
PDGFRβ Activation Induced the Bovine Embryonic Genome Activation via Enhanced NFYA Nuclear Localization

 ,
,  , , and
, , and
Abstract
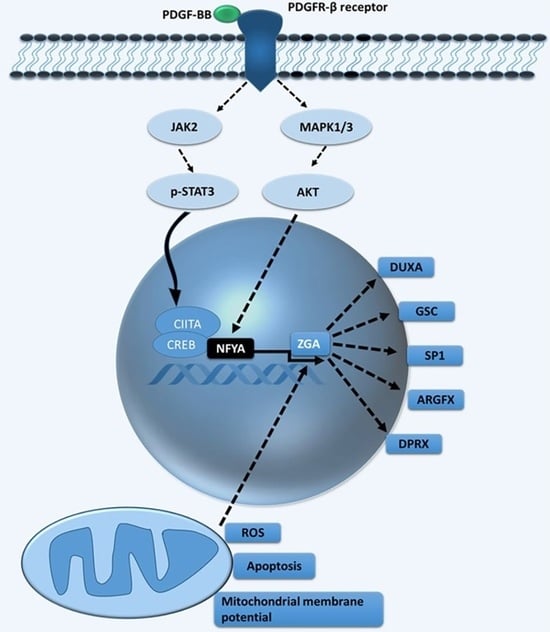
1. Introduction
2. Results
2.1. Dynamic Changes in PDGFRβ and NFYA Expression during Bovine Embryonic Development
2.2. Effect of PDGFRβ on NFYA and Embryonic Development
2.3. PDGFRβ and NFYA Are Linked to Embryonic Genome Activation
2.4. MAPK1/AKT and JAK/STAT Pathway Roles in NFYA Nuclear Localization via PDGFRβ Activation
2.5. PDGFRβ and NFYA Effects on Embryonic Mitochondria and Implantation Potential
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.1.1. Experiment 1 (PDGFRβ Activation)
4.1.2. Experiment 2 (PDGFRβ Inhibition)
4.2. Oocyte Collection and In Vitro Maturation (IVM)
4.3. In Vitro Fertilization (IVF) and Culture (IVC) of Embryos
4.4. Immunofluorescence Analysis and Antibodies
4.5. Detection of Embryonic Reactive Oxygen Species (ROS)
4.6. TUNEL Assay
4.7. JC-1 Staining
4.8. Invasion Assay
4.9. mRNA Extraction of and Complementary DNA (cDNA) Synthesis Real-Time Quantitative PCR (qRT-PCR) Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilcox, A.J.; Harmon, Q.; Doody, K.; Wolf, D.P.; Adashi, E.Y. Preimplantation loss of fertilized human ova: Estimating the unobservable. Hum. Reprod. 2020, 35, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.N.; Harrison, M.M. Mechanisms regulating zygotic genome activation. Nat. Rev. Genet. 2019, 20, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Bonneau, A.R.; Giraldez, A.J. Zygotic genome activation during the maternal-to-zygotic transition. Annu. Rev. Cell Dev. Biol. 2014, 30, 581–613. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J.; Gebuhr, T.C.; Pan, H.; Svoboda, P.; Schultz, R.M.; Magnuson, T. Maternal BRG1 regulates zygotic genome activation in the mouse. Genes Dev. 2006, 20, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Asami, M.; Lam, B.Y.; Ma, M.K.; Rainbow, K.; Braun, S.; VerMilyea, M.D.; Yeo, G.S.; Perry, A.C. Human embryonic genome activation initiates at the one-cell stage. Cell Stem Cell 2022, 29, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Sha, Q.-Q.; Zhang, J.; Fan, H.-Y. A story of birth and death: mRNA translation and clearance at the onset of maternal-to-zygotic transition in mammals. Biol. Reprod. 2019, 101, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Memili, E.; First, N.L. Zygotic and embryonic gene expression in cow: A review of timing and mechanisms of early gene expression as compared with other species. Zygote 2000, 8, 87–96. [Google Scholar] [CrossRef]
- Jukam, D.; Shariati, S.A.M.; Skotheim, J.M. Zygotic genome activation in vertebrates. Dev. Cell 2017, 42, 316–332. [Google Scholar] [CrossRef]
- Collart, C.; Owens, N.D.L.; Bhaw-Rosun, L.; Cooper, B.; De Domenico, E.; Patrushev, I.; Sesay, A.K.; Smith, J.C.; Gilchrist, M.J. High-resolution analysis of gene activity during the Xenopus mid-blastula transition. Development 2014, 141, 1927–1939. [Google Scholar] [CrossRef]
- Wu, E.; Vastenhouw, N.L. From mother to embryo: A molecular perspective on zygotic genome activation. Curr. Top. Dev. Biol. 2020, 140, 209–254. [Google Scholar]
- Halstead, M.M.; Ma, X.; Zhou, C.; Schultz, R.M.; Ross, P.J. Chromatin remodeling in bovine embryos indicates species-specific regulation of genome activation. Nat. Commun. 2020, 11, 4654. [Google Scholar] [CrossRef]
- Ribeiro, L.; Tobias-Santos, V.; Santos, D.; Antunes, F.; Feltran, G.; Menezes, J.d.S.; Aravind, L.; Venancio, T.M.; da Fonseca, R.N. Evolution and multiple roles of the Pancrustacea specific transcription factor zelda in insects. PLoS Genet. 2017, 13, e1006868. [Google Scholar] [CrossRef]
- Okuda, Y.; Yoda, H.; Uchikawa, M.; Furutani-Seiki, M.; Takeda, H.; Kondoh, H.; Kamachi, Y. Comparative genomic and expression analysis of group B1 sox genes in zebrafish indicates their diversification during vertebrate evolution. Dev. Dyn. 2006, 235, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zhang, C.; Zhang, Y. Epigenetic regulation of mouse preimplantation embryo development. Curr. Opin. Genet. Dev. 2020, 64, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Liu, Y.; Inoue, A.; Suzuki, T.; Zhao, K.; Zhang, Y. Establishing chromatin regulatory landscape during mouse preimplantation development. Cell 2016, 165, 1375–1388. [Google Scholar] [CrossRef]
- Li, G.; Zhao, H.; Wang, L.; Wang, Y.; Guo, X.; Xu, B. The animal nuclear factor Y: An enigmatic and important heterotrimeric transcription factor. Am. J. Cancer Res. 2018, 8, 1106. [Google Scholar] [PubMed]
- Cai, J.; Chen, H.; Xie, S.; Hu, Z.; Bai, Y. Research Progress of Totipotent Stem Cells. Stem Cells Dev. 2022, 31, 335–345. [Google Scholar] [CrossRef]
- Rengaraj, D.; Hwang, Y.S.; Lee, H.C.; Han, J.Y. Zygotic genome activation in the chicken: A comparative review. Cell. Mol. Life Sci. 2020, 77, 1879–1891. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. Signal transduction from the cell surface to the nucleus through the phosphorylation of transcription factors. Curr. Opin. Cell Biol. 1994, 6, 415–424. [Google Scholar] [CrossRef]
- Chen, P.H.; Chen, X.; He, X. Platelet-derived growth factors and their receptors: Structural and functional perspectives. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2013, 1834, 2176–2186. [Google Scholar] [CrossRef]
- Noskovičová, N.; Petřek, M.; Eickelberg, O.; Heinzelmann, K. Platelet-derived growth factor signaling in the lung. From lung development and disease to clinical studies. Am. J. Respir. Cell Mol. Biol. 2015, 52, 263–284. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; Fernández, P.; Esteban, V.; Pello, M.; Indolfi, C.; Rodríguez, C.; Rodríguez-Calvo, R.; López-Maderuelo, M.D.; Bauriedel, G.; Hutter, R.; et al. Inactivation of nuclear factor-Y inhibits vascular smooth muscle cell proliferation and neointima formation. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, W.; Tachibana, K. Awakening of the zygotic genome by pioneer transcription factors. Curr. Opin. Struct. Biol. 2021, 71, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Vallot, A.; Tachibana, K. The emergence of genome architecture and zygotic genome activation. Curr. Opin. Cell Biol. 2020, 64, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, C.; Liu, X.; Li, C.; Wu, Q.; Chen, X.; Yang, L.; Kou, X.; Zhao, Y.; Wang, H.; et al. Dynamic nucleosome organization after fertilization reveals regulatory factors for mouse zygotic genome activation. Cell Res. 2022, 32, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Fischer, B. Platelet-Derived Growth Factor-Receptor-β in Pig and Cattle Blastocysts. Cells Tissues Organs 1997, 159, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Frontini, M.; Imbriano, C.; Manni, I.; Mantovani, R. Cell-cycle regulation of NF-YC nuclear localization. Cell Cycle 2004, 3, 205–210. [Google Scholar] [CrossRef]
- Alabert, C.; Rogers, L.; Kahn, L.; Niellez, S.; Fafet, P.; Cerulis, S.; Blanchard, J.M.; A Hipskind, R.; Vignais, M.-L. Cell type-dependent control of NF-Y activity by TGF-β. Oncogene 2006, 25, 3387–3396. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, M.; DuHadaway, J.B.; Prendergast, G.C.; Laury-Kleintop, L.D. RhoB regulates PDGFR-β trafficking and signaling in vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2597–2605. [Google Scholar] [CrossRef]
- Stanney, W., III; Ladam, F.; Donaldson, I.J.; Parsons, T.J.; Maehr, R.; Bobola, N.; Sagerström, C.G. Combinatorial action of NF–Y and TALE at embryonic enhancers defines distinct gene expression programs during zygotic genome activation in zebrafish. Dev. Biol. 2020, 459, 161–180. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Fiorenza, M.T.; Mangia, F. A developmentally regulated GAGA box-binding factor and Sp1 are required for transcription of the hsp70. 1 gene at the onset of mouse zygotic genome activation. Development 2000, 127, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Bentsen, M.; Goymann, P.; Schultheis, H.; Klee, K.; Petrova, A.; Wiegandt, R.; Fust, A.; Preussner, J.; Kuenne, C.; Braun, T.; et al. ATAC-seq footprinting unravels kinetics of transcription factor binding during zygotic genome activation. Nat. Commun. 2020, 11, 4267. [Google Scholar] [CrossRef] [PubMed]
- De Iaco, A.; Planet, E.; Coluccio, A.; Verp, S.; Duc, J.; Trono, D. DUX-family transcription factors regulate zygotic genome activation in placental mammals. Nat. Genet. 2017, 49, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Ortega, N.M.; Winblad, N.; Reyes, A.P.; Lanner, F. Functional genetics of early human development. Curr. Opin. Genet. Dev. 2018, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Roder, K.; Wolf, S.; Larkin, K.; Schweizer, M. Interaction between the two ubiquitously expressed transcription factors NF-Y and Sp1. Gene 1999, 234, 61–69. [Google Scholar] [CrossRef]
- Yamada, K.; Tanaka, T.; Miyamoto, K.; Noguchi, T. Sp family members and nuclear factor-Y cooperatively stimulate transcription from the rat pyruvate kinase M gene distal promoter region via their direct interactions. J. Biol. Chem. 2000, 275, 18129–18137. [Google Scholar] [CrossRef] [PubMed]
- Eidahl, J.O.; Giesige, C.R.; Domire, J.S.; Wallace, L.M.; Fowler, A.M.; Guckes, S.M.; Garwick-Coppens, S.E.; Labhart, P.; Harper, S.Q. Mouse Dux is myotoxic and shares partial functional homology with its human paralog DUX4. Hum. Mol. Genet. 2016, 25, 4577–4589. [Google Scholar] [CrossRef]
- Yamada, K.; Osawa, H.; Granner, D.K. Identification of proteins that interact with NF-YA. FEBS Lett. 1999, 460, 41–45. [Google Scholar] [CrossRef]
- Oldfield, A.J.; Yang, P.; Conway, A.E.; Cinghu, S.; Freudenberg, J.M.; Yellaboina, S.; Jothi, R. Histone-fold domain protein NF-Y promotes chromatin accessibility for cell type-specific master transcription factors. Mol. Cell 2014, 55, 708–722. [Google Scholar] [CrossRef]
- Dolfini, D.; Minuzzo, M.; Pavesi, G.; Mantovani, R. The short isoform of NF-YA belongs to the embryonic stem cell transcription factor circuitry. Stem Cells 2012, 30, 2450–2459. [Google Scholar] [CrossRef]
- Gao, L.; Wu, K.; Liu, Z.; Yao, X.; Yuan, S.; Tao, W.; Yi, L.; Yu, G.; Hou, Z.; Fan, D.; et al. Chromatin accessibility landscape in human early embryos and its association with evolution. Cell 2018, 173, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Leichsenring, M.; Maes, J.; Mössner, R.; Driever, W.; Onichtchouk, D. Pou5f1 transcription factor controls zygotic gene activation in vertebrates. Science 2013, 341, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Lavagi, I.; Krebs, S.; Simmet, K.; Beck, A.; Zakhartchenko, V.; Wolf, E.; Blum, H. Single-cell RNA sequencing reveals developmental heterogeneity of blastomeres during major genome activation in bovine embryos. Sci. Rep. 2018, 8, 4071. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, N.; Takahashi, K.; Emura, N.; Fujii, T.; Hirayama, H.; Kageyama, S.; Hashizume, T.; Sawai, K. The necessity of OCT-4 and CDX2 for early development and gene expression involved in differentiation of inner cell mass and trophectoderm lineages in bovine embryos. Cell. Reprogram. 2016, 18, 309–318. [Google Scholar] [CrossRef]
- Dumollard, R.; Carroll, J.; Duchen, M.; Campbell, K.; Swann, K. Mitochondrial function and redox state in mammalian embryos. Semin. Cell Dev. Biol. 2009, 20, 346–353. [Google Scholar] [CrossRef]
- Thundathil, J.; Filion, F.; Smith, L.C. Molecular control of mitochondrial function in preimplantation mouse embryos. Mol. Reprod. Dev. 2005, 71, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, R.; Sharpley, M.S.; Chi, F.; Braas, D.; Zhou, Y.; Kim, R.; Clark, A.T.; Banerjee, U. Nuclear localization of mitochondrial TCA cycle enzymes as a critical step in mammalian zygotic genome activation. Cell 2017, 168, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhao, X.; Wu, M.; Yang, L.; Liu, X.; Li, D.; Xu, H.; Zhao, Y.; Su, X.; Wei, Z.; et al. Low Expression of Mitofusin 1 Gene Leads to Mitochondrial Dysfunction and Embryonic Genome Activation Failure in Ovine-Bovine Inter-Species Cloned Embryos. Int. J. Mol. Sci. 2022, 23, 10145. [Google Scholar] [CrossRef]
- Benatti, P.; Belluti, S.; Miotto, B.; Neusiedler, J.; Dolfini, D.; Drac, M.; Basile, V.; Schwob, E.; Mantovani, R.; Blow, J.J.; et al. Direct non transcriptional role of NF-Y in DNA replication. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 673–685. [Google Scholar] [CrossRef]
- Yang, Y.; Deng, Y.; Chen, X.; Zhang, J.; Chen, Y.; Li, H.; Wu, Q.; Yang, Z.; Zhang, L.; Liu, B. Inhibition of PDGFR by CP-673451 induces apoptosis and increases cisplatin cytotoxicity in NSCLC cells via inhibiting the Nrf2-mediated defense mechanism. Toxicol. Lett. 2018, 295, 88–98. [Google Scholar] [CrossRef]
- Darbandi, S.; Darbandi, M.; Akhondi, M.; Sadeghi, M. Studying the place of Iran in the world for medical tourism in infertility fields. Int. J. Reprod. BioMed. 2014, 12, 69. [Google Scholar]
- Idrees, M.; Xu, L.; Song, S.-H.; Joo, M.-D.; Lee, K.-L.; Muhammad, T.; El Sheikh, M.; Sidrat, T.; Kong, I.-K. PTPN11 (SHP2) is indispensable for growth factors and cytokine signal transduction during bovine oocyte maturation and blastocyst development. Cells 2019, 8, 1272. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Idrees, M.; Sidrat, T.; Joo, M.; Xu, L.; Ko, J.; Kong, I.-K. BOEC–Exo addition promotes in vitro maturation of bovine oocyte and enhances the developmental competence of early embryos. Animals 2022, 12, 424. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | No. of Presumed Zygotes | No. of Cleavage Embryo (% ± SEM) | No. of Blastocysts (% ± SEM) | No. of Hatched Blastocysts (% ± SEM) |
| Control | 240 | 190 (79.1 ± 1.32) a | 72 (30.0 ± 0.36) a | 21 (29.1 ± 1.26) a |
| Activator | 240 | 202 (84.2 ± 1.92) b | 93 (38.7 ± 1.25) b | 37 (39.7 ± 1.58) b |
| Inhibitor | 238 | 168 (70.5 ± 1.86) c | 40 (16.8 ± 0.57) c | 8 (20.0 ± 1.38) c |
| Name | Accession No. | Order Name | Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|---|
| AKT | NM_173986.2 | F | CTCGCACGGAGGATCTGTAT | 88 |
| R | TCCTCTCCATCCTGTGTTGG | |||
| ARGFX | XM 024997266.1 | F | GCTAGTGGCCTCAGTTCCTG | 73 |
| R | GGAGGTGGTCACATAACGCA | |||
| CDX2 | NM_001206299.1 | F | TGAGGAGCATGGACTCTGCTA | 82 |
| R | GGGCTAGGTCAGCTGGTAAAC | |||
| DPRX | XM 024979480.1 | F | GCGTCCAGACTTGCACAAAG | 169 |
| R | TGCCAACTGTTTCTCCGTGA | |||
| DUXA | XM 024979492.1 | F | GCCGTACCTCGTTCACAGAA | 70 |
| R | CCAGGGTAAGGGTTTTGGCT | |||
| GSC | NM_001192386.1 | F | GACCAAGTACCCAGACGTGG | 96 |
| R | TCTCAGCGTTTTCCGACTCC | |||
| KLF4 | NM_001105385.1 | F | CCAACTACCCTCCCTTCCTG | 73 |
| R | GGCATGAGCTCTTGGTAATGG | |||
| MAPK1 | NM_175793.2 | F | AACAAAGTCCGAGTCGCCAT | 194 |
| R | CGATGGTCGGTGCTCGAATA | |||
| NANOG | NM_001025344.1 | F | TGTGTCAATTTGAGGGAAGGGT | 175 |
| R | ACTTTTGCCCCCTGTGCTTA | |||
| NFYA | NM_001014956.1 | F | GATTTGGAGGGGCCATGGAA | 197 |
| R | CATTAATGGCTGCCCCTGGA | |||
| NFYB | NM_001079786.2 | F | TGTCCAACCAAACAGCCGAT | 124 |
| R | AGAACCGTGTTGTCAGTGGT | |||
| NFYC | NM_001034598.1 | F | CAGAGGTCCAGCAAGGACAG | 117 |
| R | ACTGGATGAACATGGGCTGG | |||
| OCT4 | NM_174580.3 | F | GGGCAAACGATCAAGCAGTG | 172 |
| R | CTCAGGGAATGGGACCGAAG | |||
| PDGFRα | NM_001075896.2 | F | CGCGGTTTTTGAGCCCATTA | 95 |
| R | GGAAAAAGTGTGTCCACGGC | |||
| PDGFRβ | NM_001075896.2 | F | AAGGCATCAGCAGCAAGGAT | 169 |
| R | GTGCTGGAGAGGTTGAGGAC | |||
| SP1 | NM_001078027.1 | F | TGCTACCATGAGCGACCAAG | 190 |
| R | CAAAGGGGATGGCTGGGATT | |||
| SRF | NM_001206016.1 | F | AGGCACTGATTCAGACCTGC | 98 |
| R | TGTCTCTTCAAAGCCGGTGG | |||
| SOX2 | NM_001105463.2 | F | GGCGGGGGAGACATTTTCAA | 78 |
| R | AGCGTACCGGGTTTTTCTCC | |||
| SALL4 | NM_001192836.3 | F | CGTCCAAGAAAGGCAAAGGG | 127 |
| R | TGCAAGGAGCTATCAGTCCC | |||
| ZHX1 | NM_001205310.2 | F | GTTTGAAATGCAAGATGGCGG | 145 |
| R | TTTGACACGGAAGGGTGTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perera, C.D.; Idrees, M.; Khan, A.M.; Haider, Z.; Ullah, S.; Kang, J.-S.; Lee, S.-H.; Kang, S.-M.; Kong, I.-K. PDGFRβ Activation Induced the Bovine Embryonic Genome Activation via Enhanced NFYA Nuclear Localization. Int. J. Mol. Sci. 2023, 24, 17047. https://doi.org/10.3390/ijms242317047
Perera CD, Idrees M, Khan AM, Haider Z, Ullah S, Kang J-S, Lee S-H, Kang S-M, Kong I-K. PDGFRβ Activation Induced the Bovine Embryonic Genome Activation via Enhanced NFYA Nuclear Localization. International Journal of Molecular Sciences. 2023; 24(23):17047. https://doi.org/10.3390/ijms242317047
Chicago/Turabian StylePerera, Chalani Dilshani, Muhammad Idrees, Abdul Majid Khan, Zaheer Haider, Safeer Ullah, Ji-Su Kang, Seo-Hyun Lee, Seon-Min Kang, and Il-Keun Kong. 2023. "PDGFRβ Activation Induced the Bovine Embryonic Genome Activation via Enhanced NFYA Nuclear Localization" International Journal of Molecular Sciences 24, no. 23: 17047. https://doi.org/10.3390/ijms242317047
APA StylePerera, C. D., Idrees, M., Khan, A. M., Haider, Z., Ullah, S., Kang, J.-S., Lee, S.-H., Kang, S.-M., & Kong, I.-K. (2023). PDGFRβ Activation Induced the Bovine Embryonic Genome Activation via Enhanced NFYA Nuclear Localization. International Journal of Molecular Sciences, 24(23), 17047. https://doi.org/10.3390/ijms242317047