
The Remodeling of Dermal Collagen Fibrous Structures in Mice under Zero Gravity: The Role of Mast Cells

 , ,
, ,
Abstract
1. Introduction
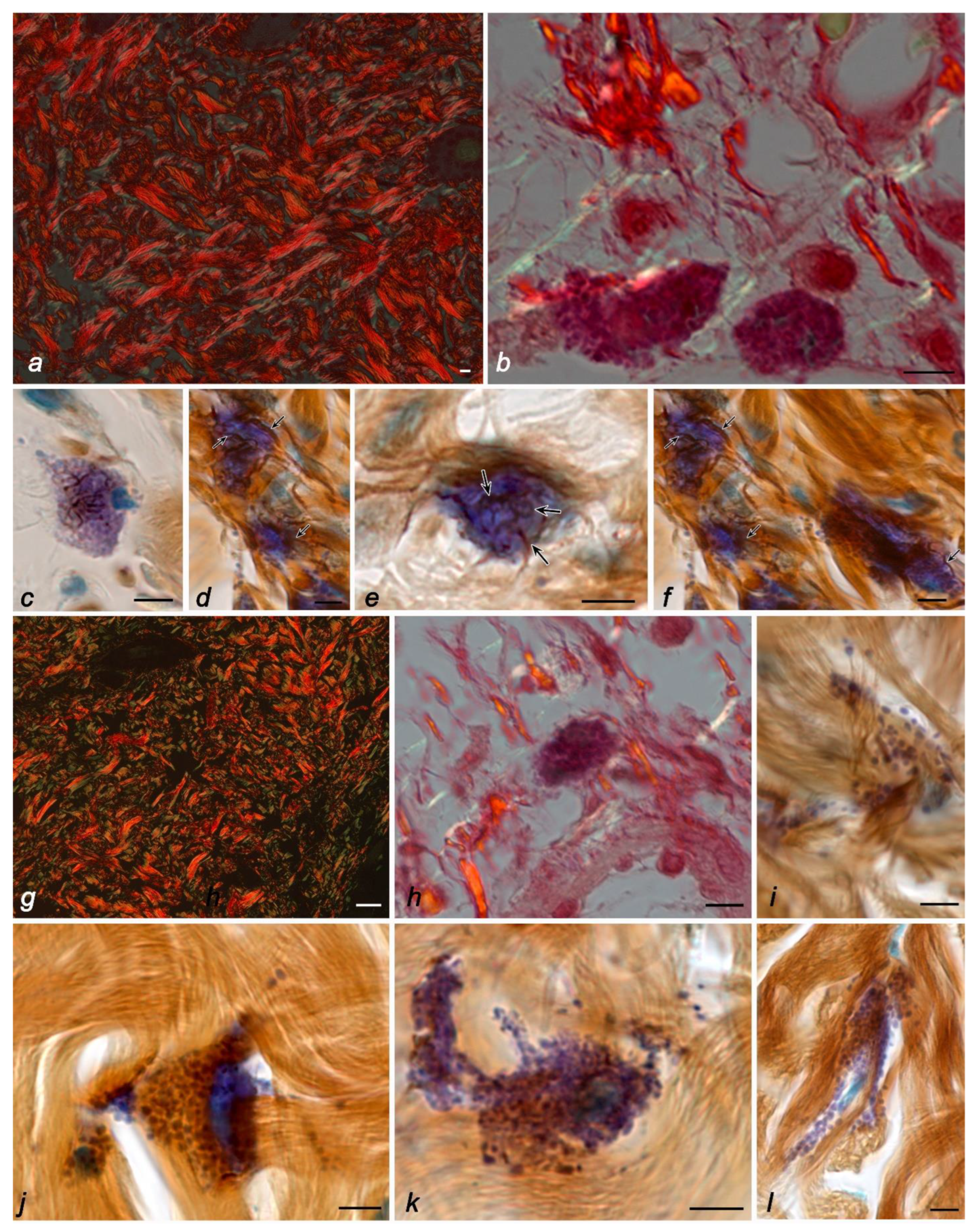
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Histoprocessing
4.3. Tissue Probe Staining
4.4. Image Acquisition
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zhivodernikov, I.V.; Ratushnyy, A.Y.; Matveeva, D.K.; Buravkova, L.B. Extracellular Matrix Proteins and Transcription of Matrix-Associated Genes in Mesenchymal Stromal Cells during Modeling of the Effects of Microgravity. Bull. Exp. Biol. Med. 2020, 170, 230–232. [Google Scholar] [CrossRef]
- Buravkova, L.; Larina, I.; Andreeva, E.; Grigoriev, A. Microgravity Effects on the Matrisome. Cells 2021, 10, 2226. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, E.; Matveeva, D.; Zhidkova, O.; Zhivodernikov, I.; Kotov, O.; Buravkova, L. Real and Simulated Microgravity: Focus on Mammalian Extracellular Matrix. Life 2022, 12, 1343. [Google Scholar] [CrossRef] [PubMed]
- Atiakshin, D.A.; Alexeeva, N.T.; Klochkova, S.V.; Nikityuk, D.B. Extracellular matrix collagen fiber structures of the gastrointestinal connective tissues in mice after a 30 day orbital flight. Vopr. Pitan. 2019, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Atiakshin, D.A.; Bykov, E.G.; Il’in, E.A.; Pashkov, A.N. Tissue-specific reaction of the mucous coat of herbals’ small gut under the influence of spaceflight factors on board biosat “Foton M3”. Aviakosm. Ekolog. Med. 2011, 45, 25–30. [Google Scholar] [PubMed]
- Atiakshin, D.A.; Il’in, E.A.; Pashkov, A.N. Morphofunctional state of hepatocytes nuclear apparatus in Mongolian herbils after the flight on space apparatus Foton-M3. Aviakosm. Ekolog. Med. 2010, 44, 29–34. [Google Scholar]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Bhutda, S.; Surve, M.V.; Anil, A.; Kamath, K.; Singh, N.; Modi, D.; Banerjee, A. Histochemical Staining of Collagen and Identification of Its Subtypes by Picrosirius Red Dye in Mouse Reproductive Tissues. Bio-Protocol 2017, 7, e2592. [Google Scholar] [CrossRef]
- Atiakshin, D.; Buchwalow, I.; Tiemann, M. Mast cells and collagen fibrillogenesis. Histochem. Cell Biol. 2020, 154, 21–40. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Olivera, A.; Beaven, M.A.; Metcalfe, D.D. Mast cells signal their importance in health and disease. J. Allergy Clin. Immunol. 2018, 142, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Atiakshin, D.; Buchwalow, I.; Horny, P.; Tiemann, M. Protease profile of normal and neoplastic mast cells in the human bone marrow with special emphasis on systemic mastocytosis. Histochem. Cell Biol. 2021, 155, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A Review of the Contribution of Mast Cells in Wound Healing: Involved Molecular and Cellular Mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Gur’eva, T.S.; Dadasheva, O.A.; Mednikova, E.I.; Dadasheva, M.T.; Sychev, V.N. Histogeny of the visceral organs of embryonic Japanese quails developed in the micro-g environment. Aviakosm. Ekolog. Med. 2009, 43, 8–13. [Google Scholar] [PubMed]
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The Collagen Suprafamily: From Biosynthesis to Advanced Biomaterial Development. Adv. Mater. 2019, 31, e1801651. [Google Scholar] [CrossRef] [PubMed]
- Changoor, A.; Tran-Khanh, N.; Methot, S.; Garon, M.; Hurtig, M.B.; Shive, M.S.; Buschmann, M.D. A polarized light microscopy method for accurate and reliable grading of collagen organization in cartilage repair. Osteoarthr. Cartil. 2011, 19, 126–135. [Google Scholar] [CrossRef]
- Gadd, V.L. Combining immunodetection with histochemical techniques: The effect of heat-induced antigen retrieval on picro-Sirius red staining. J. Histochem. Cytochem. 2014, 62, 902–906. [Google Scholar] [CrossRef]
- Surve, M.V.; Anil, A.; Kamath, K.G.; Bhutda, S.; Sthanam, L.K.; Pradhan, A.; Srivastava, R.; Basu, B.; Dutta, S.; Sen, S.; et al. Membrane Vesicles of Group B Streptococcus Disrupt Feto-Maternal Barrier Leading to Preterm Birth. PLoS Pathog. 2016, 12, e1005816. [Google Scholar] [CrossRef]
- Segnani, C.; Ippolito, C.; Antonioli, L.; Pellegrini, C.; Blandizzi, C.; Dolfi, A.; Bernardini, N. Histochemical Detection of Collagen Fibers by Sirius Red/Fast Green Is More Sensitive than van Gieson or Sirius Red Alone in Normal and Inflamed Rat Colon. PLoS ONE 2015, 10, e0144630. [Google Scholar] [CrossRef]
- Chandran, A.; Bhandary, R.; Shenoy, N.; Shetty, U.A. Analysis of collagen fibers in human gingival tissues using picrosirius red stain under polarized microscope. J. Indian Soc. Periodontol. 2021, 25, 106–111. [Google Scholar] [CrossRef]
- Rieppo, L.; Janssen, L.; Rahunen, K.; Lehenkari, P.; Finnila, M.A.J.; Saarakkala, S. Histochemical quantification of collagen content in articular cartilage. PLoS ONE 2019, 14, e0224839. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Xu, Y.J.; Zhang, Z.X.; Hu, J. Expression of pulmonary artery connective tissue growth factor and pulmonary artery remodeling in smokers and patients with chronic obstructive pulmonary disease. Zhonghua Nei Ke Za Zhi 2007, 46, 298–301. [Google Scholar] [PubMed]
- Rittie, L. Method for Picrosirius Red-Polarization Detection of Collagen Fibers in Tissue Sections. Methods Mol. Biol. 2017, 1627, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Greiner, C.; Grainger, S.; Farrow, S.; Davis, A.; Su, J.L.; Saybolt, M.D.; Wilensky, R.; Madden, S.; Sum, S.T. Robust quantitative assessment of collagen fibers with picrosirius red stain and linearly polarized light as demonstrated on atherosclerotic plaque samples. PLoS ONE 2021, 16, e0248068. [Google Scholar] [CrossRef]
- Bora, V.; Patel, D.; Johar, K.; Goyal, R.K.; Patel, B.M. Systemic study of selected histone deacetylase inhibitors in cardiac complications associated with cancer cachexia. Can. J. Physiol. Pharmacol. 2022, 100, 240–251. [Google Scholar] [CrossRef]
- Liu, J.; Xu, M.Y.; Wu, J.; Zhang, H.; Yang, L.; Lun, D.X.; Hu, Y.C.; Liu, B. Picrosirius-Polarization Method for Collagen Fiber Detection in Tendons: A Mini-Review. Orthop. Surg. 2021, 13, 701–707. [Google Scholar] [CrossRef]
- Lattouf, R.; Younes, R.; Lutomski, D.; Naaman, N.; Godeau, G.; Senni, K.; Changotade, S. Picrosirius red staining: A useful tool to appraise collagen networks in normal and pathological tissues. J. Histochem. Cytochem. 2014, 62, 751–758. [Google Scholar] [CrossRef]
- Wang, G.; Fan, W.T.; Zhang, Z.; Huang, S.G. Expression of matrix metalloproteinase-8 and matrix metalloproteinase-13 in mast cells of human periapical lesions. Int. J. Clin. Exp. Pathol. 2018, 11, 2530–2536. [Google Scholar]
- Lojda, Z.; Gossrau, R.; Schiebler, T. Enzyme Histochemistry: A Laboratory Manual; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1979. [Google Scholar]
- Adachi, E.; Hayashi, T.; Hashimoto, P.H. A comparison of the immunofluorescent localization of collagen types I, III, and V with the distribution of reticular fibers on the same liver sections of the snow monkey (Macaca fuscata). Cell Tissue Res. 1991, 264, 1–8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Groups of the Experiment | Number of MCs (Per 1 mm2) | The Frequency of MCs’ Adherence to Each Other (in % of the Total Number) |
|---|---|---|
| Vivarium control | 34.4 ± 3.1 | 4.59 ± 0.29 ∞ |
| Basal control | 39.1 ± 4.6 | 5.24 ± 0.41 ∞ * |
| Ground control | 32.6 ± 3.2 | 3.75 ± 0.27 + |
| Space flight | 36.6 ± 3.2 | 3.51 ± 0.18 + ⸋ |
| Groups of the Experiment | MCs without Signs of Secretion | MCs with Signs of Secretion | Nuclear-Free Fragments of MCs |
|---|---|---|---|
| Vivarium control | 26.2 ± 2.7 ∞ | 45.7 ± 3.7 ∞ + | 28.1 ± 1.9 |
| Basal control | 21.8 ± 2.4 ∞ | 54.8 ± 2.1 ⸋ ∞ | 23.4 ± 2.5 |
| Ground control | 25.3 ± 2.1 ∞ | 47.1 ± 4.1 ∞ | 27.6 ± 2.1 |
| Space flight | 11.5 ± 0.8 * + ⸋ | 61.5 ± 3.3 * + ⸋ | 27.0 ± 1.8 |
| Dyes | Catalogue Number | Provider | Dilution | Manufacturer |
|---|---|---|---|---|
| Toluidine blue | 07-002 | Biovitrum | Ready-to-use | ErgoProduction LLC, Saint-Petersburg, Russia |
| Giemsa solution | 20-043/L | Biovitrum | Ready-to-use | ErgoProduction LLC, Russia |
| Picrosirius red | ab150681—Picro Sirius Red Stain Kit (Connective Tissue Stain) | Abcam | Ready-to-use | Abcam, Cambridge, UK |
| Silver impregnation | 21-026 | Biovitrum | Ready-to-use | ErgoProduction LLC, Russia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shishkina, V.; Kostin, A.; Volodkin, A.; Samoilova, V.; Buchwalow, I.; Tiemann, M.; Atiakshin, D. The Remodeling of Dermal Collagen Fibrous Structures in Mice under Zero Gravity: The Role of Mast Cells. Int. J. Mol. Sci. 2023, 24, 1939. https://doi.org/10.3390/ijms24031939
Shishkina V, Kostin A, Volodkin A, Samoilova V, Buchwalow I, Tiemann M, Atiakshin D. The Remodeling of Dermal Collagen Fibrous Structures in Mice under Zero Gravity: The Role of Mast Cells. International Journal of Molecular Sciences. 2023; 24(3):1939. https://doi.org/10.3390/ijms24031939
Chicago/Turabian StyleShishkina, Viktoriya, Andrey Kostin, Artem Volodkin, Vera Samoilova, Igor Buchwalow, Markus Tiemann, and Dmitri Atiakshin. 2023. "The Remodeling of Dermal Collagen Fibrous Structures in Mice under Zero Gravity: The Role of Mast Cells" International Journal of Molecular Sciences 24, no. 3: 1939. https://doi.org/10.3390/ijms24031939
APA StyleShishkina, V., Kostin, A., Volodkin, A., Samoilova, V., Buchwalow, I., Tiemann, M., & Atiakshin, D. (2023). The Remodeling of Dermal Collagen Fibrous Structures in Mice under Zero Gravity: The Role of Mast Cells. International Journal of Molecular Sciences, 24(3), 1939. https://doi.org/10.3390/ijms24031939