
Analysis of Chromatin Accessibility Changes Induced by BMMC Recognition of Foot-and-Mouth Disease Virus-like Particles through ATAC-seq


Abstract
:1. Introduction
2. Results
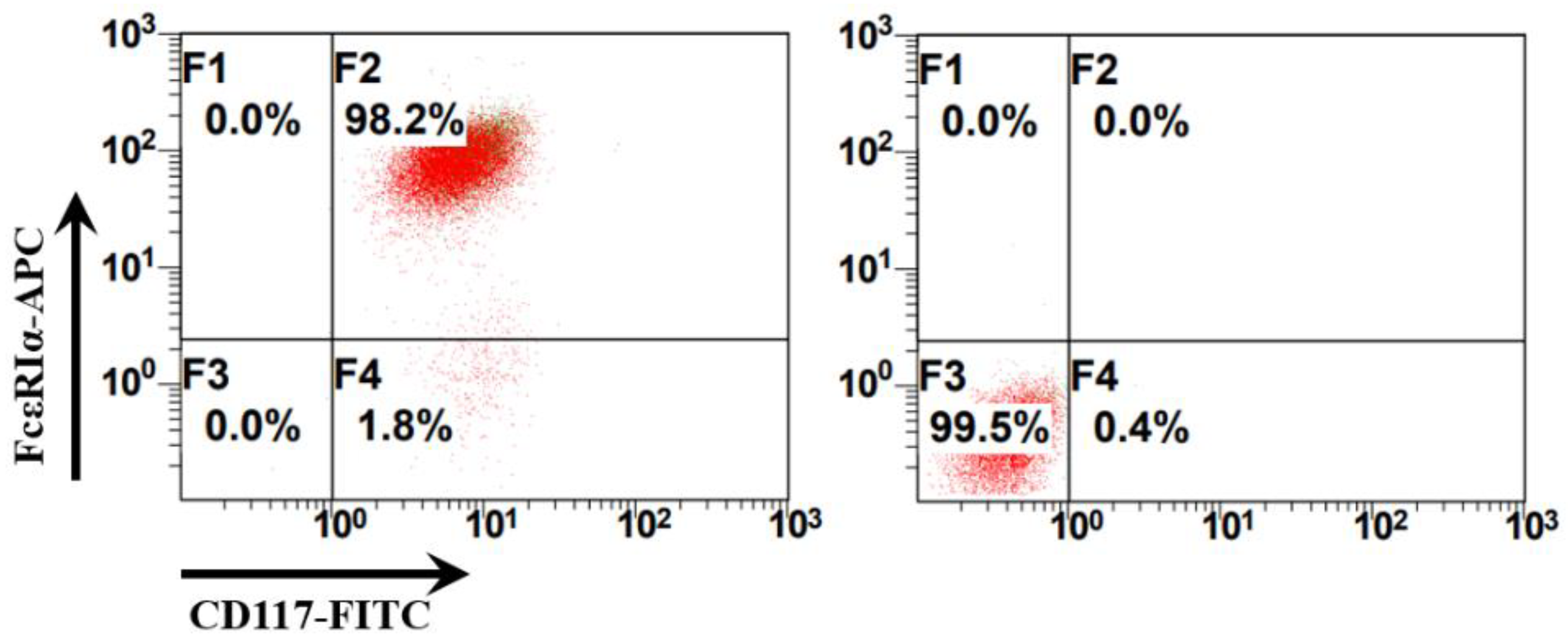
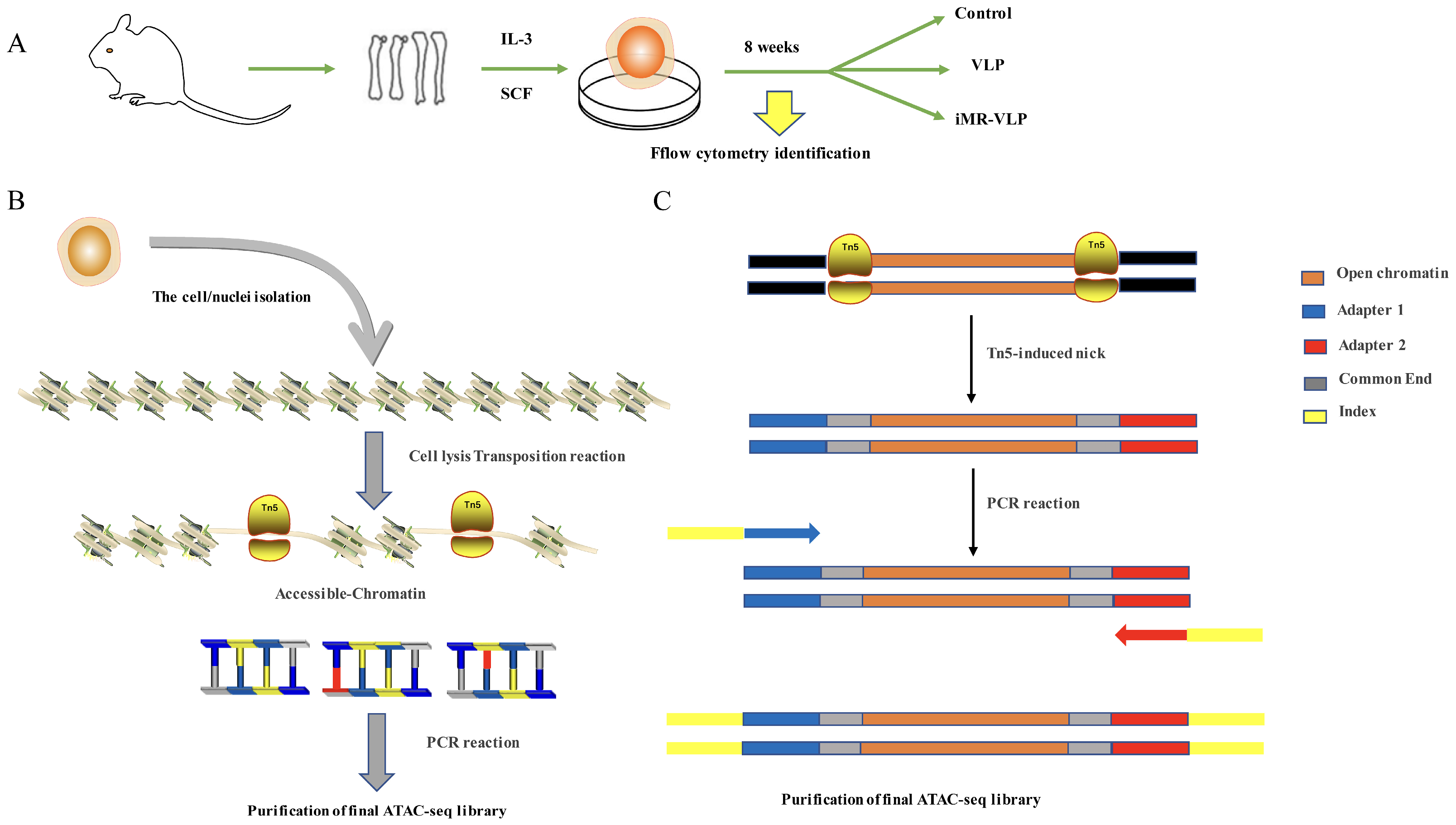
2.1. Bone Marrow-Derived Mast Cells Preparation
2.2. ATAC-seq Data Quality Control
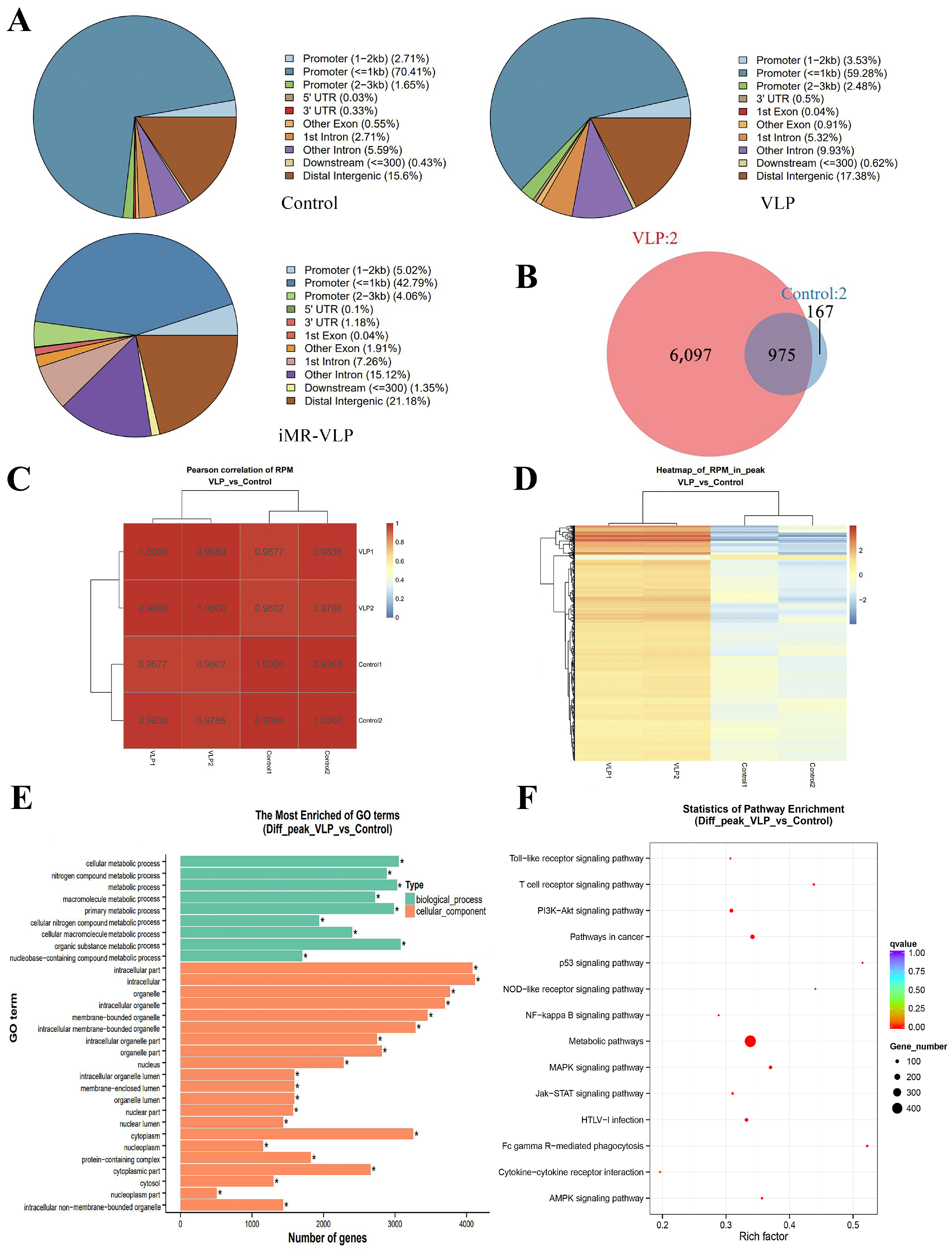
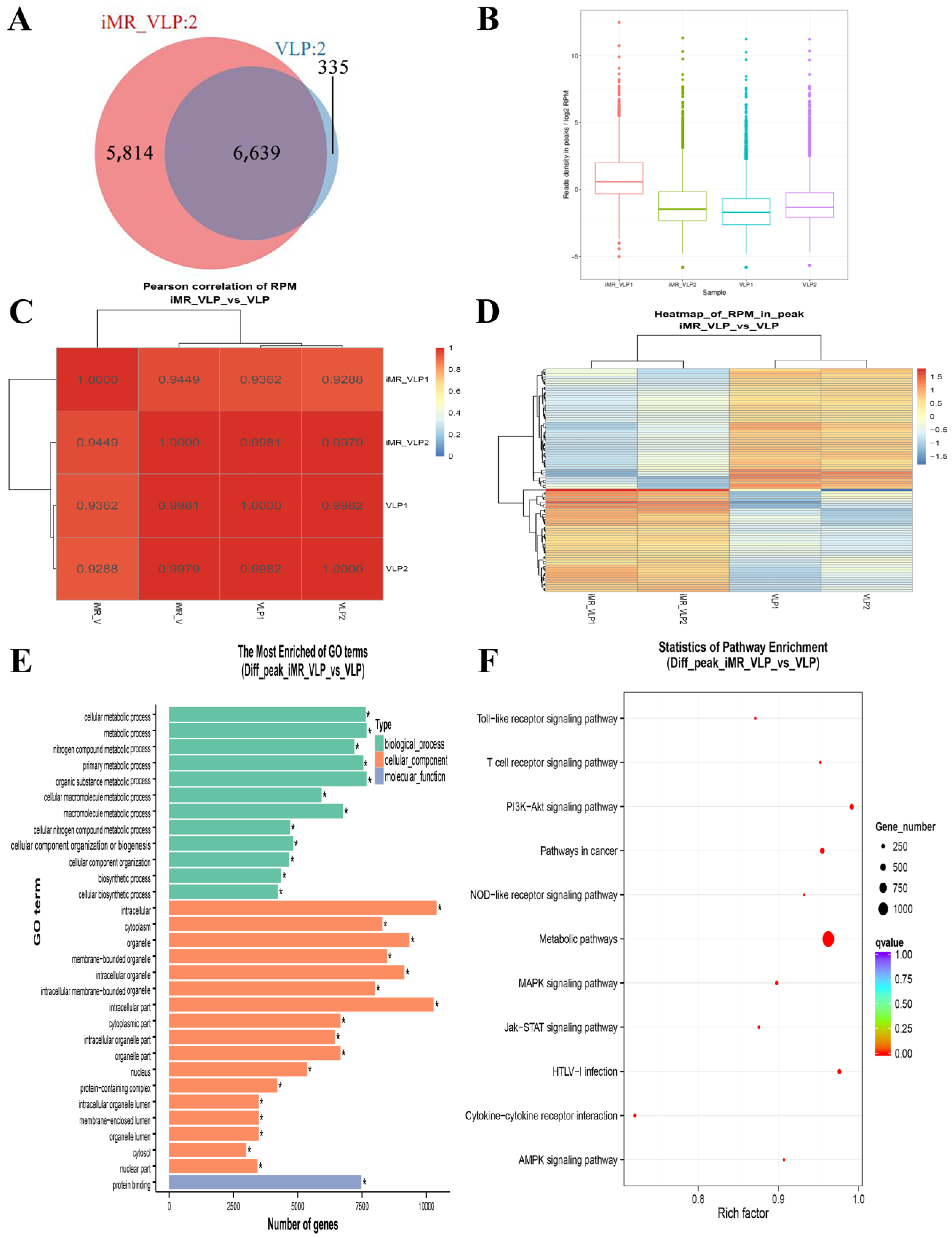
2.3. Chromatin Accessibility Profiles in BMMCs upon Their Recognition of VLPs
2.4. Motif Enrichment Analysis
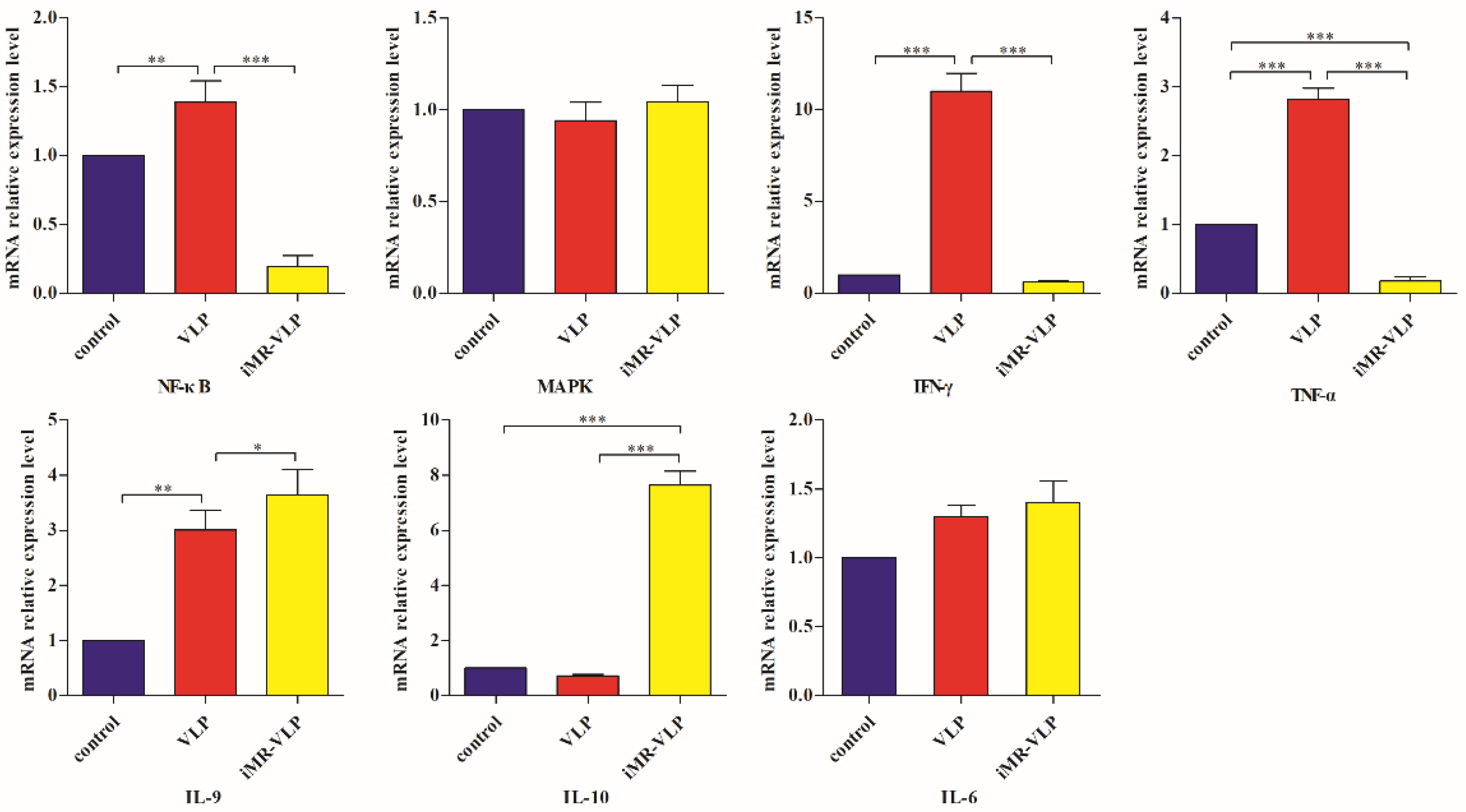
2.5. Validation of the Expression Levels of Critical Cytokine Genes Using Quantitative Reverse Transcription Polymerase Chain Reaction Tests
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cell Samples Preparation
4.3. Library Construction and Sequencing
4.4. Validation of the Expression Levels of Critical Cytokine Genes Using RT-qPCR Tests
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Los Santos, T.; Diaz-San Segundo, F.; Rodriguez, L.L. The Need for Improved Vaccines against Foot-and-Mouth Disease. Curr. Opin. Virol. 2018, 29, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Knight-Jones, T.J.; Rushton, J. The Economic Impacts of Foot and Mouth Disease-What are They, How Big are They and Where do They Occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef]
- Li, K.; Wang, C.; Yang, F.; Cao, W.; Zhu, Z.; Zheng, H. Virus-Host Interactions in Foot-and-Mouth Disease Virus Infection. Front. Immunol. 2021, 12, 571509. [Google Scholar] [CrossRef]
- Robinson, L.; Knight-Jones, T.J.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 3-Vaccines. Transbound. Emerg. Dis. 2016, 1, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.M.; Cai, J.C.; Chen, H.T.; Chen, L. Protection of a Novel Epitope-RNA VLP Double-Effective VLP Vaccine for Foot-and-Mouth Disease. Antivir. Res. 2016, 134, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Bidart, J.; Mignaqui, A.; Kornuta, C.; Lupi, G.; Gammella, M.; Soria, I.; Galarza, R.; Ferella, A.; Cardillo, S.; Langellotti, C.; et al. FMD Empty Capsids Combined with the Immunostant Particle Adjuvant -ISPA or ISA206 Induce Protective Immunity against Foot and Mouth Disease Virus. Virus Res. 2021, 297, 198339. [Google Scholar] [CrossRef]
- Primavera, V.; Simmons, J.; Clark, B.A.; Neilan, J.G.; Puckette, M. Effect of Foot-and-Mouth Disease Virus 2B Viroporin on Expression and Extraction of Mammalian Cell Culture Produced Foot-and-Mouth Disease Virus-like Particles. Vaccines 2022, 10, 1506. [Google Scholar] [CrossRef]
- Shi, X.; Yang, K.; Song, H.; Teng, Z.; Zhang, Y.; Ding, W.; Wang, A.; Tan, S.; Dong, H.; Sun, S.; et al. Development and Efficacy Evaluation of a Novel Nano-Emulsion Adjuvant for a Foot-and-Mouth Disease Virus-like Particles Vaccine Based on Squalane. Nanomaterials 2022, 12, 3934. [Google Scholar] [CrossRef]
- Arzt, J.; Pacheco, J.M.; Rodriguez, L.L. The Early Pathogenesis of Foot-and-Mouth Disease in Cattle after Aerosol Inoculation. Identification of the Nasopharynx as the Primary Site of Infection. Vet. Pathol. 2010, 47, 1048–1063. [Google Scholar] [CrossRef]
- Stenfeldt, C.; Pacheco, J.M.; Rodriguez, L.L.; Arzt, J. Infection Dynamics of Foot-and-Mouth Disease Virus in Pigs using Two Novel Simulated-Natural Inoculation Methods. Res. Vet. Sci. 2014, 96, 396–405. [Google Scholar] [CrossRef]
- Jiménez, M.; Cervantes-García, D.; Córdova-Dávalos, L.E.; Pérez-Rodríguez, M.J.; Gonzalez-Espinosa, C.; Salinas, E. Responses of Mast Cells to Pathogens: Beneficial and Detrimental Roles. Front. Immunol. 2021, 12, 685865. [Google Scholar] [CrossRef] [PubMed]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef] [PubMed]
- Katsoulis-Dimitriou, K.; Kotrba, J.; Voss, M.; Dudeck, J.; Dudeck, A. Mast Cell Functions Linking Innate Sensing to Adaptive Immunity. Cells 2020, 9, 2538. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Weaver, B.; Choi, H.W.; Abraham, S.N.; Staats, H.F. Mast cell activators as novel immune regulators. Curr. Opin. Pharmacol. 2018, 41, 89–95. [Google Scholar] [CrossRef]
- Han, W.J.; Zhang, J.J.; Zhang, Y.M.; Wang, J.X.; Li, L.M. Cytokine Responses of Bone Marrow-derived Mast Cells to FMDV-VLPs via Mannose Receptors. Acta Vet. Zootech. Sin. 2022, 53, 3917–3926. [Google Scholar] [CrossRef]
- Zhang, J.; Han, W.; Li, M.; Bai, R.; Tian, Z.; Yuan, W.; Li, L. Histone Acetylation Regulates BMMCs Recognition of Foot-and-Mouth Disease Virus-like Particles. Int. Immunopharmacol. 2023, 121, 110428. [Google Scholar] [CrossRef]
- Cavalli, G.; Heard, E. Advances in Epigenetics Link Genetics to the Environment and Disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef]
- Luo, L.; Gribskov, M.; Wang, S. Bibliometric Review of ATAC-Seq and Its Application in Gene Expression. Brief. Bioinform. 2022, 23, bbac061. [Google Scholar] [CrossRef]
- Maston, G.A.; Evans, S.K.; Green, M.R. Transcriptional Regulatory Elements in the Human Genome. Annu. Rev. Genom. Hum. Genet. 2006, 7, 29–59. [Google Scholar] [CrossRef]
- Grandi, F.C.; Modi, H.; Kampman, L.; Corces, M.R. Chromatin Accessibility Profiling by ATAC-seq. Nat. Protoc. 2022, 17, 1518–1552. [Google Scholar] [CrossRef]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of Native Chromatin for Fast and Sensitive Epigenomic Profiling of Open Chromatin, DNA-binding Proteins and Nucleosome Position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef]
- Scott-Browne, J.P.; López-Moyado, I.F.; Trifari, S.; Wong, V.; Chavez, L.; Rao, A.; Pereira, R.M. Dynamic Changes in Chromatin Accessibility Occur in CD8+ T Cells Responding to Viral Infection. Immunity 2016, 45, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Hofmeister, B.T.; Vollmers, C.; DuBois, R.M.; Schmitz, R.J. Combining ATAC-seq with Nuclei Sorting for Discovery of Cis-regulatory Regions in Plant Genomes. Nucleic Acids Res. 2017, 45, e41. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Huang, Y.; Liu, X.; Sia, K.C.S.; Zhang, J.C.; Tai, X.; Wang, M.; Toscan, C.E.; McCalmont, H.; Evans, K.; et al. Lymphocyte-Specific Chromatin Accessibility Pre-determines Glucocorticoid Resistance in Acute Lymphoblastic Leukemia. Cancer 2018, 34, 906–921.e8. [Google Scholar] [CrossRef] [PubMed]
- de León, P.; Cañas-Arranz, R.; Defaus, S.; Torres, E.; Forner, M.; Bustos, M.J.; Revilla, C.; Dominguez, J.; Andreu, D.; Blanco, E.; et al. Swine T-Cells and Specific Antibodies Evoked by Peptide Dendrimers Displaying Different FMDV T-Cell Epitopes. Front. Immunol. 2021, 11, 621537. [Google Scholar] [CrossRef]
- Roh, J.H.; Bui, N.A.; Lee, H.S.; Bui, V.N.; Dao, D.T.; Vu, T.T.; Hoang, T.T.; So, K.M.; Yi, S.W.; Kim, E.; et al. Age-dependent Immune Response in Pigs against Foot-and-mouth Disease Virus in Vitro. J. Anim. Sci. Technol. 2021, 63, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Díaz-San Segundo, F.; Rodríguez-Calvo, T.; de Avila, A.; Sevilla, N. Immunosuppression during Acute Infection with Foot-and-mouth Disease Virus in Swine is Mediated by IL-10. PLoS ONE 2009, 4, e5659. [Google Scholar] [CrossRef]
- Moretti, S.; Renga, G.; Oikonomou, V.; Galosi, C.; Pariano, M.; Iannitti, R.G.; Borghi, M.; Puccetti, M.; De Zuani, M.; Pucillo, C.E.; et al. A Mast cell-ILC2-Th9 Pathway Promotes Lung Inflammation in Cystic Fibrosis. Nat. Commun. 2017, 8, 14017. [Google Scholar] [CrossRef]
- Yin, J.J.; Hu, X.Q.; Mao, Z.F.; Bao, J.; Qiu, W.; Lu, Z.Q.; Wu, H.T.; Zhong, X.N. Neutralization of Interleukin-9 Decreasing Mast Cells Infiltration in Experimental Autoimmune Encephalomyelitis. Chin. Med. J. 2017, 130, 964–971. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef]
- Miyazawa, M.; Noguchi, K.; Kujirai, M.; Katayama, K.; Yamagoe, S.; Sugimoto, Y. IL-10 Promoter Transactivation by the Viral K-RTA Protein Involves the Host-cell Transcription Factors, Specificity Proteins 1 and 3. J. Biol. Chem. 2018, 293, 662–676. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Liang, J.; Liu, B.; Li, Y.; Gao, J.; Keating, A.; Mohamed, F.; Dai, S.; Reinhardt, R.; Jiong, Y.; et al. The Clusters of Transcription Factors NFATC2, STAT5, GATA2, AP1, RUNX1 and EGR2 Binding Sites at the Induced Il13 Enhancers Mediate Il13 Gene Transcription in Response to Antigenic Stimulation. J. Immunol. 2020, 205, 3311–3318. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, M.; Wei, X.; Wu, L.; Xu, J.; Dai, X.; Xia, J.; Cheng, M.; Yuan, Y.; Zhang, P.; et al. An ATAC-seq Atlas of Chromatin Accessibility in Mouse Tissues. Sci. Data 2019, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- GB/T35892-2018; Laboratory Animal—Guideline for Ethical Review of Animal Welfare. People’s Republic of China General Administration of Quality Supervision, Inspection and Quarantine, China National Standardization Administration: Beijing, China, 2018.
- Abbring, S.; Blokhuis, B.R.J.; Miltenburg, J.L.; Olmedo, K.G.J.R.; Garssen, J.; Redegeld, F.A.; van Esch, B.C.A.M. Direct Inhibition of the Allergic Effector Response by Raw Cow’s Milk-An Extensive In Vitro Assessment. Cells 2020, 9, 1258. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean_Reads | Mapped_MT | Mapped_non_MT | Unique_Mapped | Unique_Mapped_Dedup |
|---|---|---|---|---|---|
| Control1 | 50,330,055 | 742,506 (1.48%) | 46,268,347 (91.93%) | 41,579,440 (82.61%) | 32,311,000 (64.20%) |
| Control2 | 49,614,421 | 514,427 (1.04%) | 46,851,917 (94.43%) | 42,269,831 (85.20%) | 32,833,186 (66.18%) |
| iMR_VLP1 | 58,585,184 | 6,230,516 (10.63%) | 47,602,533 (81.25%) | 42,405,455 (72.38%) | 31,648,544 (54.02%) |
| iMR_VLP2 | 50,138,881 | 2,279,086 (4.55%) | 40,804,218 (81.38%) | 35,892,428 (71.59%) | 27,581,651 (55.01%) |
| VLP1 | 51,866,423 | 1,759,382 (3.39%) | 40,892,763 (78.84%) | 36,130,195 (69.66%) | 27,677,497 (53.36%) |
| VLP2 | 50,104,549 | 1,496,313 (2.99%) | 37,312,069 (74.47%) | 32,930,422 (65.72%) | 25,204,220 (50.30%) |
| Motifs | Factors | p-Value | %Targets | %Background |
|---|---|---|---|---|
 | GABPA(ETS) | 10−623 | 33.64% | 11.44% |
 | SP1 | 10−581 | 32.68% | 11.39% |
 | NRF1 | 10−362 | 14.56% | 3.64% |
 | RUNX(Runt) | 10−319 | 28.24% | 12.53% |
 | AP-1 | 10−304 | 17.35% | 5.74% |
 | GATA3(Zf) | 10−216 | 19.12% | 8.22% |
 | CREB1 | 10−212 | 21.38% | 9.85% |
 | NFY(CCAAT) | 10−175 | 14.46% | 5.92% |
 | GFY | 10−159 | 3.87% | 0.54% |
 | USF2 | 10−91 | 8.65% | 3.73% |
 | RARg(NR) | 10−83 | 0.84% | 0.02% |
 | YY1(Zf) | 10−82 | 4.53% | 1.41% |
 | Mitf | 10−70 | 16.30% | 9.99% |
 | ZBTB33 | 10−55 | 3.40% | 1.14% |
 | RUNX1 | 10−42 | 0.42% | 0.01% |
| Motifs | Factors | p-Value | %Targets | %Background |
|---|---|---|---|---|
 | CRX | 10−84 | 10.03% | 0.02% |
 | RUNX1 | 10−56 | 10.57% | 0.16% |
 | Esrra | 10−31 | 12.74% | 1.24% |
 | PB0166.1_Sox12_2 | 10−30 | 25.47% | 6.47% |
 | SD0002.1_at_AC_acceptor | 10−29 | 11.65% | 1.07% |
 | PB0135.1_Hoxa3_2 | 10−27 | 5.96% | 0.15% |
 | ARE(NR) | 10−25 | 12.20% | 1.50% |
 | Gfi1b | 10−25 | 11.65% | 1.35% |
 | NF1:FOXA1(CTF,Forkhead) | 10−24 | 15.45% | 2.84% |
 | Nkx2-5(var.2) | 10−23 | 3.52% | 0.03% |
 | Slug(Zf) | 10−22 | 12.20% | 1.80% |
 | Pit1 + 1 bp(Homeobox) | 10−21 | 2.98% | 0.02% |
 | NFκB-p65-Rel(RHD) | 10−20 | 4.61% | 0.12% |
 | PB0126.1_Gata5_2 | 10−20 | 10.84% | 1.57% |
 | PB0180.1_Sp4_2 | 10−20 | 7.32% | 0.60% |
| Name | Sequence (5′-3′) | Tm (°C) | Product Size (bp) |
|---|---|---|---|
| NF-κB | Forward: CATCCTCGTCCGCCTATTAC | 50 | 93 |
| Reverse: GACTCTCCTCCCTTTCCTTGT | |||
| MAPK | Forward: AACTATTTGCTTTCTCTCCC | 50 | 133 |
| Reverse: TGTTCAACTTCAATCCTCTT | |||
| IFN-γ | Forward: AGACTCTCATTGCGGGGTTGTATC | 54 | 160 |
| Reverse: ACAGTGTAGACATCTCCTCCCATCAG | |||
| TNF-α | Forward: AAAGGGAGAGTGGTCAGGTTGC | 51 | 95 |
| Reverse: CTCAGGGAAGAATCTGGAAAGGT | |||
| IL-9 | Forward: CAATGCCACACAGAAATCAA | 55 | 99 |
| Reverse: CAGGCAGGAAAAGGACGCTT | |||
| IL-10 | Forward: CGCCCTATTTAGAAAGAAGCCCA | 52 | 140 |
| Reverse: AAAGGAAGAACCCCTCCCATCAT | |||
| GADPH | Forward: CCTTCCGTGTTCCTAC | 45 | 152 |
| Reverse: GACAACCTGGTCCTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, W.; Zhang, J.; Li, M.; An, M.; Li, L. Analysis of Chromatin Accessibility Changes Induced by BMMC Recognition of Foot-and-Mouth Disease Virus-like Particles through ATAC-seq. Int. J. Mol. Sci. 2023, 24, 17044. https://doi.org/10.3390/ijms242317044
Han W, Zhang J, Li M, An M, Li L. Analysis of Chromatin Accessibility Changes Induced by BMMC Recognition of Foot-and-Mouth Disease Virus-like Particles through ATAC-seq. International Journal of Molecular Sciences. 2023; 24(23):17044. https://doi.org/10.3390/ijms242317044
Chicago/Turabian StyleHan, Weijian, Junjuan Zhang, Mingzhu Li, Manxin An, and Limin Li. 2023. "Analysis of Chromatin Accessibility Changes Induced by BMMC Recognition of Foot-and-Mouth Disease Virus-like Particles through ATAC-seq" International Journal of Molecular Sciences 24, no. 23: 17044. https://doi.org/10.3390/ijms242317044
APA StyleHan, W., Zhang, J., Li, M., An, M., & Li, L. (2023). Analysis of Chromatin Accessibility Changes Induced by BMMC Recognition of Foot-and-Mouth Disease Virus-like Particles through ATAC-seq. International Journal of Molecular Sciences, 24(23), 17044. https://doi.org/10.3390/ijms242317044