



Plasma microRNA Profiling in Type 2 Diabetes Mellitus: A Pilot Study

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
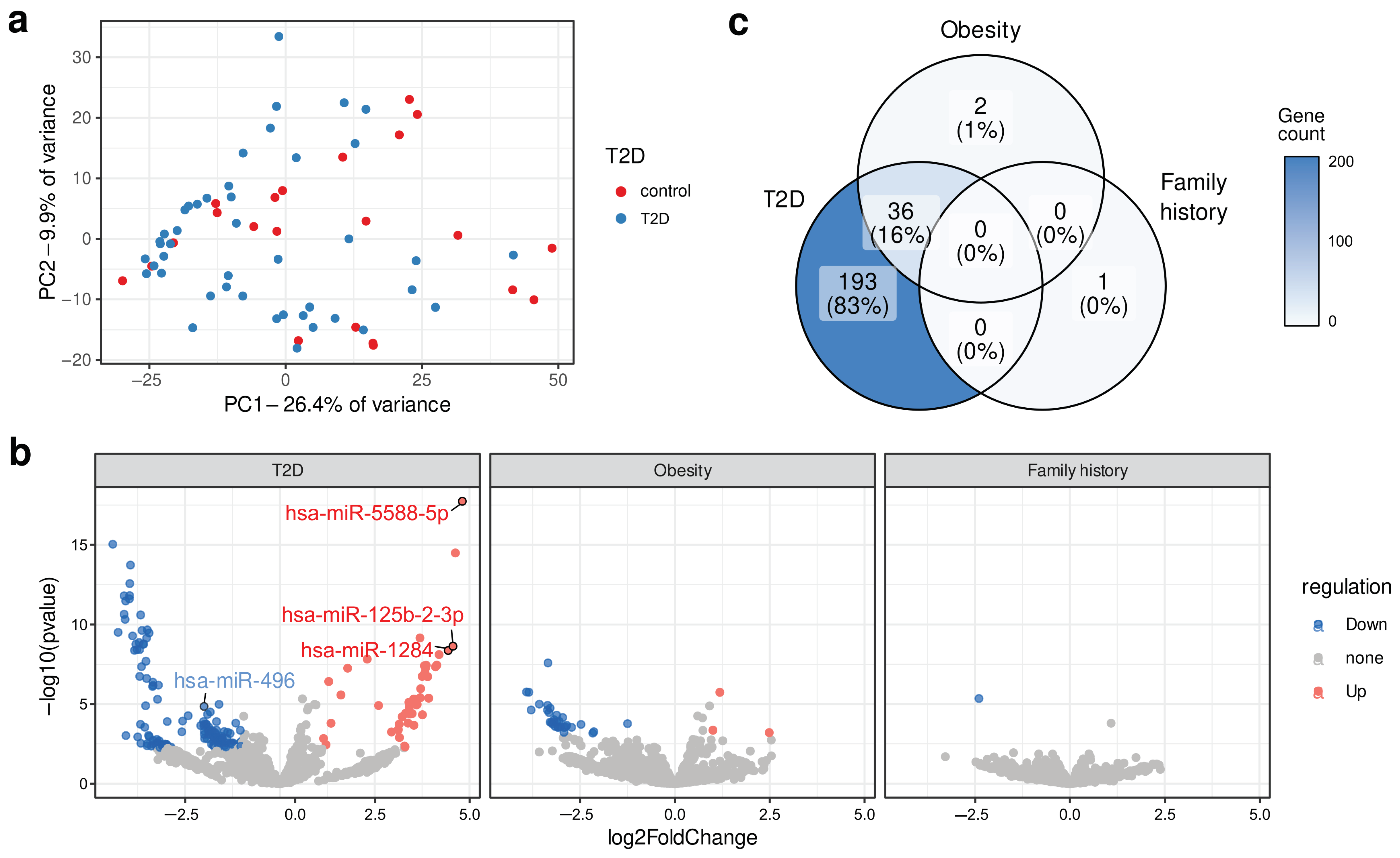
2.1. Identification of Differentially Expressed Circulating miRNA
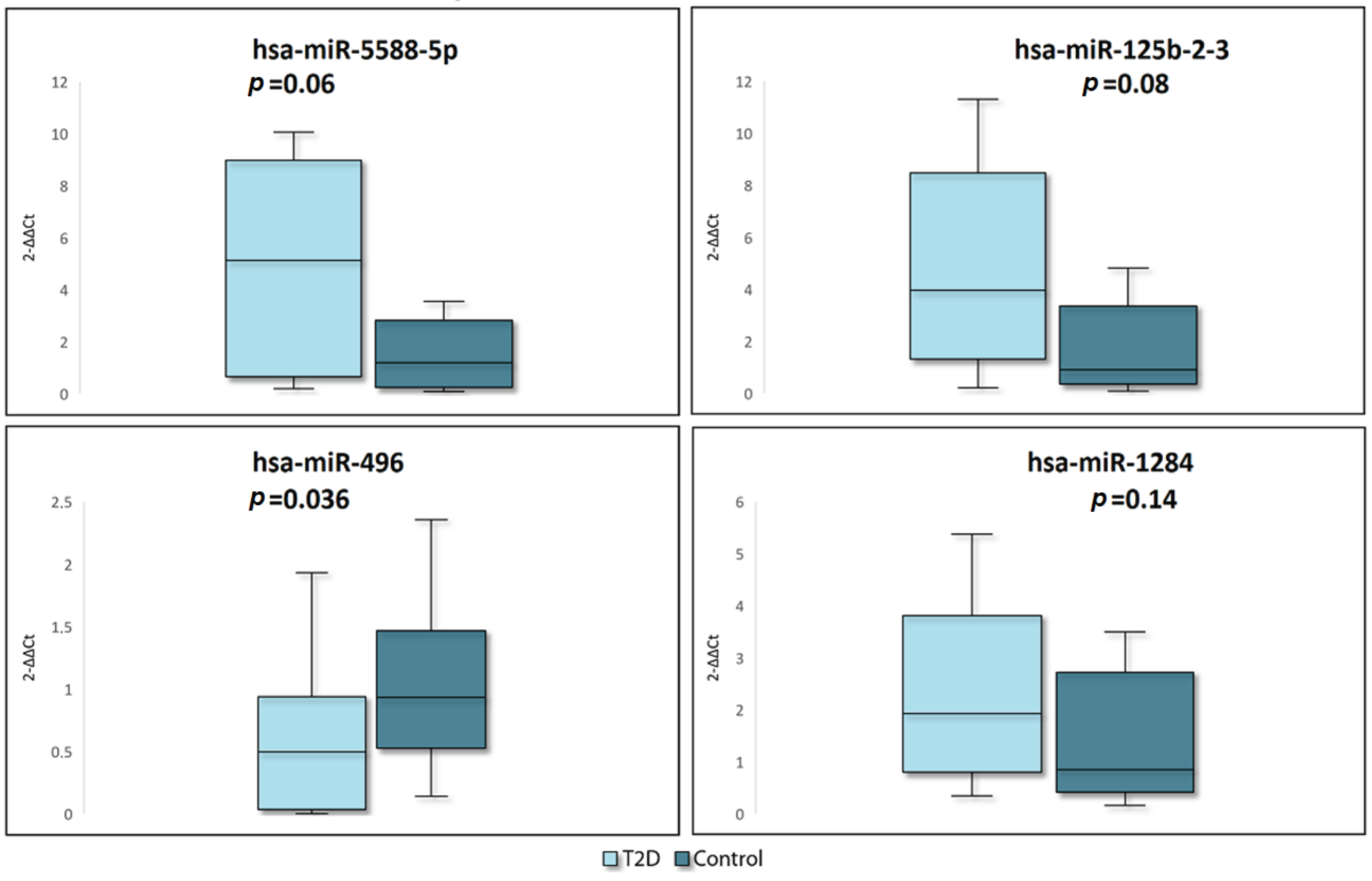
2.2. RT-qPCR Verification
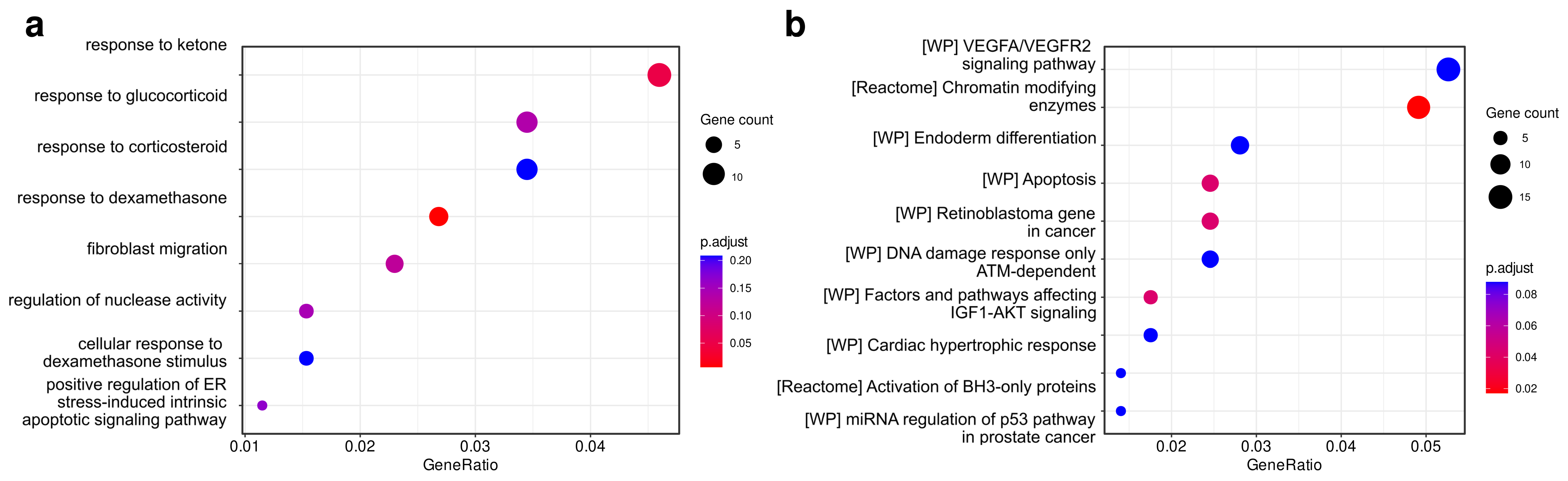
2.3. Identification and Functional Analysis of Putative miRNAs Target Genes
3. Discussion
4. Materials and Methods
4.1. Study Cohorts and Participants
4.2. Plasma Sample Collection
4.3. Small RNA Isolation
4.4. Small RNA Libraries Preparation and Sequencing
4.5. Differential Expression Analysis
4.6. Functional Analysis of miRNA Target Genes
4.7. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.8. Statistical Analysis
4.9. Machine Learning Model Construction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2009, 32, S62–S67. [CrossRef] [PubMed]
- Shehabi, Y.; Seppelt, I. Pro/Con Debate: Is Procalcitonin Useful for Guiding Antibiotic Decision Making in Critically Ill Patients? Crit. Care 2008, 12, 211. [Google Scholar] [CrossRef] [PubMed]
- Barnett, N.; Ware, L.B. Biomarkers in Acute Lung Injury—Marking Forward Progress. Crit. Care Clin. 2011, 27, 661–683. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kowdley, K.V. MicroRNAs in Common Human Diseases. Genom. Proteom. Bioinform. 2012, 10, 246–253. [Google Scholar] [CrossRef]
- Aryani, A.; Denecke, B. In Vitro Application of Ribonucleases: Comparison of the Effects on mRNA and miRNA Stability. BMC Res. Notes 2015, 8, 164. [Google Scholar] [CrossRef] [PubMed]
- Karolina, D.S.; Armugam, A.; Tavintharan, S.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. MicroRNA 144 Impairs Insulin Signaling by Inhibiting the Expression of Insulin Receptor Substrate 1 in Type 2 Diabetes Mellitus. PLoS ONE 2011, 6, e22839. [Google Scholar] [CrossRef]
- Zhang, T.; Lv, C.; Li, L.; Chen, S.; Liu, S.; Wang, C.; Su, B. Plasma miR-126 Is a Potential Biomarker for Early Prediction of Type 2 Diabetes Mellitus in Susceptible Individuals. BioMed Res. Int. 2013, 2013, 761617. [Google Scholar] [CrossRef] [PubMed]
- Massart, J.; Sjögren, R.J.O.; Lundell, L.S.; Mudry, J.M.; Franck, N.; O’Gorman, D.J.; Egan, B.; Zierath, J.R.; Krook, A. Altered miR-29 Expression in Type 2 Diabetes Influences Glucose and Lipid Metabolism in Skeletal Muscle. Diabetes 2017, 66, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Mone, P.; De Donato, A.; Varzideh, F.; Kansakar, U.; Jankauskas, S.S.; Pansini, A.; Santulli, G. Functional Role of miR-34a in Diabetes and Frailty. Front. Aging 2022, 3, 949924. [Google Scholar] [CrossRef]
- The InterAct Consortium. The Link between Family History and Risk of Type 2 Diabetes Is Not Explained by Anthropometric, Lifestyle or Genetic Risk Factors: The EPIC-InterAct Study. Diabetologia 2013, 56, 60–69. [Google Scholar] [CrossRef]
- Klein, S.; Gastaldelli, A.; Yki-Järvinen, H.; Scherer, P.E. Why Does Obesity Cause Diabetes? Cell Metab. 2022, 34, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Tonyan, Z.N.; Nasykhova, Y.A.; Danilova, M.M.; Barbitoff, Y.A.; Changalidi, A.I.; Mikhailova, A.A.; Glotov, A.S. Overview of Transcriptomic Research on Type 2 Diabetes: Challenges and Perspectives. Genes 2022, 13, 1176. [Google Scholar] [CrossRef]
- Kim, M.; Zhang, X. The Profiling and Role of miRNAs in Diabetes Mellitus. J. Diabetes Clin. Res. 2019, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Granjon, A.; Gustin, M.-P.; Rieusset, J.; Lefai, E.; Meugnier, E.; Güller, I.; Cerutti, C.; Paultre, C.; Disse, E.; Rabasa-Lhoret, R.; et al. The microRNA Signature in Response to Insulin Reveals Its Implication in the Transcriptional Action of Insulin in Human Skeletal Muscle and the Role of a Sterol Regulatory Element–Binding Protein-1c/Myocyte Enhancer Factor 2C Pathway. Diabetes 2009, 58, 2555–2564. [Google Scholar] [CrossRef]
- Klöting, N.; Berthold, S.; Kovacs, P.; Schön, M.R.; Fasshauer, M.; Ruschke, K.; Stumvoll, M.; Blüher, M. MicroRNA Expression in Human Omental and Subcutaneous Adipose Tissue. PLoS ONE 2009, 4, e4699. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.J.; Moreno-Navarrete, J.M.; Pardo, G.; Sabater, M.; Hummel, M.; Ferrer, A.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. MiRNA Expression Profile of Human Subcutaneous Adipose and during Adipocyte Differentiation. PLoS ONE 2010, 5, e9022. [Google Scholar] [CrossRef]
- Locke, J.M.; Da Silva Xavier, G.; Dawe, H.R.; Rutter, G.A.; Harries, L.W. Increased Expression of miR-187 in Human Islets from Individuals with Type 2 Diabetes Is Associated with Reduced Glucose-Stimulated Insulin Secretion. Diabetologia 2014, 57, 122–128. [Google Scholar] [CrossRef]
- Sebastiani, G.; Po, A.; Miele, E.; Ventriglia, G.; Ceccarelli, E.; Bugliani, M.; Marselli, L.; Marchetti, P.; Gulino, A.; Ferretti, E.; et al. MicroRNA-124a Is Hyperexpressed in Type 2 Diabetic Human Pancreatic Islets and Negatively Regulates Insulin Secretion. Acta Diabetol. 2015, 52, 523–530. [Google Scholar] [CrossRef]
- Latouche, C.; Natoli, A.; Reddy-Luthmoodoo, M.; Heywood, S.E.; Armitage, J.A.; Kingwell, B.A. MicroRNA-194 Modulates Glucose Metabolism and Its Skeletal Muscle Expression Is Reduced in Diabetes. PLoS ONE 2016, 11, e0155108. [Google Scholar] [CrossRef] [PubMed]
- Pek, S.L.T.; Sum, C.F.; Lin, M.X.; Cheng, A.K.S.; Wong, M.T.K.; Lim, S.C.; Tavintharan, S. Circulating and Visceral Adipose miR-100 Is down-Regulated in Patients with Obesity and Type 2 Diabetes. Mol. Cell Endocrinol. 2016, 427, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Dahlmans, D.; Houzelle, A.; Jörgensen, J.A.; Phielix, E.; Lindeboom, L.; Hesselink, M.K.C.; Schrauwen, P.; Hoeks, J. Evaluation of Muscle microRNA Expression in Relation to Human Peripheral Insulin Sensitivity: A Cross-Sectional Study in Metabolically Distinct Subject Groups. Front. Physiol. 2017, 8, 711. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Guan, M. miRNA-1284, a Regulator of HMGB1, Inhibits Cell Proliferation and Migration in Osteosarcoma. Biosci. Rep. 2018, 38, BSR20171675. [Google Scholar] [CrossRef] [PubMed]
- Qi, N.-N.; Tian, S.; Li, X.; Wang, F.-L.; Liu, B. Up-Regulation of microRNA-496 Suppresses Proliferation, Invasion, Migration and in Vivo Tumorigenicity of Human Osteosarcoma Cells by Targeting eIF4E. Biochimie 2019, 163, 1–11. [Google Scholar] [CrossRef]
- Wei, W.; Cao, W.; Zhan, Z.; Yan, L.; Xie, Y.; Xiao, Q. MiR-1284 Suppresses Gastric Cancer Progression by Targeting EIF4A1. OTT 2019, 12, 3965–3976. [Google Scholar] [CrossRef] [PubMed]
- De Felice, B.; Montanino, C.; Oliva, M.; Bonavita, S.; Di Onofrio, V.; Coppola, C. MicroRNA Expression Signature in Mild Cognitive Impairment Due to Alzheimer’s Disease. Mol. Neurobiol. 2020, 57, 4408–4416. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Q.; Chen, G.; Xiong, D.-D.; Lai, Z.-F.; Liu, L.-M.; Fang, Y.-Y.; Shen, J.-H.; Gan, X.-Y.; Liao, L.-F.; Dang, Y.-W. Down-Regulation of microRNA-125b-2-3p Is a Risk Factor for a Poor Prognosis in Hepatocellular Carcinoma. Bioengineered 2021, 12, 1627–1641. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zhu, P.; Liu, S.; Gao, B.; Wang, W. miR-496 Suppress Tumorigenesis via Targeting BDNF-Mediated PI3K/Akt Signaling Pathway in Non-Small Cell Lung Cancer. Biochem. Biophys. Res. Commun. 2019, 518, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Zhao, E.; Zhang, J. miR-496 Inhibits Proliferation via LYN and AKT Pathway in Gastric Cancer. Open. Med. 2021, 16, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Rubie, C.; Zimmer, J.; Lammert, F.; Gross, J.C.; Weber, S.N.; Kruse, B.; Halajda, B.; Wagner, M.; Wagenpfeil, S.; Glanemann, M. MicroRNA-496 and Mechanistic Target of Rapamycin Expression Are Associated with Type 2 Diabetes Mellitus and Obesity in Elderly People. Ann. Nutr. Metab. 2019, 74, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Rubie, C.; Kölsch, K.; Halajda, B.; Eichler, H.; Wagenpfeil, S.; Roemer, K.; Glanemann, M. microRNA-496—A New, Potentially Aging-Relevant Regulator of mTOR. Cell Cycle 2016, 15, 1108–1116. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W. Role of mTOR in Glucose and Lipid Metabolism. IJMS 2018, 19, 2043. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-B.; Liu, J.-L.; Wang, W.; Li, R.-Y.; Yu, D.-J.; Lan, X.-Y.; Li, J.-P. Plasma Exosomal miR-422a and miR-125b-2-3p Serve as Biomarkers for Ischemic Stroke. CNR 2018, 14, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Lu, J.; Wang, Y.; Sheng, H.; Wang, Y.; Chen, Z.; Wu, Q.; Zheng, J.; Chen, Y.; Yang, D.; et al. The lncRNA XIST/miR-125b-2-3p Axis Modulates Cell Proliferation and Chemotherapeutic Sensitivity via Targeting Wee1 in Colorectal Cancer. Cancer Med. 2021, 10, 2423–2441. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, G.; Jiang, Y. The Emerging Roles of miR-125b in Cancers. CMAR 2020, 12, 1079–1088. [Google Scholar] [CrossRef]
- Long, Y.C. AMP-Activated Protein Kinase Signaling in Metabolic Regulation. J. Clin. Investig. 2006, 116, 1776–1783. [Google Scholar] [CrossRef]
- Cheung, R.; Pizza, G.; Rolando, D.M.; Chabosseau, P.L.; Nguyen-Tu, M.-S.; Leclerc, I.; Rutter, G.A.; Martinez-Sanchez, A. 2183-P: miR-125b Is Regulated by Glucose via AMPK and Impairs ß-Cell Function. Diabetes 2019, 68, 2183-P. [Google Scholar] [CrossRef]
- Cheung, R.; Pizza, G.; Chabosseau, P.; Rolando, D.; Tomas, A.; Burgoyne, T.; Wu, Z.; Salowka, A.; Thapa, A.; Macklin, A.; et al. Glucose-Dependent miR-125b Is a Negative Regulator of β-Cell Function. Diabetes 2022, 71, 1525–1545. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, H.; Pan, X.; Wu, W.; Wang, H.; Yan, L.; Zhang, M.; Liu, X.; Xia, S.; Shao, Q. miR-34a and miR-125b Are Upregulated in Peripheral Blood Mononuclear Cells from Patients with Type 2 Diabetes Mellitus. Exp. Ther. Med. 2017, 14, 5589–5596. [Google Scholar] [CrossRef] [PubMed]
- Strycharz, J.; Wróblewski, A.; Zieleniak, A.; Świderska, E.; Matyjas, T.; Rucińska, M.; Pomorski, L.; Czarny, P.; Szemraj, J.; Drzewoski, J.; et al. Visceral Adipose Tissue of Prediabetic and Diabetic Females Shares a Set of Similarly Upregulated microRNAs Functionally Annotated to Inflammation, Oxidative Stress and Insulin Signaling. Antioxidants 2021, 10, 101. [Google Scholar] [CrossRef]
- Giroud, M.; Pisani, D.F.; Karbiener, M.; Barquissau, V.; Ghandour, R.A.; Tews, D.; Fischer-Posovszky, P.; Chambard, J.-C.; Knippschild, U.; Niemi, T.; et al. miR-125b Affects Mitochondrial Biogenesis and Impairs Brite Adipocyte Formation and Function. Mol. Metab. 2016, 5, 615–625. [Google Scholar] [CrossRef]
- Wei, L.-M.; Sun, R.-P.; Dong, T.; Liu, J.; Chen, T.; Zeng, B.; Wu, J.-H.; Luo, J.; Sun, J.-J.; Xi, Q.-Y.; et al. MiR-125b-2 Knockout Increases High-Fat Diet-Induced Fat Accumulation and Insulin Resistance. Sci. Rep. 2020, 10, 21969. [Google Scholar] [CrossRef]
- Patnaik, S.K.; Yendamuri, S.; Kannisto, E.; Kucharczuk, J.C.; Singhal, S.; Vachani, A. MicroRNA Expression Profiles of Whole Blood in Lung Adenocarcinoma. PLoS ONE 2012, 7, e46045. [Google Scholar] [CrossRef]
- Li, J.; Jin, H.; Yu, H.; Wang, B.; Tang, J. miRNA-1284 Inhibits Cell Growth and Induces Apoptosis of Lung Cancer Cells. Mol. Med. Rep. 2017, 16, 3049–3054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yang, F.; Luo, Q.; Yan, D.; Sun, S. miR-1284 Inhibits the Growth and Invasion of Breast Cancer Cells by Targeting ZIC2. Oncol. Res. 2019, 27, 253–260. [Google Scholar] [CrossRef]
- Yi, W.; Wang, J.; Yao, Z.; Kong, Q.; Zhang, N.; Mo, W.; Xu, L.; Li, X. The Expression Status of ZIC2 as a Prognostic Marker for Nasopharyngeal Carcinoma. Int. J. Clin. Exp. Pathol. 2018, 11, 4446–4460. [Google Scholar] [PubMed]
- Pan, C.; Wang, D.; Zhang, Y.; Yu, W. MicroRNA-1284 Inhibits Cell Viability and Induces Apoptosis of Ovarian Cancer Cell Line OVCAR3. Oncol. Res. 2016, 24, 429–435. [Google Scholar] [CrossRef]
- Georgia, S.; Bhushan, A. P27 Regulates the Transition of β-Cells From Quiescence to Proliferation. Diabetes 2006, 55, 2950–2956. [Google Scholar] [CrossRef] [PubMed]
- Rachdi, L.; Balcazar, N.; Elghazi, L.; Barker, D.J.; Krits, I.; Kiyokawa, H.; Bernal-Mizrachi, E. Differential Effects of P27 in Regulation of β-Cell Mass During Development, Neonatal Period, and Adult Life. Diabetes 2006, 55, 3520–3528. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, Q.; Wang, J.; Zhao, Y.; Niu, L.; Bai, L.; Shuai, S.; Li, X.; Zhang, S.; Zhu, L. miR-144-3p Promotes Adipogenesis Through Releasing C/EBPα From Klf3 and CtBP2. Front. Genet. 2018, 9, 677. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Tang, Y.; Zhao, Y.; Zhao, J.; Zhang, L.; Wei, W.; Chen, J. MiR-144-3p Targets FoxO1 to Reduce Its Regulation of Adiponectin and Promote Adipogenesis. Front. Genet. 2020, 11, 603144. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.; Reddy, S.S.; Maurya, M.; Maurya, P.; Barthwal, M.K. MicroRNA-99a Mimics Inhibit M1 Macrophage Phenotype and Adipose Tissue Inflammation by Targeting TNFα. Cell Mol. Immunol. 2019, 16, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Rakib, A.; Kiran, S.; Mandal, M.; Singh, U.P. MicroRNAs: A Crossroad That Connects Obesity to Immunity and Aging. Immun. Ageing 2022, 19, 64. [Google Scholar] [CrossRef]
- Keller, M.P.; Choi, Y.; Wang, P.; Belt Davis, D.; Rabaglia, M.E.; Oler, A.T.; Stapleton, D.S.; Argmann, C.; Schueler, K.L.; Edwards, S.; et al. A Gene Expression Network Model of Type 2 Diabetes Links Cell Cycle Regulation in Islets with Diabetes Susceptibility. Genome Res. 2008, 18, 706–716. [Google Scholar] [CrossRef]
- Deng, B.; Song, A.; Zhang, C. Cell-Cycle Dysregulation in the Pathogenesis of Diabetic Kidney Disease: An Update. IJMS 2023, 24, 2133. [Google Scholar] [CrossRef] [PubMed]
- Tomita, T. Apoptosis in Pancreatic β-Islet Cells in Type 2 Diabetes. Bosn. J. Basic. Med. Sci. 2016, 16, 162–179. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef]
- Schones, D.E.; Leung, A.; Natarajan, R. Chromatin Modifications Associated With Diabetes and Obesity. ATVB 2015, 35, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhuang, Z.; Wang, W.; Huang, T.; Liu, Z. An Improved Genome-Wide Polygenic Score Model for Predicting the Risk of Type 2 Diabetes. Front. Genet. 2021, 12, 632385. [Google Scholar] [CrossRef]
- Tillil, H.; Köbberling, J. Age-Corrected Empirical Genetic Risk Estimates for First-Degree Relatives of IDDM Patients. Diabetes 1987, 36, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, X.-P.; Yuan, J.; Cai, B.; Wang, X.-L.; Wu, X.-L.; Zhang, Y.-H.; Zhang, X.-Y.; Yin, T.; Zhu, X.-H.; et al. Association of Body Mass Index and Age with Incident Diabetes in Chinese Adults: A Population-Based Cohort Study. BMJ Open. 2018, 8, e021768. [Google Scholar] [CrossRef] [PubMed]
- Karin, A.; Jon, E.; Martin, A.; Lena, B.; Martin, L.; Naveed, S.; Marcus, L.; Maria, Å.; Annika, R. Body Mass Index in Adolescence, Risk of Type 2 Diabetes and Associated Complications: A Nationwide Cohort Study of Men. eClinicalMedicine 2022, 46, 101356. [Google Scholar] [CrossRef] [PubMed]
- Kautzky-Willer, A.; Leutner, M.; Harreiter, J. Sex Differences in Type 2 Diabetes. Diabetologia 2023, 66, 986–1002. [Google Scholar] [CrossRef] [PubMed]
- Shaker, O.G.; Abdelaleem, O.O.; Mahmoud, R.H.; Abdelghaffar, N.K.; Ahmed, T.I.; Said, O.M.; Zaki, O.M. Diagnostic and Prognostic Role of Serum miR-20b, miR-17-3p, HOTAIR, and MALAT1 in Diabetic Retinopathy. IUBMB Life 2019, 71, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Li, S.; Li, J.; Yin, L.; Zhou, T.; Zhang, C.; Chen, X.; Sun, K. Ethnic Differences in MicroRNA-375 Expression Level and DNA Methylation Status in Type 2 Diabetes of Han and Kazak Populations. J. Diabetes Res. 2014, 2014, 761938. [Google Scholar] [CrossRef]
- Scholes, A.N.; Lewis, J.A. Comparison of RNA Isolation Methods on RNA-Seq: Implications for Differential Expression and Meta-Analyses. BMC Genom. 2020, 21, 249. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dedov, I.I.; Shestakova, M.V.; Vikulova, O.K.; Zheleznyakova, A.V.; Isakov, M.A.; Sazonova, D.V.; Mokrysheva, N.G. Diabetes Mellitus in the Russian Federation: Dynamics of Epidemiological Indicators According to the Federal Register of Diabetes Mellitus for the Period 2010–2022. Diabetes Mellit. 2023, 26, 104–123. [Google Scholar] [CrossRef]
- Charan, J.; Kaur, R.; Bhardwaj, P.; Singh, K.; Ambwani, S.R.; Misra, S. Sample Size Calculation in Medical Research: A Primer. Ann. Natl. Acad. Med. Sci. 2021, 57, 74–80. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MiRNA | log2FC | Adjusted p-Value | Associated Conditions | Tissues/Biological Fluids | Pathways | References |
|---|---|---|---|---|---|---|
| miR-5588-5p | 4.81 | 3.7 × 10−15 | Alzheimer’s disease | Whole blood, serum, cerebrospinal fluid, plasma | Further studies are required | [28] |
| miR-125b-2-3p | 4.43 | 3.1 × 10−7 | Hepatocellular carcinoma, Ischemic stroke, colorectal cancer | Islet cells, visceral adipose tissue, PBMC, plasma | AMPK pathway | [29,35,36,37,38,39,40,41,42,43,44] |
| miR-1284 | 4.20 | 5.4 × 10−7 | Breast, gastric, lung cancer cancer, osteosarcoma | Tumor tissues, plasma | PI3K/Akt, p27 pathways | [45,46,47,48,49,50,51] |
| miR-496 | −3.90 | 4.4 × 10−4 | Osteosarcoma, T2D | PBMC, plasma | PI3K/AKT, mTOR pathway | [30,31,32,33,34] |
| Characteristics | T2D Patients (n = 44) | Healthy Controls (n = 22) |
|---|---|---|
| Male (n) | 14 | 11 |
| Female (n) | 30 | 11 |
| Age (years) | 73.15 ± 7.72 * | 42.90 ± 16.52 |
| FBG (mmol/L) | 8.34 ± 2.64 * | 4.62 ± 0.39 |
| Family history of diabetes (n) | 17 (38.6%) | 8 (36.3%) |
| HbA1c (%) | 8.36 ± 1.42 | NA |
| HDL (mmol/L) | 1.29 ± 0.33 * | 1.60 ± 0.54 |
| LDL (mmol/L) | 2.62 ± 0.88 | 3.35 ± 1.69 |
| Total cholesterol (mmol/L) | 4.59 ± 1.14 | 5.57 ± 1.79 |
| Creatinine (mmol/L) | 0.115 ± 0.03 * | 0.083 ± 0.01 |
| BMI (kg/m2) | 30.25 ± 4.98 * | 23.98 ± 2.23 |
| WHR | 0.95 ± 0.08 * | 0.75 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonyan, Z.N.; Barbitoff, Y.A.; Nasykhova, Y.A.; Danilova, M.M.; Kozyulina, P.Y.; Mikhailova, A.A.; Bulgakova, O.L.; Vlasova, M.E.; Golovkin, N.V.; Glotov, A.S. Plasma microRNA Profiling in Type 2 Diabetes Mellitus: A Pilot Study. Int. J. Mol. Sci. 2023, 24, 17406. https://doi.org/10.3390/ijms242417406
Tonyan ZN, Barbitoff YA, Nasykhova YA, Danilova MM, Kozyulina PY, Mikhailova AA, Bulgakova OL, Vlasova ME, Golovkin NV, Glotov AS. Plasma microRNA Profiling in Type 2 Diabetes Mellitus: A Pilot Study. International Journal of Molecular Sciences. 2023; 24(24):17406. https://doi.org/10.3390/ijms242417406
Chicago/Turabian StyleTonyan, Ziravard N., Yury A. Barbitoff, Yulia A. Nasykhova, Maria M. Danilova, Polina Y. Kozyulina, Anastasiia A. Mikhailova, Olga L. Bulgakova, Margarita E. Vlasova, Nikita V. Golovkin, and Andrey S. Glotov. 2023. "Plasma microRNA Profiling in Type 2 Diabetes Mellitus: A Pilot Study" International Journal of Molecular Sciences 24, no. 24: 17406. https://doi.org/10.3390/ijms242417406
APA StyleTonyan, Z. N., Barbitoff, Y. A., Nasykhova, Y. A., Danilova, M. M., Kozyulina, P. Y., Mikhailova, A. A., Bulgakova, O. L., Vlasova, M. E., Golovkin, N. V., & Glotov, A. S. (2023). Plasma microRNA Profiling in Type 2 Diabetes Mellitus: A Pilot Study. International Journal of Molecular Sciences, 24(24), 17406. https://doi.org/10.3390/ijms242417406