
Bacterial Motility and Its Role in Skin and Wound Infections

Abstract
1. Introduction
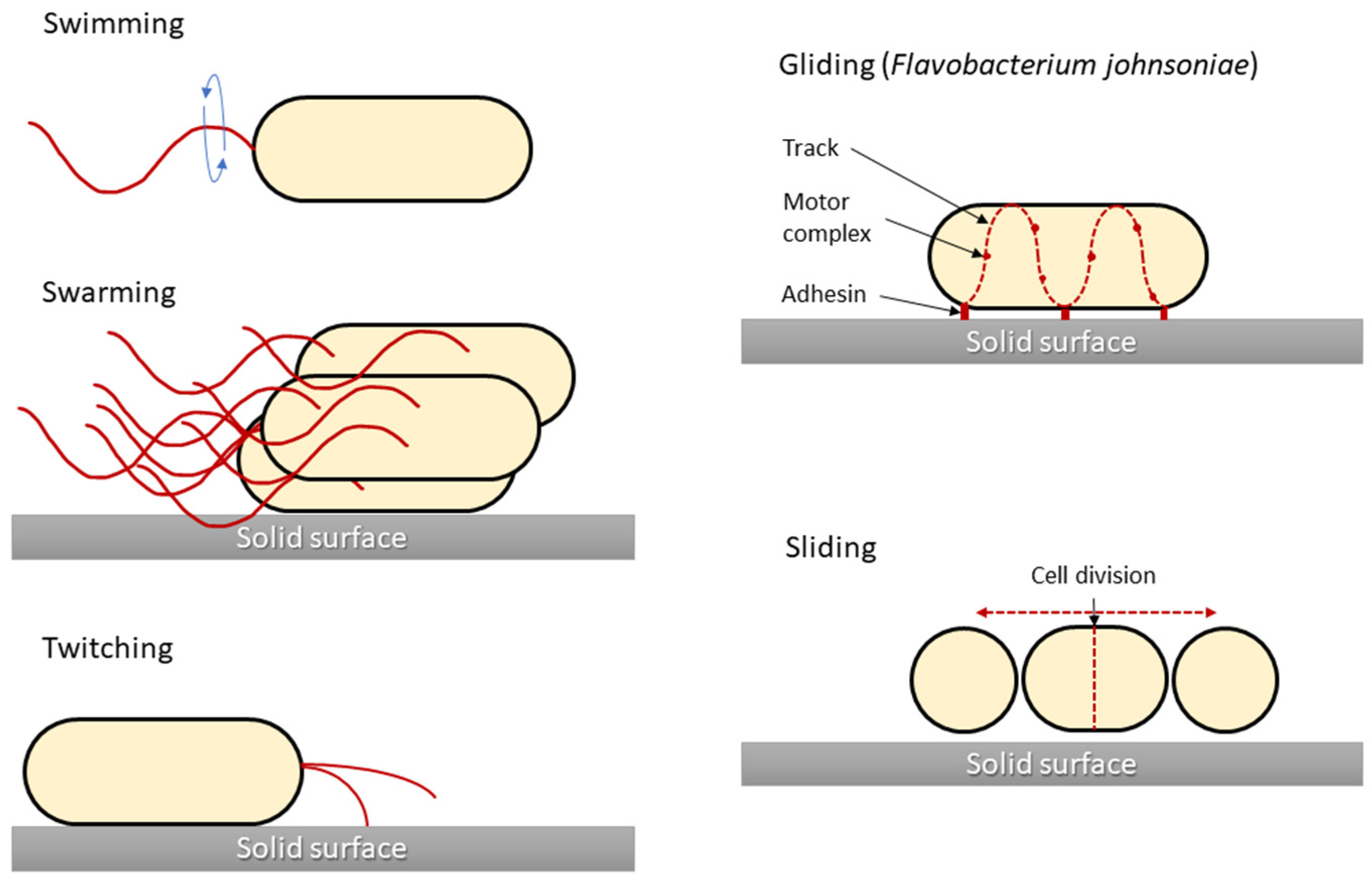
2. Types of Bacterial Movement
2.1. Swimming Motility
2.2. Swarming Motility
2.3. Twitching Motility
2.4. Gliding Motility
2.5. Sliding Motility
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schommer, N.N.; Gallo, R.L. Structure and Function of the Human Skin Microbiome. Trends Microbiol. 2013, 21, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- DiNubile, M.J.; Lipsky, B.A. Complicated Infections of Skin and Skin Structures: When the Infection Is More than Skip Deep. J. Antimicrob. Chemother. 2004, 53, ii37–ii50. [Google Scholar] [CrossRef] [PubMed]
- Kaye, K.S.; Petty, L.A.; Shorr, A.F.; Zilberberg, M.D. Current Epidemiology, Etiology, and Burden of Acute Skin Infections in the United States. Clin. Infect. Dis. 2019, 68, S193–S199. [Google Scholar] [CrossRef] [PubMed]
- Tognetti, L.; Martinelli, C.; Berti, S.; Hercogova, J.; Lotti, T.; Leoncini, F.; Moretti, S. Bacterial Skin and Soft Tissue Infections: Review of the Epidemiology, Microbiology, Aetiopathogenesis and Treatment: A Collaboration between Dermatologists and Infectivologists. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Plewig, G.; French, L.; Ruzicka, T.; Kaufmann, R.; Hertl, M. Braun-Falco’s Dermatology, 2020th ed.; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Cates, J.E.; Mitrani-Gold, F.S.; Li, G.; Mundy, L.M. Systematic Review and Meta-Analysis to Estimate Antibacterial Treatment Effect in Acute Bacterial Skin and Skin Structure Infection. Antimicrob. Agents Chemother. 2015, 59, 4510–4520. [Google Scholar] [CrossRef]
- Olsson, M.; Järbrink, K.; Divakar, U.; Bajpai, R.; Upton, Z.; Schmidtchen, A.; Car, J. The Humanistic and Economic Burden of Chronic Wounds: A Systematic Review: The Humanistic and Economic Burden of Chronic Wounds. Wound Repair Regen. 2019, 27, 114–125. [Google Scholar] [CrossRef]
- Bessa, L.J.; Fazii, P.; di Giulio, M.; Cellini, L. Bacterial Isolates from Infected Wounds and Their Antibiotic Susceptibility Pattern: Some Remarks about Wound Infection. Int. Wound J. 2015, 12, 47–52. [Google Scholar] [CrossRef]
- Puca, V.; Marulli, R.Z.; Grande, R.; Vitale, I.; Niro, A.; Molinaro, G.; Prezioso, S.; Muraro, R.; di Giovanni, P. Microbial Species Isolated from Infected Wounds and Antimicrobial Resistance Analysis: Data Emerging from a Three-Years Retrospective Study. Antibiotics 2021, 10, 1162. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.D.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the Chronic Wound Microbiota of 2,963 Patients by 16S RDNA Pyrosequencing. Wound Repair Regen. 2016, 24, 163–174. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Guoqi, W.; Zhirui, L.; Song, W.; Tongtong, L.; Lihai, Z.; Licheng, Z.; Peifu, T. Negative Pressure Wound Therapy Reduces the Motility of Pseudomonas Aeruginosa and Enhances Wound Healing in a Rabbit Ear Biofilm Infection Model. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2018, 111, 1557–1570. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, M.; Behrouz, B.; Irajian, G.; Mahdavi, M.; Korpi, F.; Motamedifar, M. Passive Immunization against Pseudomonas Aeruginosa Recombinant PilA in a Murine Burn Wound Model. Microb. Pathog. 2016, 101, 83–88. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, G.; Morohoshi, T.; Ikeda, T.; Ohta, Y.; Sagara, H.; Huang, L.; Nagase, T.; Sugama, J.; Sanada, H. Contribution of Quorum Sensing to the Virulence of Pseudomonas Aeruginosa in Pressure Ulcer Infection in Rats. Wound Repair Regen. 2011, 19, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Clinmicrorevs, M.; Bowler, P.; Armstrong, D.G. Wound Microbiology and Associated Approaches to Wound. Clin. Microbiol. Rev. 2016, 14, 244–269. [Google Scholar] [CrossRef]
- Jawień, A.; Bartoszewicz, M.; Przondo-Mordarska, A.; Szewczyk, M.T.; Kaszuba, A.; Urbanek, T.; Staszkiewicz, W.; Sopata, M.; Kucharzewski, M.; Korzon-Burakowska, A.; et al. Wytyczne Postępowania Miejscowego i Ogólnego w Ranach Objętych Procesem Infekcji. Leczenie Ran 2012, 9, 59–75. [Google Scholar]
- White, R.; Cutting, K. Critical Colonisation of Chronic Wounds: Microbial Mechanisms. Wounds 2008, 4, 70–78. [Google Scholar]
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef]
- Huang, X.; Liang, P.; Jiang, B.; Zhang, P.; Yu, W.; Duan, M.; Guo, L.; Cui, X.; Huang, M.; Huang, X. Hyperbaric Oxygen Potentiates Diabetic Wound Healing by Promoting Fibroblast Cell Proliferation and Endothelial Cell Angiogenesis. Life Sci. 2020, 259, 118246. [Google Scholar] [CrossRef]
- Agarwal, P.; Kukrele, R.; Sharma, D. Vacuum Assisted Closure (VAC)/Negative Pressure Wound Therapy (NPWT) for Difficult Wounds: A Review. J. Clin. Orthop. Trauma 2019, 10, 845–848. [Google Scholar] [CrossRef]
- Gupta, S.; Gabriel, A.; Lantis, J.; Téot, L. Clinical Recommendations and Practical Guide for Negative Pressure Wound Therapy with Instillation. Int. Wound J. 2016, 13, 159–174. [Google Scholar] [CrossRef]
- Kloth, L.C. Discussion: Advanced Technologies to Improve Wound Healing: Electrical Stimulation, Vibration Therapy, and Ultrasound-What Is the Evidence? Plast. Reconstr. Surg. 2016, 138, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Balakatounis, K.C.; Angoules, A.G. Low-Intensity Electrical Stimulation in Wound Healing: Review of the Efficacy of Externally Applied Currents Resembling the Current of Injury. Eplasty 2008, 8, e28. [Google Scholar]
- Ashrafi, M.; Alonso-Rasgado, T.; Baguneid, M.; Bayat, A. The Efficacy of Electrical Stimulation in Lower Extremity Cutaneous Wound Healing: A Systematic Review. Exp. Dermatol. 2017, 26, 171–178. [Google Scholar] [CrossRef]
- Singh, M.R.; Saraf, S.; Vyas, A.; Jain, V.; Singh, D. Innovative Approaches in Wound Healing: Trajectory and Advances. Artif. Cells Nanomed. Biotechnol. 2013, 41, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Bekara, F.; Vitse, J.; Fluieraru, S.; Masson, R.; de Runz, A.; Georgescu, V.; Bressy, G.; Labbé, J.L.; Chaput, B.; Herlin, C. New Techniques for Wound Management: A Systematic Review of Their Role in the Management of Chronic Wounds. Arch. Plast. Surg. 2018, 45, 102–110. [Google Scholar] [CrossRef]
- Henrichsen, J. Bacterial Surface Translocation: A Survey and a Classification. Bacteriol. Rev. 1972, 36, 478–503. [Google Scholar] [CrossRef]
- Shrout, J.D. A Fantastic Voyage for Sliding Bacteria. Trends Microbiol. 2015, 23, 244–246. [Google Scholar] [CrossRef]
- Wadhwa, N.; Berg, H.C. Bacterial Motility: Machinery and Mechanisms. Nat. Rev. Microbiol. 2022, 20, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Minnullina, L.; Kostennikova, Z.; Evtugin, V.; Akosah, Y.; Sharipova, M.; Mardanova, A. Diversity in the Swimming Motility and Flagellar Regulon Structure of Uropathogenic Morganella Morganii Strains. Int. Microbiol. 2022, 25, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Corral, J.; Sebastià, P.; Coll, N.S.; Barbé, J.; Aranda, J.; Valls, M. Twitching and Swimming Motility Play a Role in Ralstonia Solanacearum Pathogenicity. Msphere 2020, 5, e00740-19. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Robinson, R.C.; Uyeda, T.Q.P.; Fukumori, Y.; Fukushima, S.I.; Haruta, S.; Homma, M.; Inaba, K.; Ito, M.; Kaito, C.; et al. Tree of Motility—A Proposed History of Motility Systems in the Tree of Life. Genes Cells 2020, 25, 6–21. [Google Scholar] [CrossRef]
- Colin, R.; Ni, B.; Laganenka, L.; Sourjik, V. Multiple Functions of Flagellar Motility and Chemotaxis in Bacterial Physiology. FEMS Microbiol. Rev. 2021, 45, fuab038. [Google Scholar] [CrossRef]
- Li, Y.; Xia, H.; Bai, F.; Song, X.; Zhuang, L.; Xu, H.; Zhang, X.; Qiao, M. PA5001 Gene Involves in Swimming Motility and Biofilm Formation in Pseudomonas Aeruginosa. Microb. Pathog. 2020, 144, 103982. [Google Scholar] [CrossRef]
- Terashima, H.; Kawamoto, A.; Morimoto, Y.V.; Imada, K.; Minamino, T. Structural Differences in the Bacterial Flagellar Motor among Bacterial Species. Biophys. Physicobiol. 2017, 14, 191–198. [Google Scholar] [CrossRef]
- Bardy, S.L.; Ng, S.Y.M.; Jarrell, K.F. Prokaryotic Motility Structures. Microbiology 2003, 149, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Liu, S.; Hancock, R.E.W. Surfing Motility: A Conserved yet Diverse Adaptation among Motile Bacteria. J. Bacteriol. 2018, 191, 5569–5576. [Google Scholar] [CrossRef]
- Schauer, O.; Mostaghaci, B.; Colin, R.; Hürtgen, D.; Kraus, D.; Sitti, M.; Sourjik, V. Motility and Chemotaxis of Bacteria-Driven Microswimmers Fabricated Using Antigen 43-Mediated Biotin Display. Sci. Rep. 2018, 8, 9801. [Google Scholar] [CrossRef]
- Schwanbeck, J.; Oehmig, I.; Groß, U.; Zautner, A.E.; Bohne, W. Clostridioides Difficile Single Cell Swimming Strategy: A Novel Motility Pattern Regulated by Viscoelastic Properties of the Environment. Front. Microbiol. 2021, 12, 715220. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Vu, A.; Lee, K.; Dahlquist, F.W. CheA-Receptor Interaction Sites in Bacterial Chemotaxis. J. Mol. Biol. 2012, 422, 282–290. [Google Scholar] [CrossRef]
- Khong, N.Z.J.; Zeng, Y.; Lai, S.K.; Koh, C.G.; Liang, Z.X.; Chiam, K.H.; Li, H.Y. Dynamic Swimming Pattern of Pseudomonas Aeruginosa near a Vertical Wall during Initial Attachment Stages of Biofilm Formation. Sci. Rep. 2021, 11, 1952. [Google Scholar] [CrossRef] [PubMed]
- Samad, T.; Billings, N.; Birjiniuk, A.; Crouzier, T.; Doyle, P.S.; Ribbeck, K. Swimming Bacteria Promote Dispersal of Non-Motile Staphylococcal Species. ISME J. 2017, 11, 1933–1937. [Google Scholar] [CrossRef] [PubMed]
- Madukoma, C.S.; Liang, P.; Dimkovikj, A.; Chen, J.; Lee, S.W.; Chen, D.Z.; Shrout, J.D. Single Cells Exhibit Differing Behavioral Phases during Early Stages of Pseudomonas Aeruginosa Swarming. J. Bacteriol. 2019, 201, e00184-19. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.D.; Hewitt, J.L.; Wolfe, L.G.; Kamatkar, N.G.; Chapman, S.M.; Diener, J.M.; Courtney, A.J.; Leevy, W.M.; Shrout, J.D. Imaging and Analysis of Pseudomonas Aeruginosa Swarming and Rhamnolipid Production. Appl. Environ. Microbiol. 2011, 77, 8310–8317. [Google Scholar] [CrossRef] [PubMed]
- Be’er, A.; Ariel, G. A Statistical Physics View of Swarming Bacteria. Mov. Ecol. 2019, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A Field Guide to Bacterial Swarming Motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef]
- Walker, K.E.; Moghaddame-Jafari, S.; Lockatell, C.V.; Johnson, D.; Belas, R. ZapA, the IgA-Degrading Metalloprotease of Proteus Mirabilis, Is a Virulence Factor Expressed Specifically in Swarmer Cells. Mol. Microbiol. 1999, 32, 825–836. [Google Scholar] [CrossRef]
- Yan, J.; Monaco, H.; Xavier, J.B. The Ultimate Guide to Bacterial Swarming: An Experimental Model to Study the Evolution of Cooperative Behavior. Annu. Rev. Microbiol. 2019, 73, 293–312. [Google Scholar] [CrossRef]
- Morgenstein, R.M.; Szostek, B.; Rather, P.N. Regulation of Gene Expression during Swarmer Cell Differentiation in Proteus Mirabilis. FEMS Microbiol. Rev. 2010, 34, 753–763. [Google Scholar] [CrossRef]
- Toguchi, A.; Siano, M.; Burkart, M.; Harshey, R.M. Genetics of Swarming Motility in Salmonella Enterica Serovar Typhimurium: Critical Role for Lipopolysaccharide. J. Bacteriol. 2000, 182, 6308–6321. [Google Scholar] [CrossRef] [PubMed]
- Damton, N.C.; Turner, L.; Rojevsky, S.; Berg, H.C. Dynamics of Bacterial Swarming. Biophys. J. 2010, 98, 2082–2090. [Google Scholar] [CrossRef]
- Lai, S.; Tremblay, J.; Déziel, E. Swarming Motility: A Multicellular Behaviour Conferring Antimicrobial Resistance. Environ. Microbiol. 2009, 11, 126–136. [Google Scholar] [CrossRef]
- Poudel, S.; Giannone, R.J.; Farmer, A.T.; Campagna, S.R.; Bible, A.N.; Morrell-Falvey, J.L.; Elkins, J.G.; Hettich, R.L. Integrated Proteomics and Lipidomics Reveal That the Swarming Motility of Paenibacillus Polymyxa Is Characterized by Phospholipid Modification, Surfactant Deployment, and Flagellar Specialization Relative to Swimming Motility. Front. Microbiol. 2019, 10, 2594. [Google Scholar] [CrossRef] [PubMed]
- Carabelli, A.M.; Isgró, M.; Sanni, O.; Figueredo, G.P.; Winkler, D.A.; Burroughs, L.; Blok, A.J.; Dubern, J.F.; Pappalardo, F.; Hook, A.L.; et al. Single-Cell Tracking on Polymer Microarrays Reveals the Impact of Surface Chemistry on Pseudomonas Aeruginosa Twitching Speed and Biofilm Development. ACS Appl. Bio Mater. 2020, 3, 8471–8480. [Google Scholar] [CrossRef]
- Taylor, V.L.; Fitzpatrick, A.D.; Islam, Z.; Maxwell, K.L. The Diverse Impacts of Phage Morons on Bacterial Fitness and Virulence. In Advances in Virus Research; Academic Press Inc.: Cambridge, MA, USA, 2019; Volume 103, pp. 1–31. ISBN 9780128177228. [Google Scholar]
- Shanmugasundarasamy, T.; Karaiyagowder Govindarajan, D.; Kandaswamy, K. A Review on Pilus Assembly Mechanisms in Gram-Positive and Gram-Negative Bacteria. Cell Surf. 2022, 8, 100077. [Google Scholar] [CrossRef]
- Korotkov, K.V.; Sandkvist, M.; Hol, W.G.J. The Type II Secretion System: Biogenesis, Molecular Architecture and Mechanism. Nat. Rev. Microbiol. 2012, 10, 336–351. [Google Scholar] [CrossRef]
- Mercier, R.; Bautista, S.; Delannoy, M.; Gibert, M.; Guiseppi, A.; Herrou, J.; Mauriello, E.M.F.; Mignot, T. The Polar Ras-like GTPase MglA Activates Type IV Pilus via SgmX to Enable Twitching Motility in Myxococcus Xanthus. Proc. Natl. Acad. Sci. USA 2020, 117, 28366–28373. [Google Scholar] [CrossRef]
- Craig, L.; Forest, K.T.; Maier, B. Type IV Pili: Dynamics, Biophysics and Functional Consequences. Nat. Rev. Microbiol. 2019, 17, 429–440. [Google Scholar] [CrossRef]
- Jacobsen, T.; Bardiaux, B.; Francetic, O.; Izadi-Pruneyre, N.; Nilges, M. Structure and Function of Minor Pilins of Type IV Pili. Med. Microbiol. Immunol. 2020, 209, 301–308. [Google Scholar] [CrossRef]
- Chang, Y.W.; Rettberg, L.A.; Treuner-Lange, A.; Iwasa, J.; Søgaard-Andersen, L.; Jensen, G.J. Architecture of the Type IVa Pilus Machine. Science 2016, 351, aad2001. [Google Scholar] [CrossRef] [PubMed]
- Leong, C.G.; Bloomfield, R.A.; Boyd, C.A.; Dornbusch, A.J.; Lieber, L.; Liu, F.; Owen, A.; Slay, E.; Lang, K.M.; Lostroh, C.P. The Role of Core and Accessory Type IV Pilus Genes in Natural Transformation and Twitching Motility in the Bacterium Acinetobacter Baylyi. PLoS ONE 2017, 12, e0182139. [Google Scholar] [CrossRef] [PubMed]
- Zaburdaev, V.; Biais, N.; Schmiedeberg, M.; Eriksson, J.; Jonsson, A.B.; Sheetz, M.P.; Weitz, D.A. Uncovering the Mechanism of Trapping and Cell Orientation during Neisseria Gonorrhoeae Twitching Motility. Biophys. J. 2014, 107, 1523–1531. [Google Scholar] [CrossRef]
- Oliveira, N.M.; Foster, K.R.; Durham, W.M. Single-Cell Twitching Chemotaxis in Developing Biofilms. Proc. Natl. Acad. Sci. USA 2016, 113, 6532–6537. [Google Scholar] [CrossRef] [PubMed]
- Marathe, R.; Meel, C.; Schmidt, N.C.; Dewenter, L.; Kurre, R.; Greune, L.; Alexander Schmidt, M.; Müller, M.J.I.; Lipowsky, R.; Maier, B.; et al. Bacterial Twitching Motility Is Coordinated by a Two-Dimensional Tug-of-War with Directional Memory. Nat. Commun. 2014, 5, 3759. [Google Scholar] [CrossRef]
- Kühn, M.J.; Talà, L.; Inclan, Y.F.; Patino, R.; Pierrat, X.; Vos, I.; Al-Mayyah, Z.; Macmillan, H.; Negrete Jr, J.; Engel, J.N.; et al. Mechanotaxis Directs Pseudomonas Aeruginosa Twitching Motility. Proc. Natl. Acad. Sci. USA 2021, 118, e2101759118. [Google Scholar] [CrossRef]
- Xia, A.; Qian, M.; Wang, C.; Huang, Y.; Liu, Z.; Ni, L.; Jin, F. Optogenetic Modification of Pseudomonas Aeruginosa Enables Controllable Twitching Motility and Host Infection. ACS Synth. Biol. 2021, 10, 531–541. [Google Scholar] [CrossRef]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in Vivo Polymicrobial Biofilm Wound Infection Model to Study Interspecies Interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef]
- Wu, Y.K.; Cheng, N.C.; Cheng, C.M. Biofilms in Chronic Wounds: Pathogenesis and Diagnosis. Trends Biotechnol. 2019, 37, 505–517. [Google Scholar] [CrossRef]
- Schulze, A.; Mitterer, F.; Pombo, J.P.; Schild, S. Biofilms by Bacterial Human Pathogens: Clinical Relevance—Development, Composition and Regulation—Therapeutical Strategies. Microb. Cell 2021, 8, 28–56. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm Activity of Host Defence Peptides: Complexity Provides Opportunities. Nat. Rev. Microbiol. 2021, 19, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Nan, B. Bacterial Gliding Motility: Rolling Out a Consensus Model. Curr. Biol. 2017, 27, R154–R156. [Google Scholar] [CrossRef]
- Tchoufag, J.; Ghosh, P.; Pogue, C.B.; Nan, B.; Mandadapu, K.K. Mechanisms for Bacterial Gliding Motility on Soft Substrates. Proc. Natl. Acad. Sci. USA 2019, 116, 25087–25096. [Google Scholar] [CrossRef] [PubMed]
- Mauriello, E.M.F.; Mignot, T.; Yang, Z.; Zusman, D.R. Gliding Motility Revisited: How Do the Myxobacteria Move without Flagella? Microbiol. Mol. Biol. Rev. 2010, 74, 229–249. [Google Scholar] [CrossRef]
- Faure, L.M.; Fiche, J.B.; Espinosa, L.; Ducret, A.; Anantharaman, V.; Luciano, J.; Lhospice, S.; Islam, S.T.; Tréguier, J.; Sotes, M.; et al. The Mechanism of Force Transmission at Bacterial Focal Adhesion Complexes. Nature 2016, 539, 530–535. [Google Scholar] [CrossRef]
- Islam, S.T.; My, L.; Jolivet, N.Y.; Belgrave, A.M.; Fleuchot, B.; Brasseur, G.; Faure, L.M.; Sharma, G.; Lemon, D.J.; Saïdi, F.; et al. CglB Adhesins Secreted at Bacterial Focal Adhesions Mediate Gliding Motility. bioRxiv 2020, 1–70. [Google Scholar] [CrossRef]
- Nan, B.; Bandaria, J.N.; Moghtaderi, A.; Sun, I.H.; Yildiz, A.; Zusman, D.R. Flagella Stator Homologs Function as Motors for Myxobacterial Gliding Motility by Moving in Helical Trajectories. Proc. Natl. Acad. Sci. USA 2013, 110, E1508–E1513. [Google Scholar] [CrossRef] [PubMed]
- Nan, B.; Zusman, D.R. Novel Mechanisms Power Bacterial Gliding Motility. Mol. Microbiol. 2016, 101, 186–193. [Google Scholar] [CrossRef]
- Spormann, A.M. Gliding Motility in Bacteria: Insights from Studies of Myxococcus xanthus. Microbiol. Mol. Biol. Rev. 1999, 63, 621–641. [Google Scholar] [CrossRef]
- Martinez, A.; Torello, S.; Kolter, R. Sliding Motility in Mycobacteria. J. Bacteriol. 1999, 181, 7331–7338. [Google Scholar] [CrossRef]
- Stewart, C.R.; Rossier, O.; Cianciotto, N.P. Surface Translocation by Legionella Pneumophila: A Form of Sliding Motility That Is Dependent upon Type II Protein Secretion. J. Bacteriol. 2009, 191, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, T.; Kovács, Á.T. Sliding on the Surface: Bacterial Spreading without an Active Motor. Environ. Microbiol. 2017, 19, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Pollitt, E.J.G.; Crusz, S.A.; Diggle, S.P. Staphylococcus Aureus Forms Spreading Dendrites That Have Characteristics of Active Motility. Sci. Rep. 2015, 5, 17698. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| State | Microbial Activity and Host Response | Recommended Action |
|---|---|---|
| Contamination | Microbes enter the wound but there is no host response. | Observation |
| Colonisation | Microorganisms multiply without causing wound deterioration with only a weak host immune response. | |
| Critical colonisation | A large number of bacteria cause a delay in the wound healing process with a poor immune response of the host. | Intervention |
| Infection | A vast number of bacteria lead to delays in wound healing and trigger an often symptomatic inflammatory response caused by a strong immunological host response. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zegadło, K.; Gieroń, M.; Żarnowiec, P.; Durlik-Popińska, K.; Kręcisz, B.; Kaca, W.; Czerwonka, G. Bacterial Motility and Its Role in Skin and Wound Infections. Int. J. Mol. Sci. 2023, 24, 1707. https://doi.org/10.3390/ijms24021707
Zegadło K, Gieroń M, Żarnowiec P, Durlik-Popińska K, Kręcisz B, Kaca W, Czerwonka G. Bacterial Motility and Its Role in Skin and Wound Infections. International Journal of Molecular Sciences. 2023; 24(2):1707. https://doi.org/10.3390/ijms24021707
Chicago/Turabian StyleZegadło, Katarzyna, Monika Gieroń, Paulina Żarnowiec, Katarzyna Durlik-Popińska, Beata Kręcisz, Wiesław Kaca, and Grzegorz Czerwonka. 2023. "Bacterial Motility and Its Role in Skin and Wound Infections" International Journal of Molecular Sciences 24, no. 2: 1707. https://doi.org/10.3390/ijms24021707
APA StyleZegadło, K., Gieroń, M., Żarnowiec, P., Durlik-Popińska, K., Kręcisz, B., Kaca, W., & Czerwonka, G. (2023). Bacterial Motility and Its Role in Skin and Wound Infections. International Journal of Molecular Sciences, 24(2), 1707. https://doi.org/10.3390/ijms24021707