
Yellow-Green Leaf 19 Encoding a Specific and Conservative Protein for Photosynthetic Organisms Affects Tetrapyrrole Biosynthesis, Photosynthesis, and Reactive Oxygen Species Metabolism in Rice

 ,
, 
Abstract
:1. Introduction
2. Results
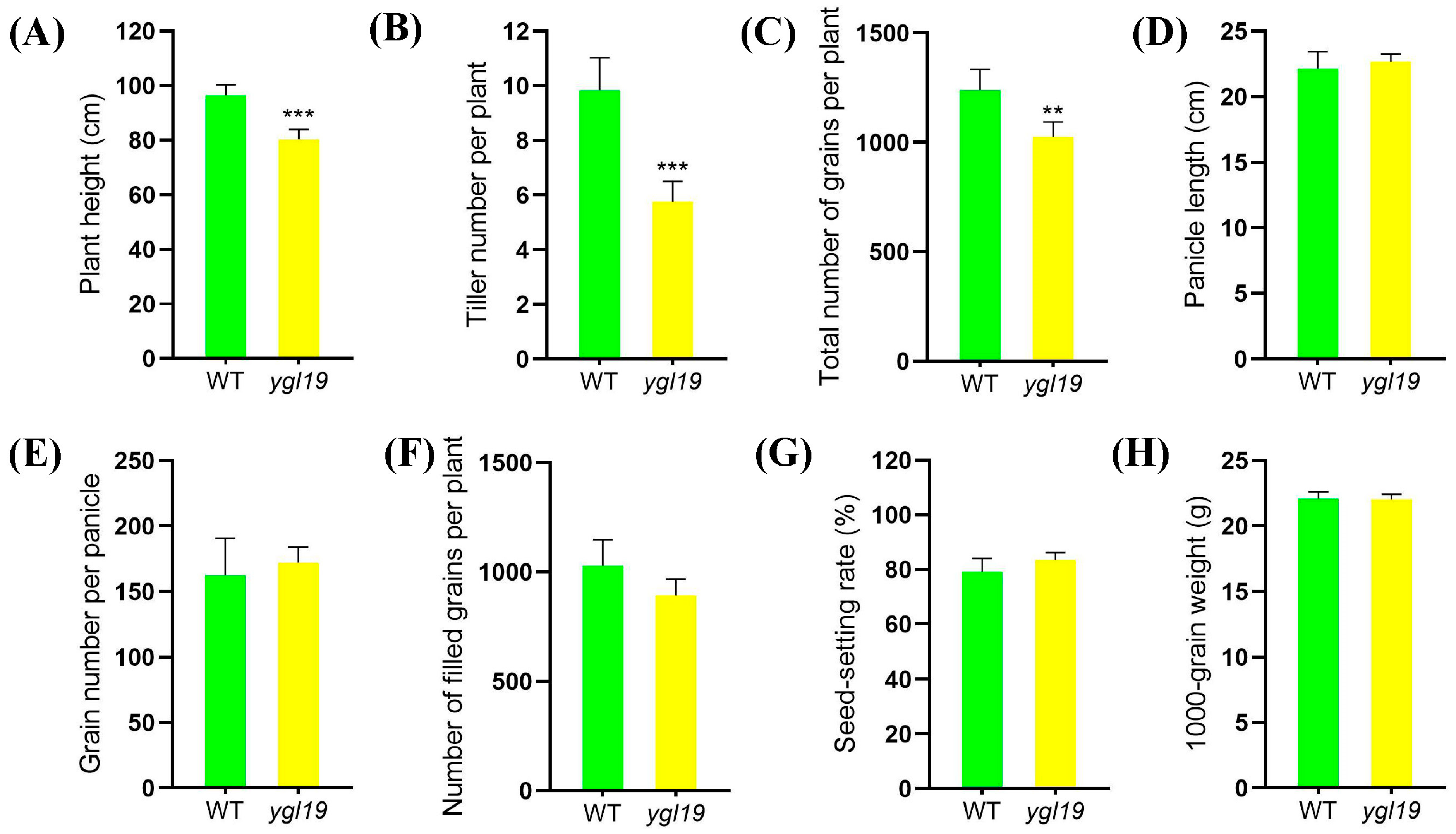
2.1. Phenotypic Characterization of the ygl19 Mutant
2.2. Map-Based Cloning of the YGL19 Gene
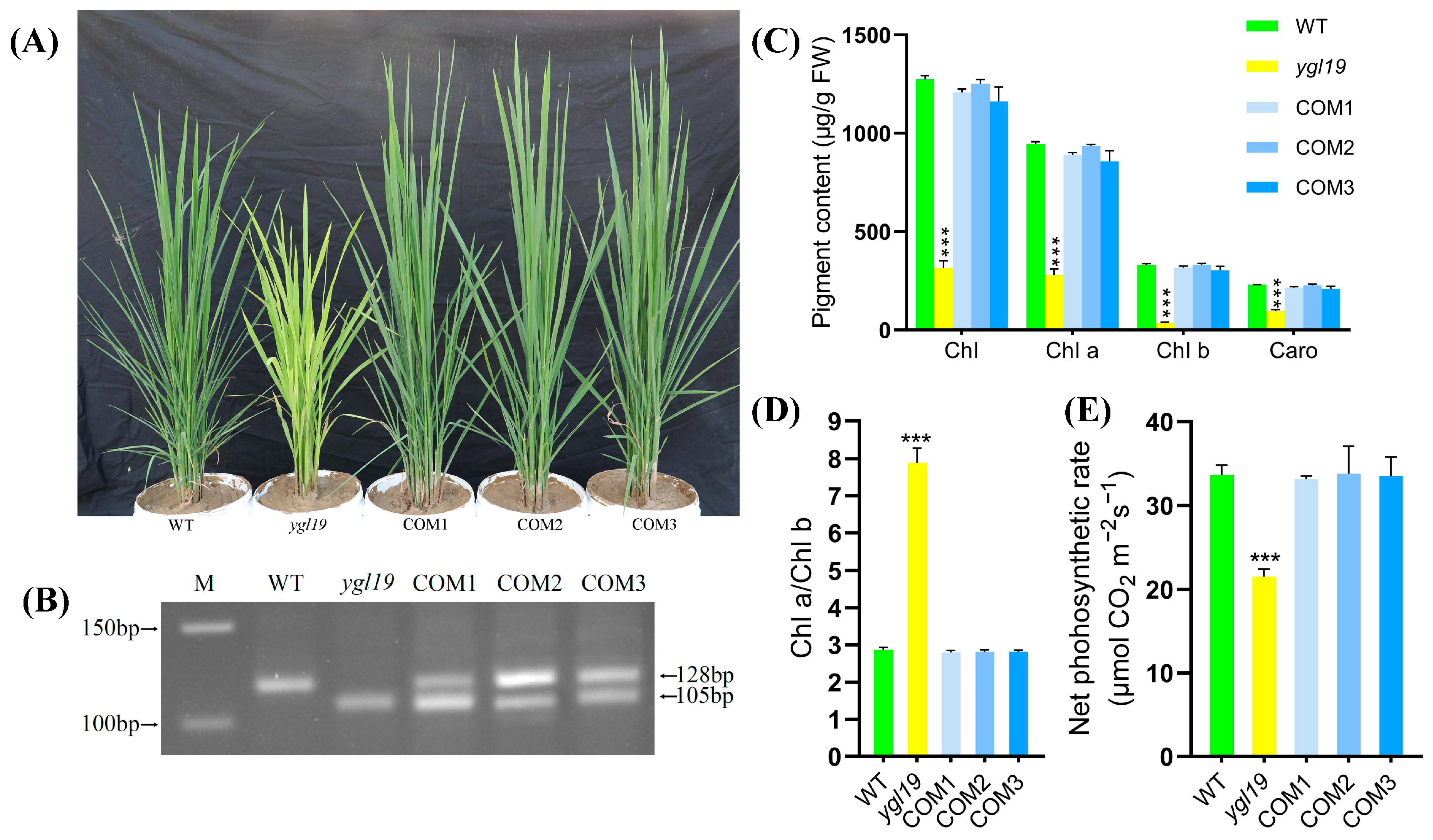
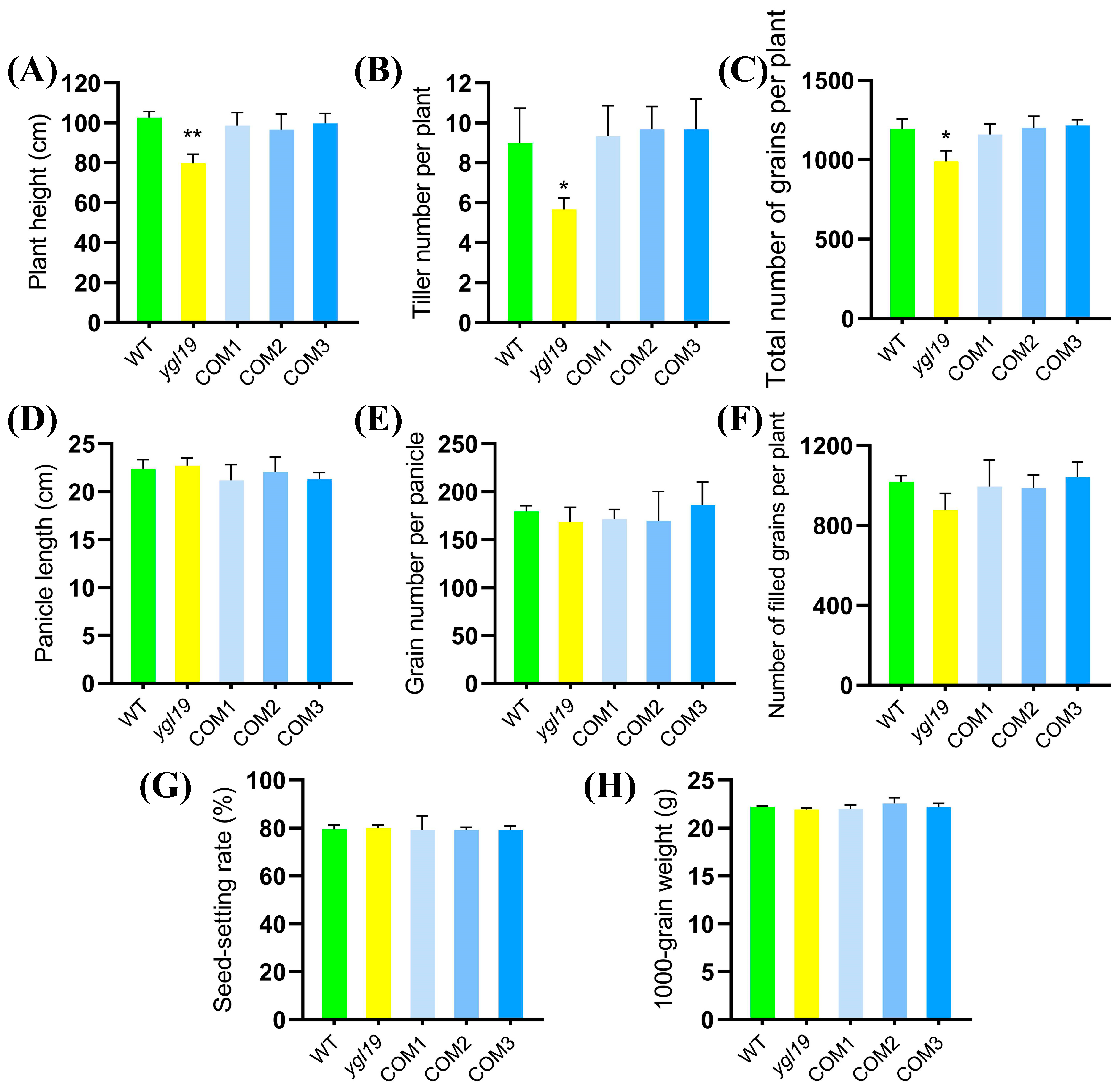
2.3. A Complementation Experiment of LOC_Os03g21370 into the ygl19 Mutant
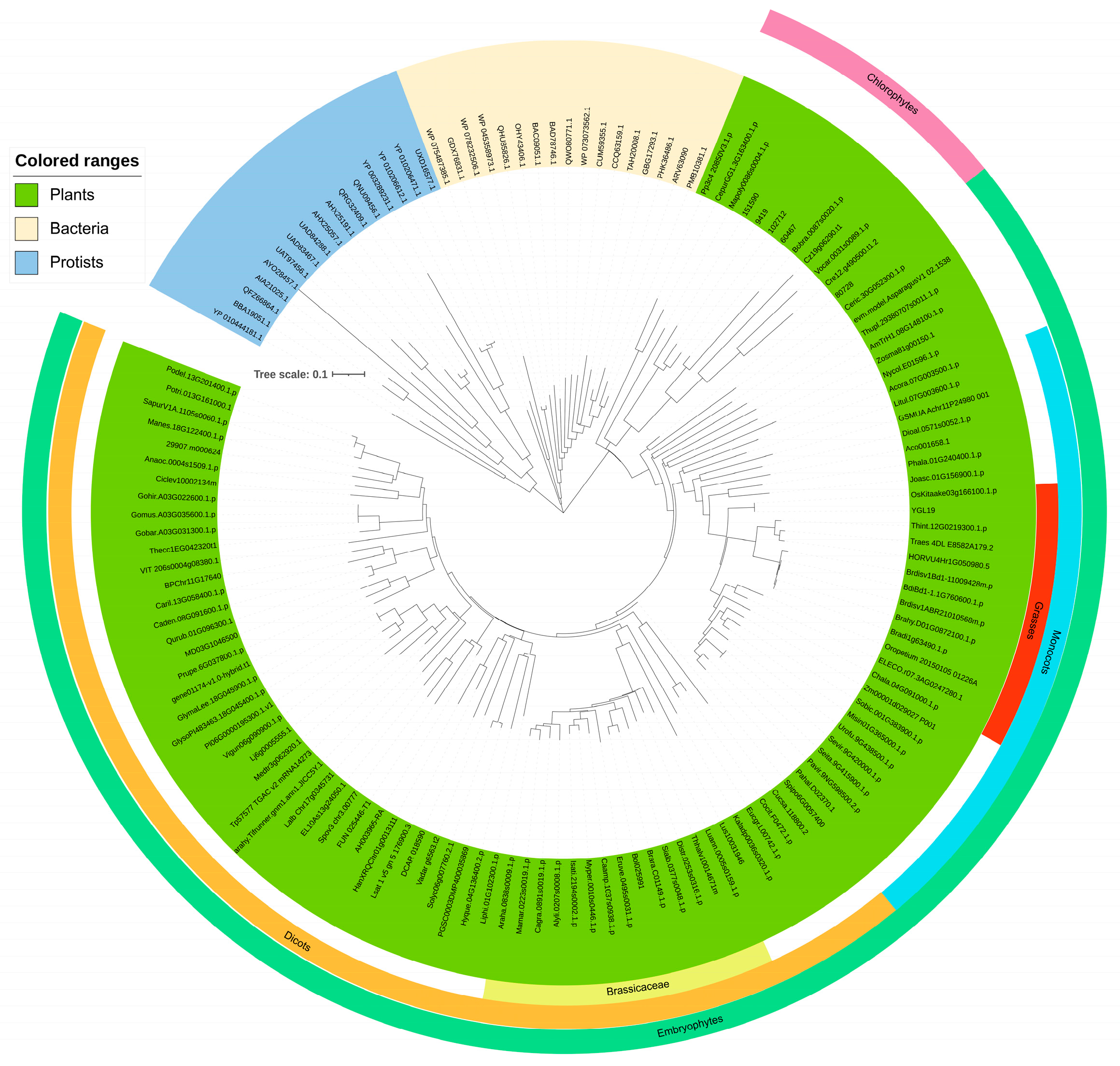
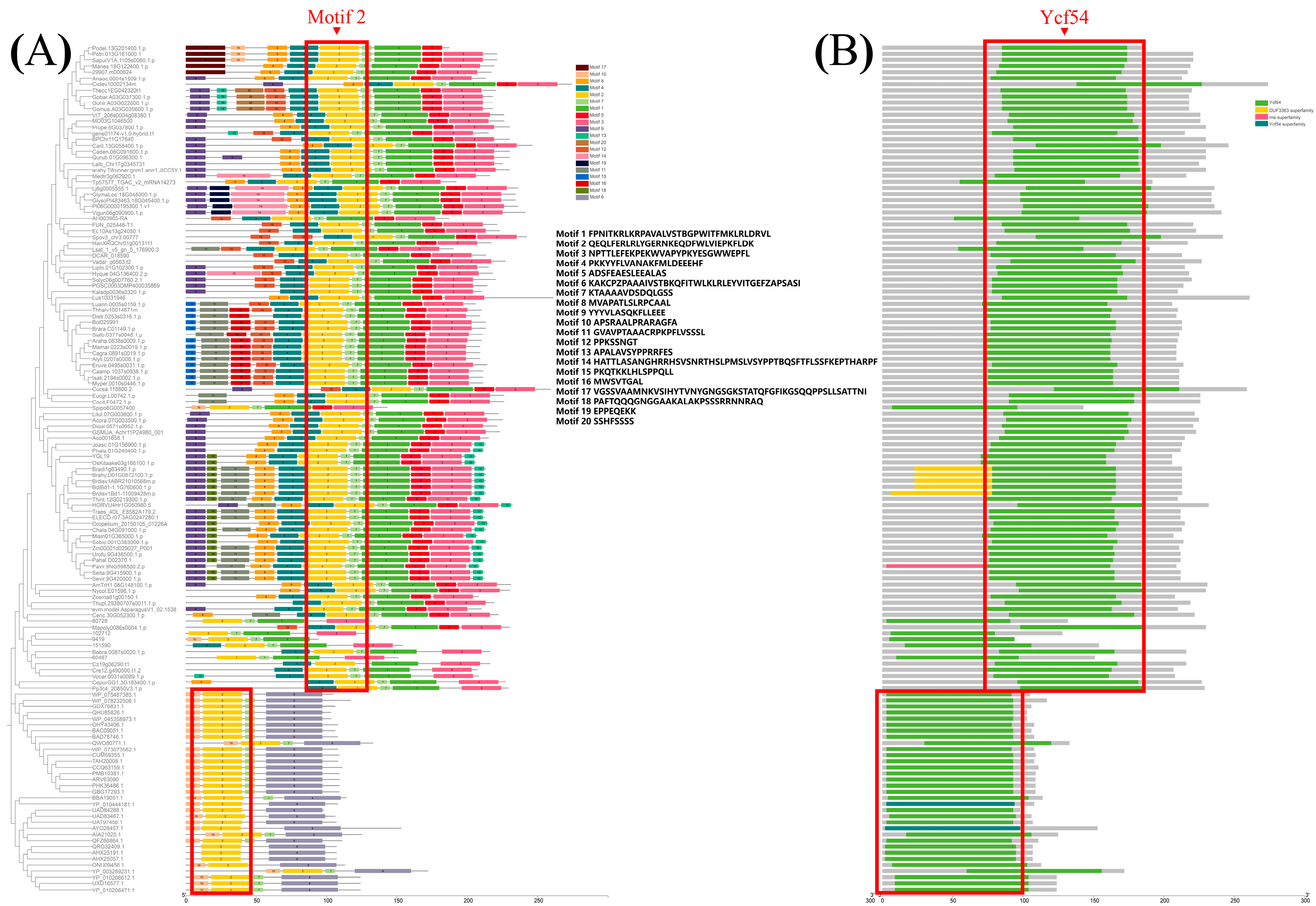
2.4. The YGL19 Protein Homologs Are Unique for Photosynthetic Organisms and Are Evolutionarily Conserved, Containing a Ycf54 Functional Domain
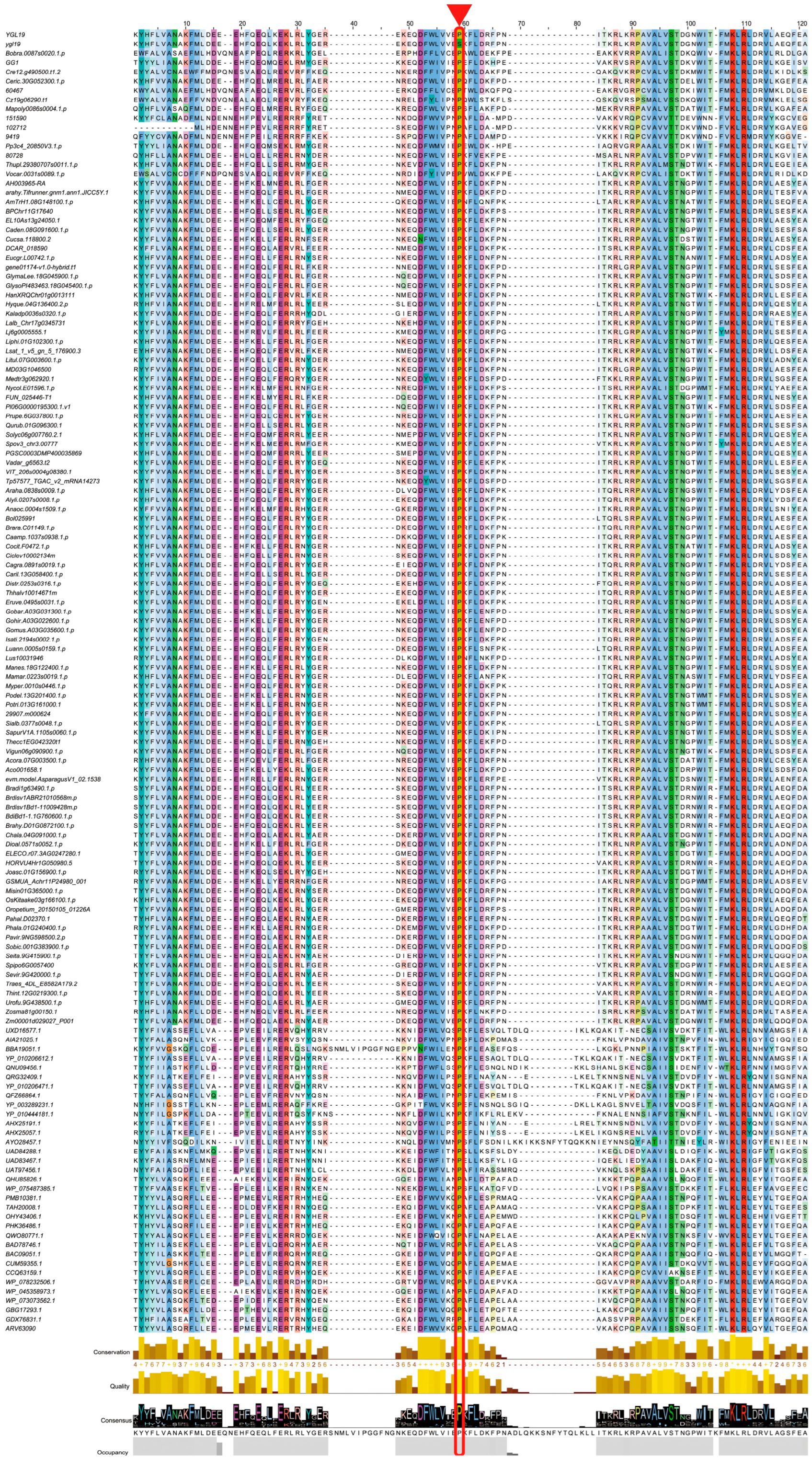
2.5. A Conserved Amino Acid on the Ycf54 Domain Is Substituted to Form the ygl19 Protein
2.6. The YGL19 Gene Is Conserved in a Natural Rice Variant
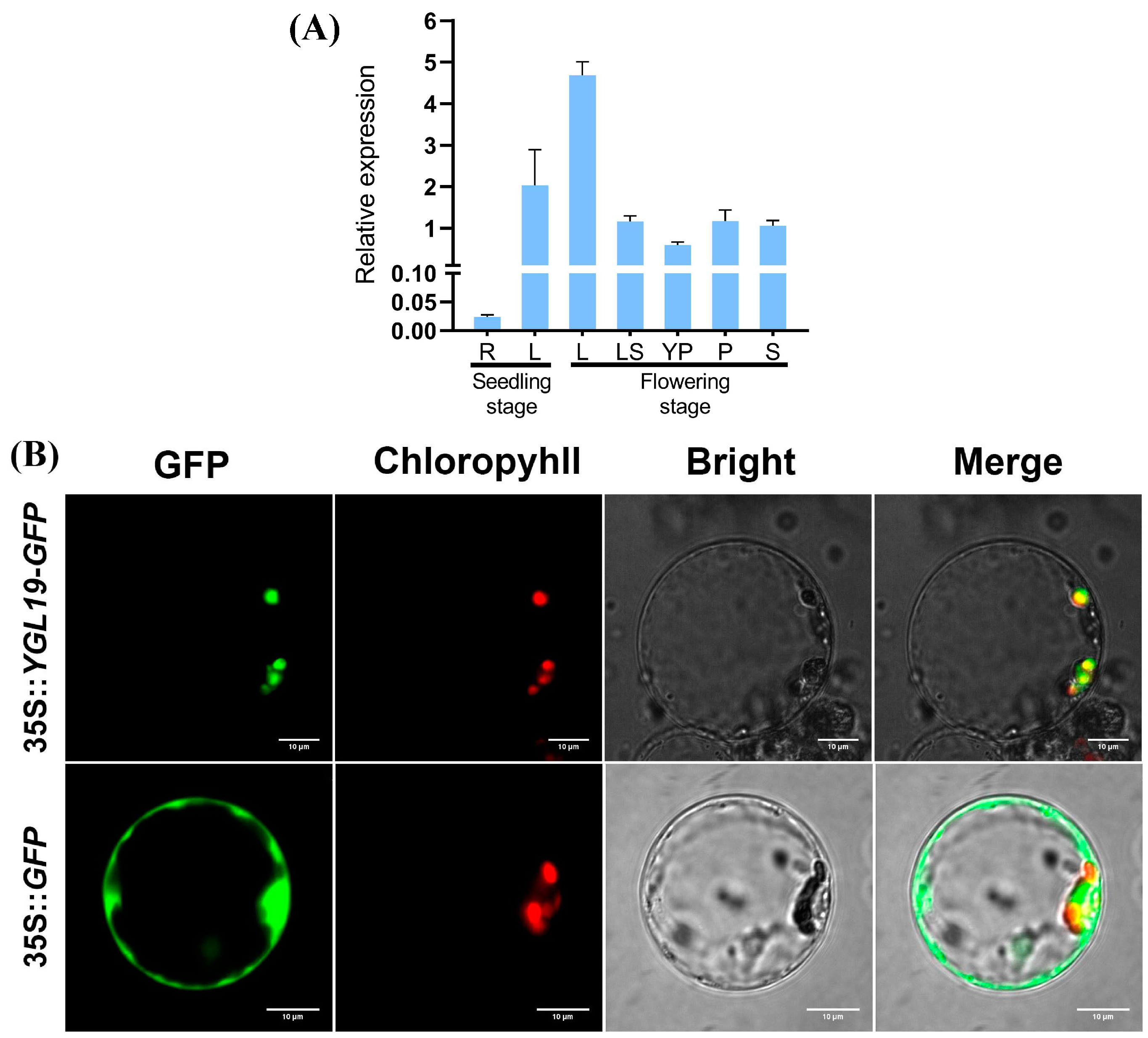
2.7. The YGL19 Gene Is Mainly Expressed in Green Tissues and Localized in Chloroplasts for Function
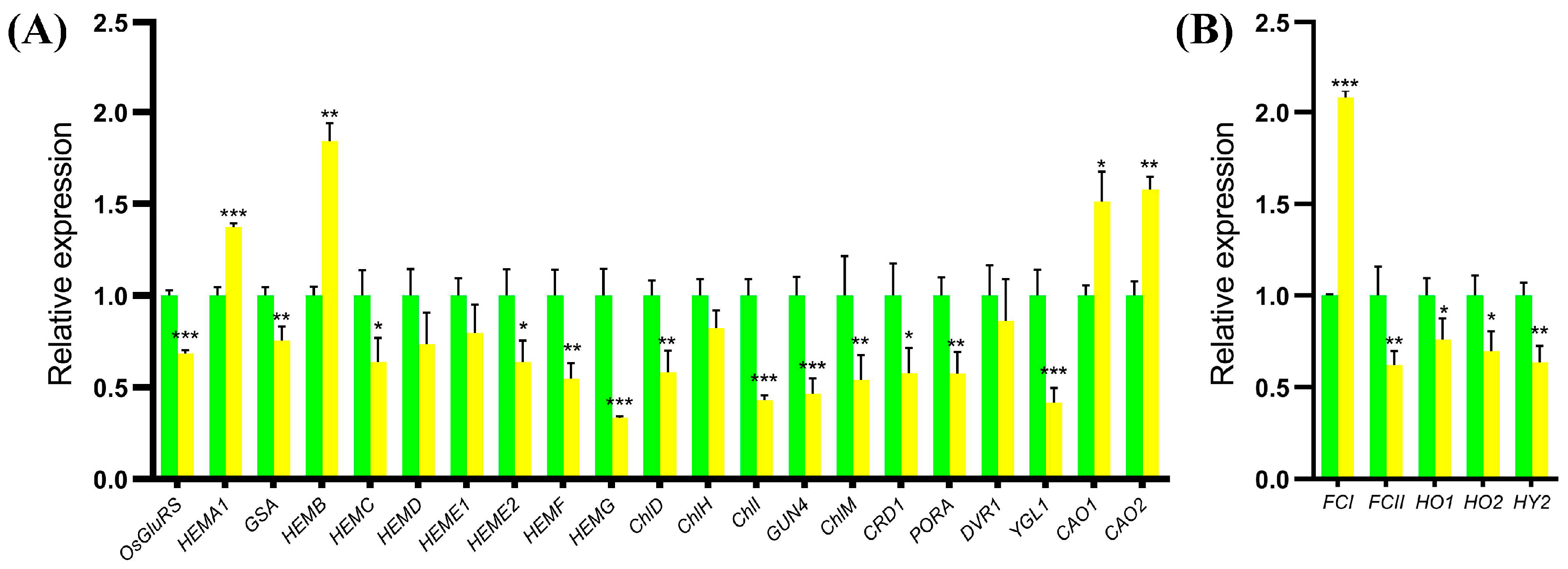
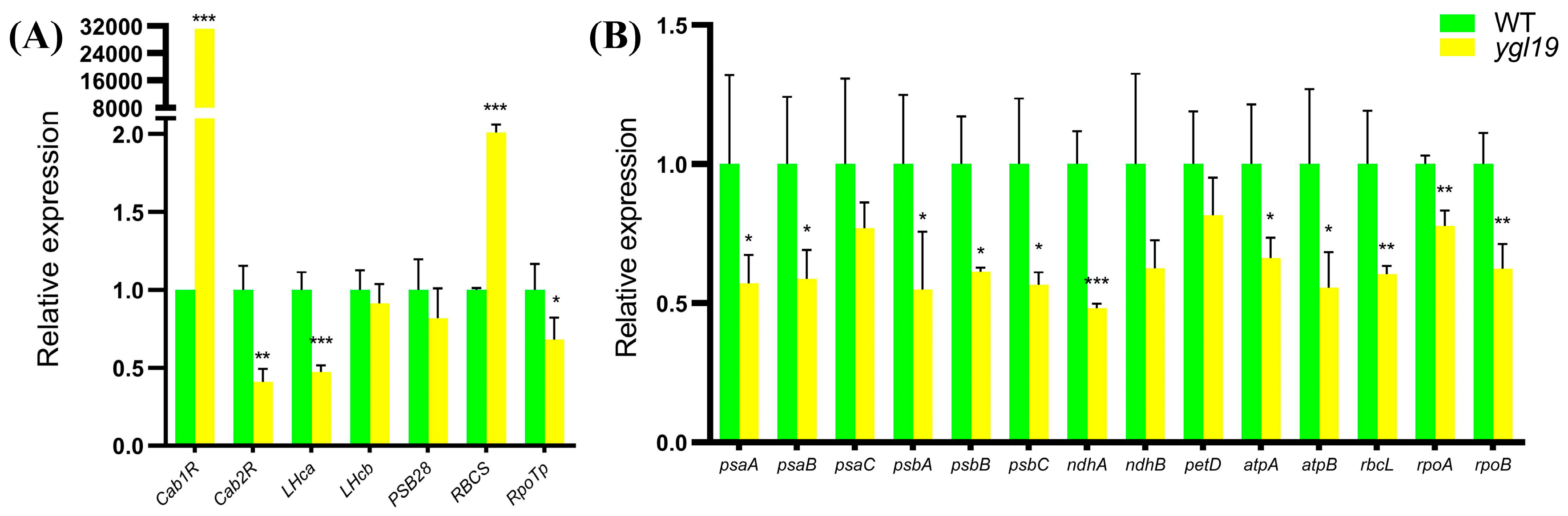
2.8. Expression Analysis of the Genes for Tetrapyrrole Biosynthesis in the ygl19 Mutant
2.9. Expression Analysis of the Genes for Photosynthesis in the ygl19 Mutant
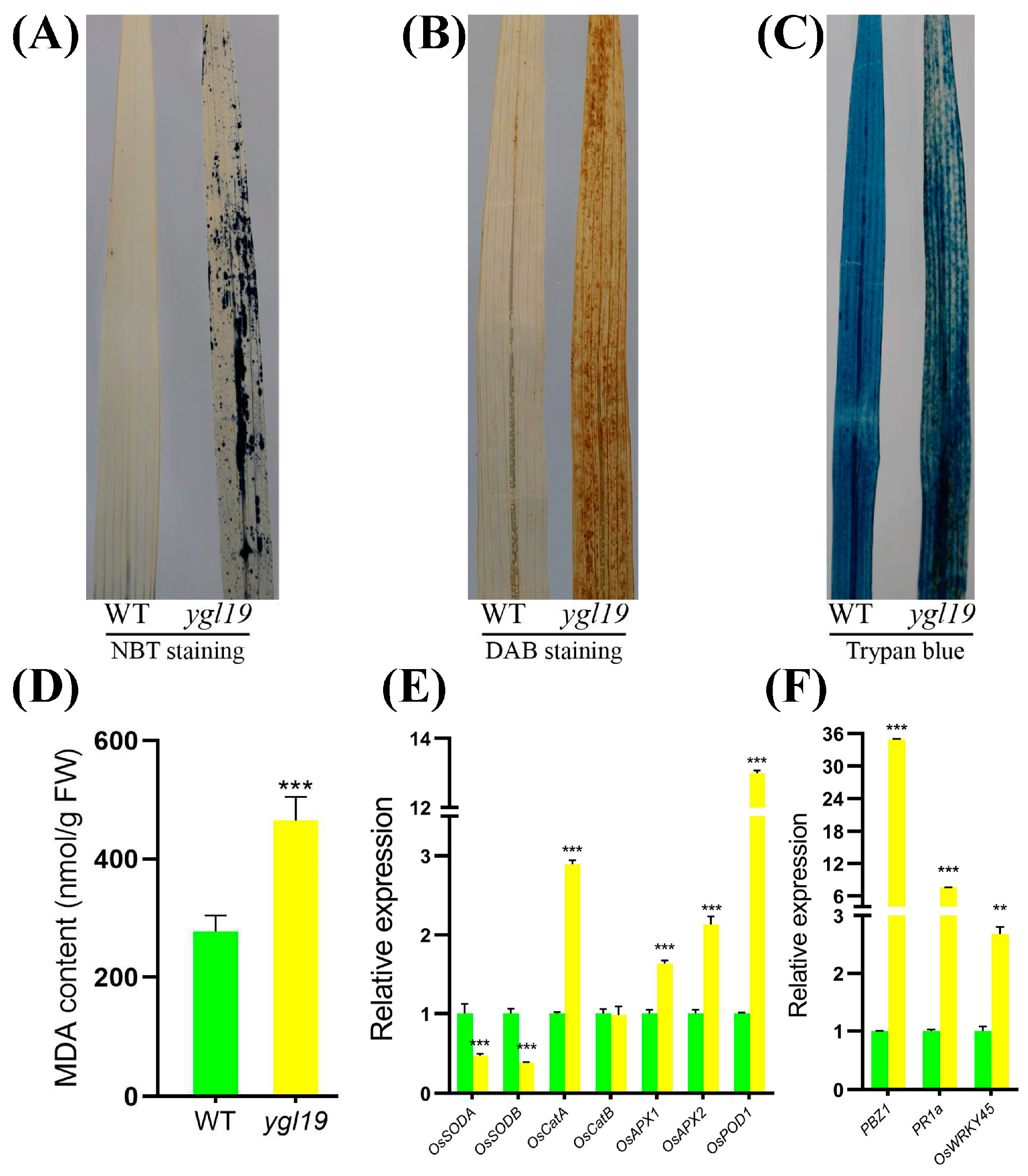
2.10. ROS Accumulation, the Regulation of ROS-Scavenging Genes, and ROS-Responsive Defense Signaling Genes in ygl19 Leaves
3. Discussion
3.1. A Single-Base Substitution of the YGL19 Gene (LOC_Os03g21370) Is Responsible for Phenotype Variation in the ygl19 Mutant
3.2. YGL19 Protein Homologues Are Unique for Photoautotrophs and Contain a Conserved Ycf54 Domain
3.3. The Mutation of the YGL19 Gene Affects the Expression of Tetrapyrrole Biosynthesis-Related Genes, Thereby Regulating the Chlorophyll Content
3.4. The Mutation of the YGL19 Gene Affects the Expression of Photosynthesis-Related Genes, Thereby Regulating Photosynthetic Capacity
3.5. The Mutation of the YGL19 Gene Affects the Expression of ROS Pathway-Related Genes, Thereby Regulating the Metabolism of ROS
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Measurement of Photosynthetic Pigments
4.3. Measurement of the Net Photosynthetic Rate
4.4. Map-Based Cloning of the YGL19 Gene
4.5. Complementation of the YGL19 Gene
4.6. Gene Expression Analysis via qRT-PCR
4.7. Subcellular Localization
4.8. Determination of Reactive Oxygen Species (ROS)
4.9. Data Sources
4.10. Phylogenetic Tree Construction and Multiple Sequence Alignment
4.11. Analysis of Gene Structure and Sequence Motifs
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsugane, K.; Maekawa, M.; Takagi, K.; Takahara, H.; Qian, Q.; Eun, C.H.; Iida, S. An active DNA transposon nDart causing leaf variegation and mutable dwarfism and its related elements in rice. Plant J. 2006, 45, 46–57. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Rahman, M.L.; Cho, S.H.; Kim, Y.S.; Koh, H.J.; Yoo, S.C.; Paek, N.C. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J. 2013, 74, 122–133. [Google Scholar] [CrossRef]
- Wang, Y.S.; Wang, J.J.; Chen, L.Q.; Meng, X.W.; Zhen, X.X.; Liang, Y.P.; Han, Y.; Li, H.; Zhang, B. Identification and function analysis of yellow-leaf mutant (YX-yl) of broomcorn millet. BMC Plant Biol. 2022, 22, 463. [Google Scholar] [CrossRef]
- Awan, M.A.; Konzak, C.F.; Rutger, J.N.; Nilan, R.A. Mutagenic effects of sodium azide in rice. Crop Sci. 1980, 20, 663–668. [Google Scholar] [CrossRef]
- Falbel, T.G.; Andrew, S. Partial blocks in the early steps of the chlorophyll synthesis pathway: A common feathure of chlorophyll b-deficient mutants. Physiol. Plant. 1996, 97, 311–320. [Google Scholar] [CrossRef]
- Voitsekhovskaja, O.V.; Tyutereva, E.V. Chlorophyll b in angiosperms: Functions in photosynthesis, signaling and ontogenetic regulation. J. Plant Physiol. 2015, 189, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Nakagawara, E.; Sakuraba, Y.; Yamasato, A.; Tanaka, R.; Tanaka, A. Clp protease controls chlorophyll b synthesis by regulating the level of chlorophyllide a oxygenase. Plant J. 2007, 49, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Sun, X.Y.; Amombo, E.; Zhu, Q.; Zhao, Z.J.; Chen, L.; Xu, Q.G.; Fu, J.M. High correlation between thermotolerance and photosystem II activity in tall fescue. Photosynth. Res. 2014, 122, 305–314. [Google Scholar] [CrossRef]
- Chen, J.H.; Kell, A.; Acharya, K.; Kupitz, C.; Fromme, P.; Jankowiak, R. Critical assessment of the emission spectra of various photosystem II core complexes. Photosynth. Res. 2015, 124, 253–265. [Google Scholar] [CrossRef]
- Xu, C.H.; Zhu, Q.J.; Chen, J.H.; Shen, L.G.; Yi, X.H.; Huang, Z.H.; Wang, W.; Chen, M.; Kuang, T.; Shen, J.R.; et al. A unique photosystem I reaction center from a chlorophyll d-containing cyanobacterium Acaryochloris marina. J. Integr. Plant Biol. 2021, 63, 1740–1752. [Google Scholar] [CrossRef]
- Lin, Y.P.; Shen, Y.Y.; Shiu, Y.B.; Charng, Y.Y.; Grimm, B. Chlorophyll dephytylase 1 and chlorophyll synthase: A chlorophyll salvage pathway for the turnover of photosystems I and II. Plant J. 2022, 111, 979–994. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Charng, Y.Y. Supraoptimal activity of CHLOROPHYLL DEPHYTYLASE1 results in an increase in tocopherol level in mature arabidopsis seeds. Plant Signal. Behav. 2017, 12, 2974–2990. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.W.; Montgomery, B.L. Interdependence of tetrapyrrole metabolism, the generation of oxidative stress and the mitigative oxidative stress response. Redox Biol. 2015, 4, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, W.; Wang, L.J.; Han, S.; Zhang, Y.Z.; Liu, Q.Z.; Liu, B.S.; Zhao, X.Y. A maize necrotic leaf mutant caused by defect of coproporphyrinogen III oxidase in the porphyrin pathway. Genes 2022, 13, 272. [Google Scholar] [CrossRef]
- Ma, J.J.; Yang, S.X.; Wang, D.M.; Tang, K.Q.; Feng, X.X.; Feng, X.Z. Genetic mapping of a light-dependent lesion mimic mutant reveals the function of coproporphyrinogen III oxidase homolog in soybean. Front. Plant Sci. 2020, 11, 557. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, D.; Sinha, N.; Grimm, B. Thioredoxin-dependent control balances the metabolic activities of tetrapyrrole biosynthesis. Biol. Chem. 2021, 402, 379–397. [Google Scholar] [CrossRef]
- Kobayashi, K.; Masuda, T. Transcriptional Regulation of Tetrapyrrole Biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1811. [Google Scholar] [CrossRef]
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef]
- Richter, A.S.; Grimm, B. Thiol-based redox control of enzymes involved in the tetrapyrrole biosynthesis pathway in plants. Front. Plant Sci. 2013, 4, 371. [Google Scholar] [CrossRef]
- Vasileuskaya, Z.; Oster, U.; Beck, C.F. Mg-protoporphyrin IX and heme control HEMA, the gene encoding the first specific step of tetrapyrrole biosynthesis, in Chlamydomonas reinhardtii. Eukaryot. Cell 2005, 4, 1620–1628. [Google Scholar] [CrossRef]
- Xue, Y.J.; Dong, H.X.; Huang, H.R.; Li, S.P.; Shan, X.H.; Li, H.; Liu, H.; Xia, D.; Su, S.; Yuan, Y. Mutation in Mg-protoporphyrin IX monomethyl ester (oxidative) cyclase gene ZmCRD1 causes chlorophyll-deficiency in maize. Front. Plant Sci. 2022, 13, 912215. [Google Scholar] [CrossRef]
- Herbst, J.; Girke, A.; Hajirezaei, M.R.; Hanke, G.; Grimm, B. Potential roles of YCF54 and ferredoxin-NADPH reductase for magnesium protoporphyrin monomethylester cyclase. Plant J. 2018, 94, 485–496. [Google Scholar] [CrossRef]
- Liu, W.Z.; Fu, Y.P.; Hu, G.C.; Si, H.M.; Zhu, L.; Wu, C.; Sun, Z.X. Identification and fine mapping of a thermo-sensitive chlorophyll deficient mutant in rice (Oryza sativa L.). Planta 2007, 226, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Li, J.J.; Yoo, J.H.; Yoo, S.C.; Cho, S.H.; Koh, H.J.; Seo, H.S.; Paek, N.C. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol. Biol. 2006, 62, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.R.; Wan, C.M.; Xu, Z.J.; Wang, P.Y.; Wang, W.M.; Sun, C.H.; Ma, X.Z.; Xiao, Y.H.; Zhu, J.Q.; Gao, X.L.; et al. One divinyl reductase reduces the 8-vinyl groups in various intermediates of chlorophyll biosynthesis in a given higher plant species, but the isozyme differs between species. Plant Physiol. 2013, 161, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.M.; Zhang, X.; He, B.; Diao, L.P.; Sheng, S.L.; Wang, J.L.; Guo, X.P.; Su, N.; Wang, L.F.; Jiang, L.; et al. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol. 2007, 145, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.M.; Yang, J.; Wu, Z.C.; Liu, H.L.; Huang, F.L.; Wu, Y.R.; Carrie, C.; Narsai, R.; Murcha, M.; Whelan, J.; et al. Identification of a dual-targeted protein belonging to the mitochondrial carrier family that is required for early leaf development in rice. Plant Physiol. 2013, 161, 2036–2048. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Y.; Guo, S.; Wang, Z.W.; Du, Q.; Xing, Y.D.; Zhang, T.Q.; Shen, W.Q.; Sang, X.C.; Ling, Y.H.; He, G.H. Map-based cloning and functional analysis of YGL8, which controls leaf colour in rice (Oryza sativa). BMC Plant Biol. 2016, 16, 134. [Google Scholar] [CrossRef]
- Chen, N.G.; Wang, P.R.; Li, C.M.; Wang, Q.; Pan, J.H.; Xiao, F.L.; Wang, Y.; Zhang, K.; Li, C.; Yang, B.; et al. A single nucleotide mutation of the IspE gene participating in the MEP pathway for isoprenoid biosynthesis causes a green-revertible yellow leaf phenotype in rice. Plant Cell Physiol. 2018, 59, 1905–1917. [Google Scholar] [CrossRef]
- Li, C.M.; Liu, X.; Pan, J.H.; Guo, J.; Wang, Q.; Chen, C.P.; Li, N.; Zhang, K.; Yang, B.; Sun, C.H.; et al. A lil3 chlp double mutant with exclusive accumulation of geranylgeranyl chlorophyll displays a lethal phenotype in rice. BMC Plant Biol. 2019, 19, 456. [Google Scholar] [CrossRef]
- Chen, F.; Dong, G.J.; Wu, L.M.; Wang, F.; Yang, X.Z.; Ma, X.H.; Wang, H.L.; Wu, J.H.; Zhang, Y.L.; Wang, H.Z.; et al. A nucleus-encoded chloroplast protein YL1 is involved in chloroplast development and efficient biogenesis of chloroplast ATP synthase in rice. Sci. Rep. 2016, 6, 32295. [Google Scholar] [CrossRef]
- Wang, Z.H.; Wang, Y.; Hong, X.; Hu, D.H.; Liu, C.X.; Yang, J.; Li, Y.; Huang, Y.Q.; Feng, Y.Q.; Gong, H.Y.; et al. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defence responses in rice. J. Exp. Bot. 2015, 66, 973–987. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Wang, Y.B.; Zhang, Q.; Chen, C.Z.; Qian, Q.; Guo, L.B. WAL3 encoding a PLS-type PPR protein regulates chloroplast development in rice. Plant Sci. 2022, 323, 111382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y.L.; Xie, W.; Chen, C.Z.; Ren, D.Y.; Hu, J.; Zhu, L.; Zhang, G.H.; Gao, Z.Y.; Guo, L.B.; et al. OsMORF9 is necessary for chloroplast development and seedling survival in rice. Plant Sci. 2021, 307, 110907. [Google Scholar] [CrossRef] [PubMed]
- Ambavaram, M.M.; Basu, S.; Krishnan, A.; Ramegowda, V.; Batlang, U.; Rahman, L.; Baisakh, N.; Pereira, A. Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress. Nat. Commun. 2014, 5, 5302. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Qu, M.; Chen, F.; Essemine, J.; Khan, N.; Lyu, M.A.; Chang, T.; Song, Q.; Chen, G.Y.; Zhu, X.G. Overexpression of maize transcription factor mEmBP-1 increases photosynthesis, biomass, and yield in rice. J. Exp. Bot. 2020, 71, 4944–4957. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.Q.; Wang, Y.Q.; Guo, S.; Ma, L.; Wang, Z.W.; Zhu, X.Y.; Sang, X.C.; Ling, Y.H.; Wang, N.; Zhao, F.M.; et al. Molecular mapping and candidate gene analysis of a yellow-green leaf 6 (ygl6) mutant in rice. Crop Sci. 2015, 55, 669–680. [Google Scholar] [CrossRef]
- Ma, X.Z.; Sun, X.Q.; Li, C.M.; Huan, R.; Sun, C.H.; Wang, Y.; Xiao, F.; Wang, Q.; Chen, P.; Ma, F.; et al. Map-based cloning and characterization of the novel yellow-green leaf gene ys83 in rice (Oryza sativa). Plant Physiol. Biochem. 2017, 111, 1–9. [Google Scholar] [CrossRef]
- Zhu, Y.; Yan, P.W.; Dong, S.Q.; Hu, Z.J.; Wang, Y.; Yang, J.S.; Xin, X.Y.; Luo, X.J. Map-based cloning and characterization of YGL22, a new yellow-green leaf gene in rice (Oryza sativa). Crop Sci. 2020, 61, 529–538. [Google Scholar] [CrossRef]
- Huang, R.; Wang, Y.; Wang, P.R.; Li, C.M.; Xiao, F.L.; Chen, N.G.; Li, N.; Li, C.X.; Sun, C.; Li, L.; et al. A single nucleotide mutation of IspF gene involved in the MEP pathway for isoprenoid biosynthesis causes yellow-green leaf phenotype in rice. Plant Mol. Biol. Rep. 2018, 96, 5–16. [Google Scholar] [CrossRef]
- Lan, J.; Lin, Q.B.; Zhou, C.L.; Liu, X.; Miao, R.; Ma, T.F.; Chen, Y.; Mou, C.; Jing, R.; Feng, M.; et al. Young Leaf White Stripe encodes a P-type PPR protein required for chloroplast development. J. Integr. Plant Biol. 2023, 65, 1687–1702. [Google Scholar] [CrossRef]
- Chen, G.E.; Hitchcock, A.; Mares, J.; Gong, Y.; Tichy, M.; Pilny, J.; Kovarova, L.; Zdvihalova, B.; Xu, J.; Hunter, C.N.; et al. Evolution of Ycf54-independent chlorophyll biosynthesis in cyanobacteria. Proc. Natl. Acad. Sci. USA 2021, 118, e2024633118. [Google Scholar] [CrossRef] [PubMed]
- Hollingshead, S.; Kopečná, J.; Armstrong, D.R.; Bučinská, L.; Jackson, P.J.; Chen, G.E.; Dickman, M.J.; Williamson, M.P.; Sobotka, R.; Hunter, C.N. Synthesis of chlorophyll-binding proteins in a fully segregated Δycf54 strain of the cyanobacterium Synechocystis PCC 6803. Front. Plant Sci. 2016, 7, 292. [Google Scholar] [CrossRef] [PubMed]
- Hollingshead, S.; Kopecna, J.; Jackson, P.J.; Canniffe, D.P.; Davison, P.A.; Dickman, M.J.; Sobotka, R.; Hunter, C.N. Conserved chloroplast open-reading frame ycf54 is required for activity of the magnesium protoporphyrin monomethylester oxidative cyclase in Synechocystis PCC 6803. J. Biol. Chem. 2012, 287, 27823–27833. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.E.; Hunter, C.N. Protochlorophyllide synthesis by recombinant cyclases from eukaryotic oxygenic phototrophs and the dependence on Ycf54. Biochem. J. 2020, 477, 2313–2325. [Google Scholar] [CrossRef] [PubMed]
- Bollivar, D.; Braumann, I.; Berendt, K.; Gough, S.P.; Hansson, M. The Ycf54 protein is part of the membrane component of Mg-protoporphyrin IX monomethyl ester cyclase from barley (Hordeum vulgare L.). FEBS J. 2014, 281, 2377–2386. [Google Scholar] [CrossRef]
- Zhao, H.; Li, J.C.; Yang, L.; Qin, G.; Xia, C.J.; Xu, X.B.; Su, Y.M.; Liu, Y.M.; Ming, L.C.; Chen, L.L.; et al. An inferred functional impact map of genetic variants in rice. Mol. Plant. 2021, 14, 1584–1599. [Google Scholar] [CrossRef]
- Zhao, H.; Yao, W.; Ouyang, Y.D.; Yang, W.N.; Wang, G.W.; Lian, X.M.; Xing, Y.Z.; Chen, L.L.; Xie, W.B. RiceVarMap: A comprehensive database of rice genomic variations. Nucleic Acids Res. 2015, 43, D1018–D1022. [Google Scholar] [CrossRef]
- Liu, X.; Deng, X.J.; Li, C.Y.; Xiao, Y.K.; Zhao, K.; Guo, J.; Yang, X.R.; Zhang, H.S.; Chen, C.P.; Luo, Y.T.; et al. Mutation of protoporphyrinogen IX oxidase gene causes spotted and rolled leaf and its overexpression generates herbicide resistance in rice. Int. J. Mol. Sci. 2022, 23, 5781. [Google Scholar] [CrossRef]
- Hollingshead, S.; Bliss, S.; Baker, P.J.; Neil Hunter, C. Conserved residues in Ycf54 are required for protochlorophyllide formation in Synechocystis sp. PCC 6803. Biochem. J. 2017, 474, 667–681. [Google Scholar] [CrossRef]
- Li, Q.Z.; Zhu, F.Y.; Gao, X.L.; Sun, Y.; Li, S.J.; Tao, Y.Z.; Lo, C.; Liu, H.J. Young Leaf Chlorosis 2 encodes the stroma-localized heme oxygenase 2 which is required for normal tetrapyrrole biosynthesis in rice. Planta 2014, 240, 701–712. [Google Scholar] [CrossRef]
- Kim, N.; Jeong, J.; Kim, J.; Oh, J.; Choi, G. Shade represses photosynthetic genes by disrupting the DNA binding of GOLDEN2-LIKE1. Plant Physiol. 2023, 191, 2334–2352. [Google Scholar] [CrossRef] [PubMed]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Dai, S.; Zheng, Y.C.; Li, R.Q.; Tan, Y.Y.; Pan, G.; Moller, I.M.; Song, S.Y.; Huang, J.Z.; Shu, Q.Y. An alanine to valine mutation of glutamyl-tRNA reductase enhances 5-aminolevulinic acid synthesis in rice. Theor. Appl. Genet. 2022, 135, 2817–2831. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.; Eirich, J.; Finkemeier, I.; Grimm, B. Glutamate 1-semialdehyde aminotransferase is connected to GluTR by GluTR-binding protein and contributes to the rate-limiting step of 5-aminolevulinic acid synthesis. Plant Cell 2022, 34, 4623–4640. [Google Scholar] [CrossRef]
- Wang, Z.H.; Hong, X.; Hu, K.K.; Wang, Y.; Wang, X.X.; Du, S.Y.; Li, Y.; Hu, D.D.; Cheng, K.X.; An, B.G.; et al. Impaired magnesium protoporphyrin IX methyltransferase (ChlM) impedes chlorophyll synthesis and plant growth in rice. Front. Plant Sci. 2017, 8, 1694. [Google Scholar] [CrossRef]
- Kong, W.Y.; Yu, X.W.; Chen, H.Y.; Liu, L.L.; Xiao, Y.J.; Wang, Y.L.; Wang, C.L.; Lin, Y.; Yu, Y.; Wang, C.M.; et al. The catalytic subunit of magnesium-protoporphyrin IX monomethyl ester cyclase forms a chloroplast complex to regulate chlorophyll biosynthesis in rice. Plant Mol. Biol. 2016, 92, 177–191. [Google Scholar] [CrossRef]
- Nagata, N.; Tanaka, R.; Satoh, S.; Tanaka, A. Identification of a vinyl reductase gene for chlorophyll synthesis in Arabidopsis thaliana and implications for the evolution of Prochlorococcus species. Plant Cell 2005, 17, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.P.; Wang, J.Y.; Zeng, F.L.; Wang, Q.; Zhu, L.; Li, H.Y.; Guo, N.H.; Chen, H. Photorespiration regulates carbon-nitrogen metabolism by magnesium chelatase D subunit in rice. J. Agric. Food Chem. 2021, 69, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, N.; Izawa, T.; Oikawa, T.; Shimamoto, K. Light regulation of circadian clock-controlled gene expression in rice. Plant J. 2001, 26, 607–615. [Google Scholar] [CrossRef]
- Ye, J.; Ye, S.H.; Zhai, R.R.; Wu, M.M.; Yu, F.M.; Zhu, G.F.; Zhang, X.M. SCL, encoding a chloroplast signal recognition particle receptor, affects chlorophyll synthesis and chloroplast development in rice. J. Plant Growth Regul. 2022, 42, 423–432. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zhang, M.H.; Li, X.Y.; Zhou, R.X.; Xue, X.Y.; Zhang, J.; Liu, N.N.; Xue, R.L.; Qi, X.L. Overexpression of the wheat TaPsb28 gene enhances drought tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2023, 24, 5226. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Suzuki, Y.; Yoshizawa, R.; Kanno, K.; Makino, A. Effect of individual suppression of RBCS multigene family on Rubisco contents in rice leaves. Plant Cell Environ. 2012, 35, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, K.; Yara, A.; Mitsui, N.; Tozawa, Y.; Iba, K. Characterization of a rice nuclear-encoded plastid RNA polymerase gene OsRpoTp. Plant Cell Physiol. 2004, 45, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- An, G.H.; Qi, Y.T.; Zhang, W.Y.; Gao, H.R.; Qian, J.L.; Larkin, R.M.; Chen, J.; Kuang, H. LsNRL4 enhances photosynthesis and decreases leaf angles in lettuce. Plant Biotechnol. J. 2022, 20, 1956–1967. [Google Scholar] [CrossRef] [PubMed]
- Manavski, N.; Torabi, S.; Lezhneva, L.; Arif, M.A.; Frank, W.; Meurer, J. HIGH CHLOROPHYLL FLUORESCENCE145 binds to and stabilizes the psaA 5’ UTR via a newly defined repeat motif in embryophyta. Plant Cell 2015, 27, 2600–2615. [Google Scholar] [CrossRef]
- Peltier, G.; Aro, E.M.; Shikanai, T. NDH-1 and NDH-2 plastoquinone reductases in oxygenic photosynthesis. Annu. Rev. Plant Biol. 2016, 67, 55–80. [Google Scholar] [CrossRef]
- Dixit, R.; Trivedi, P.; Nath, P.; Sane, P.V. Characterization of petB and petD genes of the Populus deltoides chloroplast psbB operon. Plant Mol. Biol. Rep. 2002, 20, 357–368. [Google Scholar] [CrossRef]
- An, H.Q.; Ke, X.L.; Li, L.; Liu, Y.T.; Yuan, S.H.; Wang, Q.Y.; Hou, X.; Zhao, J. ALBINO EMBRYO AND SEEDLING is required for RNA splicing and chloroplast homeostasis in Arabidopsis. Plant Physiol. 2023, 193, 483–501. [Google Scholar] [CrossRef]
- Ermakova, M.; Heyno, E.; Woodford, R.; Massey, B.; Birke, H.; Caemmerer, S.V. Enhanced abundance and activity of the chloroplast ATP synthase in rice through the overexpression of the AtpD subunit. J. Exp. Bot. 2022, 73, 6891–6901. [Google Scholar] [CrossRef]
- Suzuki, Y.; Makino, A. Translational downregulation of RBCL is operative in the coordinated expression of Rubisco genes in senescent leaves in rice. J. Exp. Bot. 2013, 64, 1145–1152. [Google Scholar] [CrossRef]
- Liu, Y.C.; Huang, C.H.; Chang, C.C. A transcriptomic analysis of tobacco leaf with the functional loss of the plastid rpoB operon caused by TALEN-mediated double-strand breakage. Plants-Basel 2022, 11, 2860. [Google Scholar] [CrossRef]
- Liang, J.Y.; Zhang, Q.; Liu, Y.R.; Zhang, J.J.; Wang, W.Y.; Zhang, Z.M. Chlorosis seedling lethality 1 encoding a MAP3K protein is essential for chloroplast development in rice. BMC Plant Biol. 2022, 22, 20. [Google Scholar] [CrossRef]
- Chen, H.F.; Zhang, Q.; Zhang, Z.H. Comparative transcriptome combined with metabolomic and physiological analyses revealed ROS-mediated redox signaling affecting rice growth and cellular iron homeostasis under varying pH conditions. Plant Soil. 2018, 434, 343–361. [Google Scholar] [CrossRef]
- Zhang, S.R.; Apel, K.; Kim, C. Singlet oxygen-mediated and EXECUTER-dependent signalling and acclimation of Arabidopsis thaliana exposed to light stress. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130227. [Google Scholar] [CrossRef]
- Song, S.; Wang, Y.; Ding, X.; Tian, Y.L.; Wu, Z.W.; Li, H.; Li, Q.; Wang, Y.P.; Zhou, S.R.; Dong, X.O.; et al. A plastid RNA polymerase-associated protein is involved in early chloroplast development in rice. Agronomy 2023, 13, 1424. [Google Scholar] [CrossRef]
- Wang, L.L.; Yu, C.C.; Chen, C.; He, C.L.; Zhu, Y.G.; Huang, W.C. Identification of rice Di19 family reveals OsDi19-4 involved in drought resistance. Plant Cell Rep. 2014, 33, 2047–2062. [Google Scholar] [CrossRef]
- Hu, B.; Zhou, Y.F.; Zhou, Z.H.; Sun, B.; Zhou, F.; Yin, C.X.; Ma, W.H.; Chen, H.; Lin, Y.J. Repressed OsMESL expression triggers reactive oxygen species-mediated broad-spectrum disease resistance in rice. Plant Biotechnol. J. 2021, 19, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed. Nat. Protoc. 2008, 3, 824–834. [Google Scholar] [CrossRef]
- Wang, Z.H.; Wang, Y.; Yang, J.; Hu, K.K.; An, B.G.; Deng, X.L.; Li, Y.S. Reliable selection and holistic stability evaluation of reference genes for rice under 22 different experimental conditions. Appl. Biochem. Biotechnol. 2016, 179, 753–775. [Google Scholar] [CrossRef] [PubMed]
- Kariola, T.; Brader, G.; Li, J.; Palva, E.T. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants. Plant Cell 2005, 17, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, R.; Jambunathan, N.; Gunjan, S.K.; Faustin, E.; Weng, H.; Ayoubi, P. Analysis of oxidative signalling induced by ozone in Arabidopsis thaliana. Plant Cell Environ. 2006, 29, 1357–1371. [Google Scholar] [CrossRef] [PubMed]
- Bowling, S.A.; Clarke, J.D.; Liu, Y.; Klessig, D.F.; Dong, X. The cpr5 mutant of Arabidopsis expresses both NPR1-dependent and NPR1-independent resistance. Plant Cell 1997, 9, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kwon, C.T.; Song, G.; Koh, H.J.; An, G.; Paek, N.C. The rice zebra3 (z3) mutation disrupts citrate distribution and produces transverse dark-green/green variegation in mature leaves. Rice 2018, 11, 1. [Google Scholar] [CrossRef]
- Qiao, Y.L.; Jiang, W.Z.; Lee, J.H.; Park, B.S.; Choi, M.S.; Piao, R.H.; Woo, M.O.; Roh, J.H.; Han, L.; Paek, N.C.; et al. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit micro 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol. 2010, 185, 258–274. [Google Scholar] [CrossRef]
- Yin, Z.C.; Chen, J.; Zeng, L.R.; Goh, M.; Leung, H.; Khush, G.S.; Wang, G.L. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol. Plant-Microbe Interact. 2000, 13, 869–876. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Y.L.; Long, W.H.; Niu, M.; Zhao, Z.G.; Teng, X.; Zhu, X.P.; Zhu, J.P.; Hao, Y.Y.; Wang, Y.F.; et al. SGD1, a key enzyme in tocopherol biosynthesis, is essential for plant development and cold tolerance in rice. Plant Sci. 2017, 260, 90–100. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cross | Wild-Type Green Leaf | Yellow-Green and Spotted Leaf | χ2 (3:1) | p Value |
|---|---|---|---|---|
| ygl19/9311 | 144 | 44 | 0.18 | 0.92 |
| ygl19/02428 | 3173 | 1062 | 0.01 | 0.99 |
| SNP Position | Primary Allele | Secondary Allele | Primary Allele Frequency | Amino Acid Mutation and Position | snpEff Annotation |
|---|---|---|---|---|---|
| 28 | A | G | 99.90% | no | 5_prime_UTR_variant |
| 180 | T | G | 60.40% | Ala16Ala | synonymous_variant |
| 321 | C | T | 60.30% | Asp63Asp | synonymous_variant |
| 559 | A | G | 59.90% | Val111Val | synonymous_variant |
| 742 | A | T | 99.90% | no | intron_variant |
| 764 | A | C | 93.60% | no | intron_variant |
| 853 | C | T | 99.70% | no | intron_variant |
| 1304 | T | C | 59.90% | no | intron_variant |
| 1391 | A | G | 60.10% | no | intron_variant |
| 1420 | T | C | 60.00% | no | intron_variant |
| 1508 | G | A | 60.30% | no | intron_variant |
| 1686 | G | A | 94.40% | no | intron_variant |
| 1748 | G | A | 99.80% | no | intron_variant |
| 1817 | C | T | 99.90% | no | intron_variant |
| 1918 | C | T | 99.80% | no | intron_variant |
| 2000 | T | C | 60.00% | no | intron_variant |
| 2226 | T | C | 59.90% | no | intron_variant |
| 2243 | A | T | 91.30% | no | intron_variant |
| 2351 | G | A | 99.90% | no | intron_variant |
| 2427 | G | A | 98.90% | no | intron_variant |
| 2492 | G | A | 99.90% | no | intron_variant |
| 2520 | G | A | 99.30% | no | intron_variant |
| 2732 | T | C | 59.90% | no | intron_variant |
| 2917 | T | C | 99.20% | no | splice_region_variant&intron_variant |
| 3212 | T | G | 60.20% | no | 3_prime_UTR_variant |
| 3405 | C | A | 96.40% | no | 3_prime_UTR_variant |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhang, H.; Wei, L.; Guo, R.; Liu, X.; Zhang, M.; Fan, J.; Liu, S.; Liao, J.; Huang, Y.; et al. Yellow-Green Leaf 19 Encoding a Specific and Conservative Protein for Photosynthetic Organisms Affects Tetrapyrrole Biosynthesis, Photosynthesis, and Reactive Oxygen Species Metabolism in Rice. Int. J. Mol. Sci. 2023, 24, 16762. https://doi.org/10.3390/ijms242316762
Wang Q, Zhang H, Wei L, Guo R, Liu X, Zhang M, Fan J, Liu S, Liao J, Huang Y, et al. Yellow-Green Leaf 19 Encoding a Specific and Conservative Protein for Photosynthetic Organisms Affects Tetrapyrrole Biosynthesis, Photosynthesis, and Reactive Oxygen Species Metabolism in Rice. International Journal of Molecular Sciences. 2023; 24(23):16762. https://doi.org/10.3390/ijms242316762
Chicago/Turabian StyleWang, Qiang, Hongyu Zhang, Lingxia Wei, Rong Guo, Xuanzhi Liu, Miao Zhang, Jiangmin Fan, Siyi Liu, Jianglin Liao, Yingjin Huang, and et al. 2023. "Yellow-Green Leaf 19 Encoding a Specific and Conservative Protein for Photosynthetic Organisms Affects Tetrapyrrole Biosynthesis, Photosynthesis, and Reactive Oxygen Species Metabolism in Rice" International Journal of Molecular Sciences 24, no. 23: 16762. https://doi.org/10.3390/ijms242316762
APA StyleWang, Q., Zhang, H., Wei, L., Guo, R., Liu, X., Zhang, M., Fan, J., Liu, S., Liao, J., Huang, Y., & Wang, Z. (2023). Yellow-Green Leaf 19 Encoding a Specific and Conservative Protein for Photosynthetic Organisms Affects Tetrapyrrole Biosynthesis, Photosynthesis, and Reactive Oxygen Species Metabolism in Rice. International Journal of Molecular Sciences, 24(23), 16762. https://doi.org/10.3390/ijms242316762