller, R.; Bechthold, A. Genes and Enzymes Involved in Caffeic Acid Biosynthesis in the Actinomycete Saccharothrix espanaensis. J. Bacteriol. 2006, 188, 2666–2673. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV–VIS specroscopy. In Current Protocols in Food Analytical Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; p. 4. [Google Scholar] [CrossRef]
- Souri, Z.; Karimi, N. Enhanced Phytoextraction by As Hyperaccumulator Isatis cappadocica Spiked with Sodium Nitroprusside. Soil Sediment Contam. Int. J. 2017, 26, 457–468. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP Concentration | Mn, and Cr Contents | Mn and Cr Accumulation (Leaves) | SNP Accumulation (Leaves) | Mn and Cr Accumulation (Stem) | SNP Accumulation (Stem) | Mn and Cr Accumulation (Root) | SNP Accumulation (Root) |
|---|---|---|---|---|---|---|---|
| µmol L−1 | µmol L−1 | µg g−1 | µg g−1 | µg g−1 | µg g−1 | µg g−1 | µg g−1 |
| 0 | 0 | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| 0 | 150 µM Mn | 27.42 ± 1.04 i | 0 a | 39.50 ± 0.72 l | 0 a | 45.00 ± 0.74 k | 0 a |
| 0 | 150 µM Cr | 31.35 ± 0.97 j | 0 a | 43.52 ± 0.79 m | 0 a | 49.20 ± 0.84 l | 0 a |
| 50 µM | 0 | 0 a | 25.07 ± 0.74 j | 0 a | 31.2 ± 0.77 h | 0 a | 41.30 ± 0.73 jk |
| 50 µM | 150 µM Mn | 19.25 ± 0.87 f | 11.10 ± 0.69 d | 28.77 ± 0.67 h | 16.2 ± 0.83 g | 31.10 ± 0.91 g | 23.7 ± 0.87 de |
| 50 µM | 150 µM Cr | 25.35 ± 1.07 hi | 6.2 ± 0.71 b | 36.35 ±0.98 k | 11.5 ± 0.83 b | 41.15 ± 0.63 j | 14.2 ± 0.76 b |
| 80 µM | 0 | 0 a | 27.20 ± 0.42 k | 0 a | 35.1 ± 0.75 i | 0 a | 44.20 ± 0.97 kl |
| 80 µM | 150 µM Mn | 17.40 ± 1.10 def | 15.10 ± 0.80 f | 24.50± 0.62 f | 19.3 ± 0.82 ef | 26.15 ± 0.61 e | 29.6 ± 0.86 fg |
| 80 µM | 150 µM Cr | 23.37 ± 1.06 gh | 7.1 ± 0.69 b | 33.67 ± 0.70 j | 13.2 ± 0.80 dc | 37.12 ± 0.67 i | 15.9 ± 0.77 bc |
| 150 µM | 0 | 0 a | 31.00 ± 0.82 l | 0 a | 37.2 ± 0.98 j | 0 a | 7.70 ± 0.79 lm |
| 150 µM | 150 µM Mn | 15.32 ± 1.06 cd | 18.12 ± 0.70 g | 20.20 ± 0.73 d | 24.6 ± 0.90 cb | 23.10 ± 0.50 d | 33.6 ± 0.75 ghi |
| 150 µM | 150 µM Cr | 21.65 ± 0.85 g | 9.1 ± 0.69 c | 31.45 ± 0.96 i | 15.5 ± 0.90 c | 33.15 ± 0.80 h | 19.8 ± 1.06 cd |
| 250 µM | 0 | 0 a | 33.15 ± 0.81 m | 0 a | 42.1 ± 0.71 k | 0 a | 51.25 ± 8.12 mn |
| 250 µM | 150 µM Mn | 14.40 ± 1.11 c | 20.10 ± 0.74 h | 17.27 ± 0.70 c | 36.2 ± 0.94 ij | 21.25 ± 0.99 c | 36.5 ± 0.82 hij |
| 250 µM | 150 µM Cr | 18.27 ± 1.10 ef | 13.0 ± 0.78 e | 26.55 ± 0.65 g | 18.2 ± 0.94 de | 28.12 ± 0.72 f | 26.6 ± 0.82 ef |
| 400 µM | 0 | 0 a | 36.05 ± 0.87 n | 0 a | 45.3 ± 1.03 l | 0 a | 55.30 ± 1.03 n |
| 400 µM | 150 µM Mn | 12.15 ± 0.62 b | 23.10 ± 0.79 i | 15.40 ± 0.72 b | 29.2 ± 0.96 h | 18.02 ± 0.75 b | 38.2 ± 0.88 ij |
| 400 µM | 150 µM Cr | 16.30 ± 1.08 cde | 16.1 ± 0.61 f | 22.30 ± 0.71 e | 21.2 ± 0.80 f | 24.22 ± 0.56 d | 31.2 ± 0.96 fgh |
| Treatment | Chl-a (mg g−1 F.w.) | Chl-b (mg g−1 F.w.) | Chl a + b (mg g−1 F.w.) | Caratenoids (mg g−1 F.w.) |
|---|---|---|---|---|
| Control | 13.20 ± 0.15 ghi | 11.87 ± 0.66 gh | 22.32 ±4.87 efgh | 3.36 ± 0.52 defg |
| 150 µM Mn | 8.59 ± 0.42 ab | 6.96 ± 0.75 ab | 15.55 ± 1.17 ab | 2.02 ± 0.17 ab |
| 150 µM Cr | 7.77 ± 0.23 a | 6.09 ± 0.56 a | 13.86 ± 0.47 a | 1.46 ± 0.54 a |
| 50 µM SNP | 13.62 ± 0.28 hij | 13.43 ± 0.69 hij | 27.06 ± 0.97 ij | 3.57 ± 0.14 efgh |
| 50 µM SNP +150 µM Mn | 10.67 ± 0.40 de | 9.16 ± 0.74 cde | 19.83 ± 0.98 cdef | 2.63 ± 0.16 bcd |
| 50 µM SNP +150 µM Cr | 8.74 ± 0.38 ab | 7.96 ± 0.76 abc | 16.71 ± 1.12 abc | 2.41 ± 0.17 bc |
| 80 µM SNP | 13.91 ± 0.38 ij | 13.90 ± 0.52 ijk | 27.82 ± 0.30 ijk | 3.67 ± 0.10 fgh |
| 80 µM SNP +150 µM Mn | 11.64 ± 0.23 ef | 10.29 ± 0.84 defg | 21.93 ± 0.70 efgh | 2.86 ± 0.21 cde |
| 80 µM SNP +150 µM Cr | 9.50 ± 0.36 bc | 8.21 ± 0.73 bc | 17.71 ± 0.94 bcd | 2.45 ± 0.14 bc |
| 150 µM SNP | 14.17 ± 0.53 ijk | 15.26 ± 1.05 jkl | 29.43 ± 1.01 jk | 4.14 ± 0.55 h |
| 150 µM SNP +150 µM Mn | 12.73 ± 0.24 gh | 11.37 ± 0.95 fg | 24.11 ± 1.19 ghi | 3.01 ± 0.16 cdef |
| 150 µM SNP +150 µM Cr | 10.05 ± 0.54 cd | 8.62 ± 0.44 bcd | 18.67 ± 0.12 bcde | 2.48 ± 0.06 bc |
| 250 µM SNP | 14.46 ± 0.19 jk | 15.52 ± 0.68 kl | 29.91 ± 0.89 jk | 4.01 ± 0.13 gh |
| 250 µM SNP +150 µM Mn | 13.35 ± 0.32 hi | 12.05 ± 0.62 ghi | 25.40 ± 0.94 hi | 3.12 ± 0.09 cdef |
| 250 µM SNP +150 µM Cr | 11.28 ± 0.49 e | 9.67 ± 0.85 cdef | 20.95 ± 0.37 defg | 2.79 ± 0.51 cd |
| 400 µM SNP | 14.94 ± 0.70 k | 16.11 ± 0.66 l | 31.06 ± 1.31 k | 4.16 ± 0.15 h |
| 400 µM SNP + 150 µM Mn | 13.56 ± 0.28 hij | 13.31 ± 0.88 hi | 26.88 ± 1.15 ij | 3.56 ± 0.19 efgh |
| 400 µM SNP + 150 µM Cr | 12.33 ± 0.40 fg | 10.61 ± 0.72 efg | 22.95 ±1.11 fgh | 2.85 ± 0.16 cde |
| SNP Levels (µM) | (Mn) and(Cr) Levels | Dryshoot Weight | Dry Root Weight | Shoot Lenght |
|---|---|---|---|---|
| 0 | 150 µM Mn | 33% ↓ | 32%↓ | 18% ↓ |
| 0 | 150 µM Cr | 42% ↓ | 36%↓ | 22% ↓ |
| 50 | 0 | 10% ↑ | 12%↑ | 5% ↑ |
| 50 | 150 µM Mn | 22% ↓ | 18%↓ | 9% ↓ |
| 50 | 150 µM Cr | 31% ↓ | 29%↓ | 16% ↓ |
| 100 | 0 | 14% ↑ | 16%↑ | 6% ↑ |
| 100 | 150 µM Mn | 15% ↓ | 12%↓ | 5% ↓ |
| 100 | 150 µM Cr | 29% ↓ | 25%↓ | 14% ↓ |
| 150 | 0 | 19% ↑ | 8% ↑ | 21% ↑ |
| 150 | 150 µM Mn | 4% ↓ | 1% ↓ | 4% ↓ |
| 150 | 150 µM Cr | 26% ↓ | 11% ↓ | 22% ↓ |
| 250 | 0 | 24% ↑ | 16% ↑ | 25% ↑ |
| 250 | 150 µM Mn | 1% ↑ | 4%↑ | 2% ↑ |
| 250 | 150 µM Cr | 18% ↓ | 15% ↓ | 7% ↓ |
| 400 | 0 | 31% ↑ | 5% ↑ | 35% ↑ |
| 400 | 150 µM Mn | 7% ↑ | 3% ↑ | 8% ↑ |
| 400 | 150 µM Cr | 11% ↓ | 4% ↓ | 8% ↓ |
| Treatments | Translocation Factor (TF) (Leaves) | Tolerance Index (TI) (Shoot) | Tolerance Index (TI) (Root) | Bioaccumulation Factor (Leaves) (BAF) |
|---|---|---|---|---|
| Control | 0.00 ± 0.00 a | 1.00 ± 0.00 ghi | 1.00 ± 0.00 hij | 0.00 ± 0.00 a |
| 150 µM Mn | 0.69 ± 0.01 bc | 0.66 ± 0.04 ab | 0.67 ± 0.03 ab | 0.18 ± 0.001 i |
| 150 µM Cr | 0.69 ± 0.04 c | 0.57 ± 0.04 a | 0.63 ± 0.08 a | 0.21 ± 0.006 j |
| 50 µM SNP | 0.00 ± 0.00 a | 1.10 ± 0.05 ijk | 1.12 ± 0.10 klm | 0.00 ± 0.00 a |
| 50 µM SNP +150 µM Mn | 0.65 ± 0.02 bc | 0.77 ± 0.05 bcde | 0.81 ± 0.02 cdef | 0.12 ± 0.005 f |
| 50 µM SNP +150 µM Cr | 0.67 ± 0.02 bc | 0.68 ± 0.02 ab | 0.70 ± 0.01 abc | 0.16 ± 0.007 h |
| 80 µM SNP | 0.00 ± 0.00 a | 1.14 ± 0.05 jkl | 1.16 ± 0.05 lmn | 0.00 ± 0.00 a |
| 80 µM SNP +150 µM Mn | 0.66 ± 0.03 bc | 0.84 ± 0.05 def | 0.87 ± 0.03 efg | 0.11 ± 0.004 de |
| 80 µM SNP +150 µM Cr | 0.67 ± 0.01 bc | 0.70 ± 0.05 bc | 0.74 ± 0.00 abcd | 0.15 ± 0.006 gh |
| 150 µM SNP | 0.00 ± 0.00 a | 1.19 ± 0.02 kl | 1.22 ± 0.09 mn | 0.00 ± 0.00 a |
| 150 µM SNP +150 µM Mn | 0.63 ± 0.03 bc | 0.95 ± 0.06 fgh | 0.95 ± 0.01 ghi | 0.10 ± 0.006 cd |
| 150 µM SNP+150 µM Cr | 0.67 ± 0.01 bc | 0.73 ± 0.04 bcd | 0.77 ± 0.00 bcde | 0.14 ± 0.005 g |
| 250 µM SNP | 0.00 ± 0.00 a | 1.24 ± 0.03 lm | 1.25 ± 0.03 no | 0.00 ± 0.00 a |
| 250 µM SNP +150 µM Mn | 0.62 ± 0.01 bc | 1.01 ± 0.05 hi | 1.04 ± 0.02 ijk | 0.09 ± 0.007 c |
| 250 µM SNP +150 µM Cr | 0.63 ± 0.01 bc | 0.81 ± 0.04 cde | 0.84 ± 0.02 defg | 0.12 ± 0.007 ef |
| 400 µM SNP | 0.00 ± 0.00 a | 1.31 ± 0.03 m | 1.35 ± 0.02 o | 0.00 ± 0.00 a |
| 400 µM SNP +150 µM Mn | 0.60 ± 0.02 b | 1.07 ± 0.04 ij | 1.08 ± 0.02 jkl | 0.08 ± 0.004 b |
| 400 µM SNP +150 µM Cr | 0.63 ± 0.01 bc | 0.89 ± 0.04 efg | 0.91 ± 0.04 fgh | 0.10 ± 0.007 cde |
| Experiment Treatments | Concentrations |
|---|---|
| Control | 0 |
| Mn | 150 µM Mn |
| Cr | 150 µM Cr |
| SNP | 50 µM SNP |
| SNP +Mn | 50 µM SNP + 150 µM Mn |
| SNP +Cr | 50 µM SNP + 150 µM Cr |
| SNP | 80 µM SNP |
| SNP +Mn | 80 µM SNP + 150 µM Mn |
| SNP +Cr | 80 µM SNP + 150 µM Cr |
| SNP | 150 µM SNP |
| SNP +Mn | 150 µM SNP + 150 µM Mn |
| SNP +Cr | 150 µM SNP + 150 µM Cr |
| SNP | 250 µM SNP |
| SNP +Mn | 250 µM SNP + 150 µM Mn |
| SNP +Cr | 250 µM SNP + 150 µM Cr |
| SNP | 400 µM SNP |
| SNP +Mn | 400 µM SNP + 150 µM Mn |
| SNP +Cr | 400 µM SNP + 150 µM Cr |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emamverdian, A.; Ding, Y.; Barker, J.; Liu, G.; Li, Y.; Mokhberdoran, F. Sodium Nitroprusside Improves Bamboo Resistance under Mn and Cr Toxicity with Stimulation of Antioxidants Activity, Relative Water Content, and Metal Translocation and Accumulation. Int. J. Mol. Sci. 2023, 24, 1942. https://doi.org/10.3390/ijms24031942
Emamverdian A, Ding Y, Barker J, Liu G, Li Y, Mokhberdoran F. Sodium Nitroprusside Improves Bamboo Resistance under Mn and Cr Toxicity with Stimulation of Antioxidants Activity, Relative Water Content, and Metal Translocation and Accumulation. International Journal of Molecular Sciences. 2023; 24(3):1942. https://doi.org/10.3390/ijms24031942
Chicago/Turabian StyleEmamverdian, Abolghassem, Yulong Ding, James Barker, Guohua Liu, Yang Li, and Farzad Mokhberdoran. 2023. "Sodium Nitroprusside Improves Bamboo Resistance under Mn and Cr Toxicity with Stimulation of Antioxidants Activity, Relative Water Content, and Metal Translocation and Accumulation" International Journal of Molecular Sciences 24, no. 3: 1942. https://doi.org/10.3390/ijms24031942
APA StyleEmamverdian, A., Ding, Y., Barker, J., Liu, G., Li, Y., & Mokhberdoran, F. (2023). Sodium Nitroprusside Improves Bamboo Resistance under Mn and Cr Toxicity with Stimulation of Antioxidants Activity, Relative Water Content, and Metal Translocation and Accumulation. International Journal of Molecular Sciences, 24(3), 1942. https://doi.org/10.3390/ijms24031942