
The Role of CPNE7 (Copine-7) in Colorectal Cancer Prognosis and Metastasis

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Relationship of CPNE7 Expression and Clinicopathologic Factor of Patients with CRC
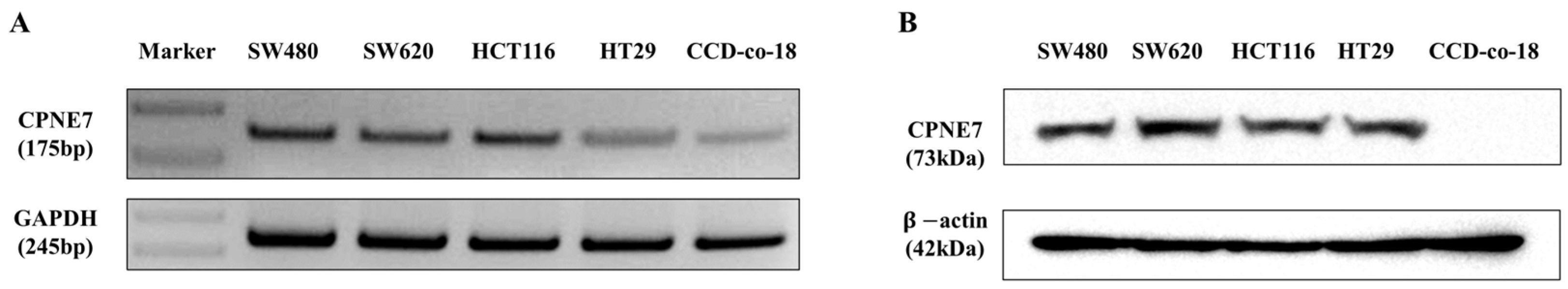
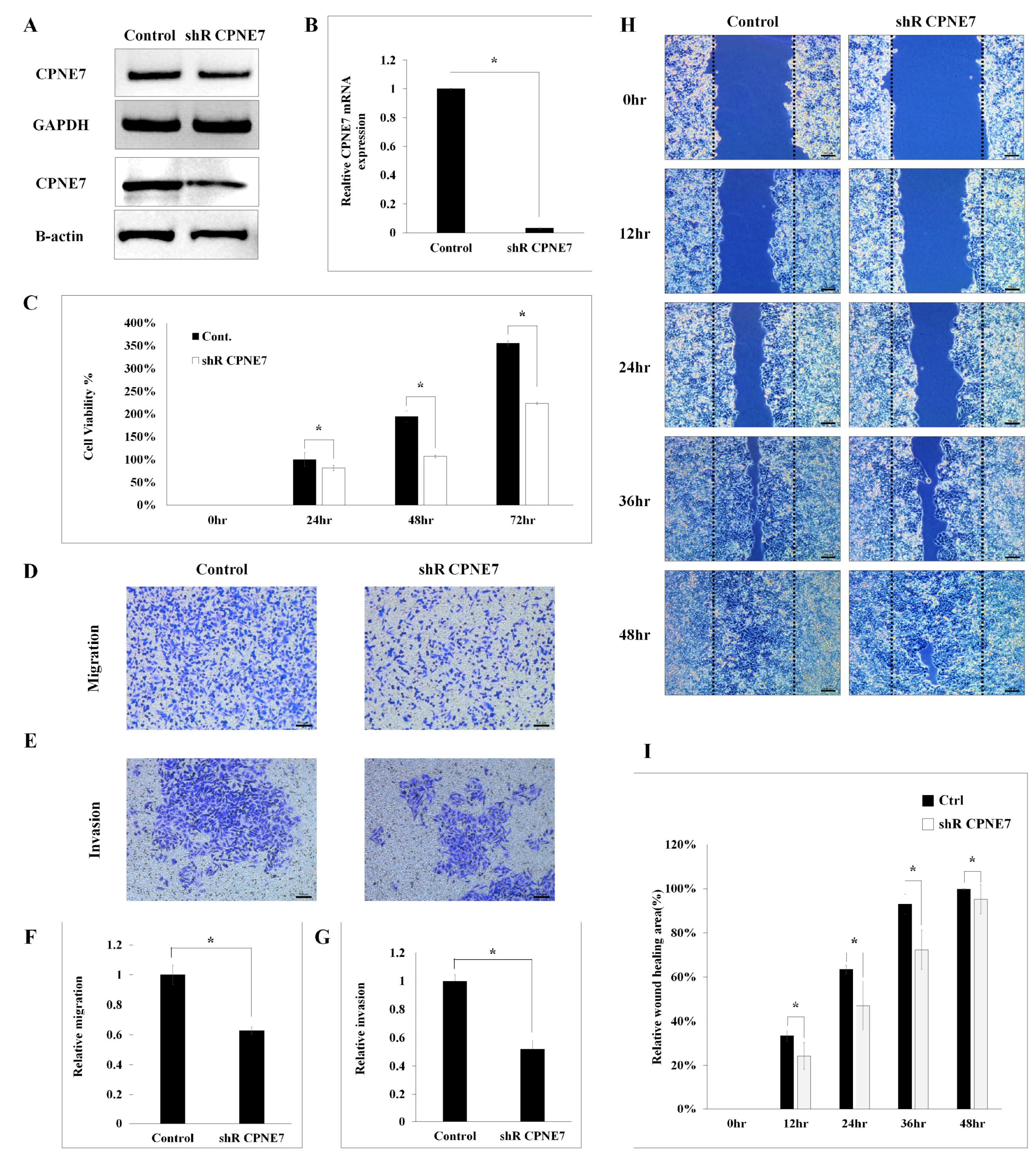
2.2. Functional Effect of CPNE7 in CRC Cell Lines
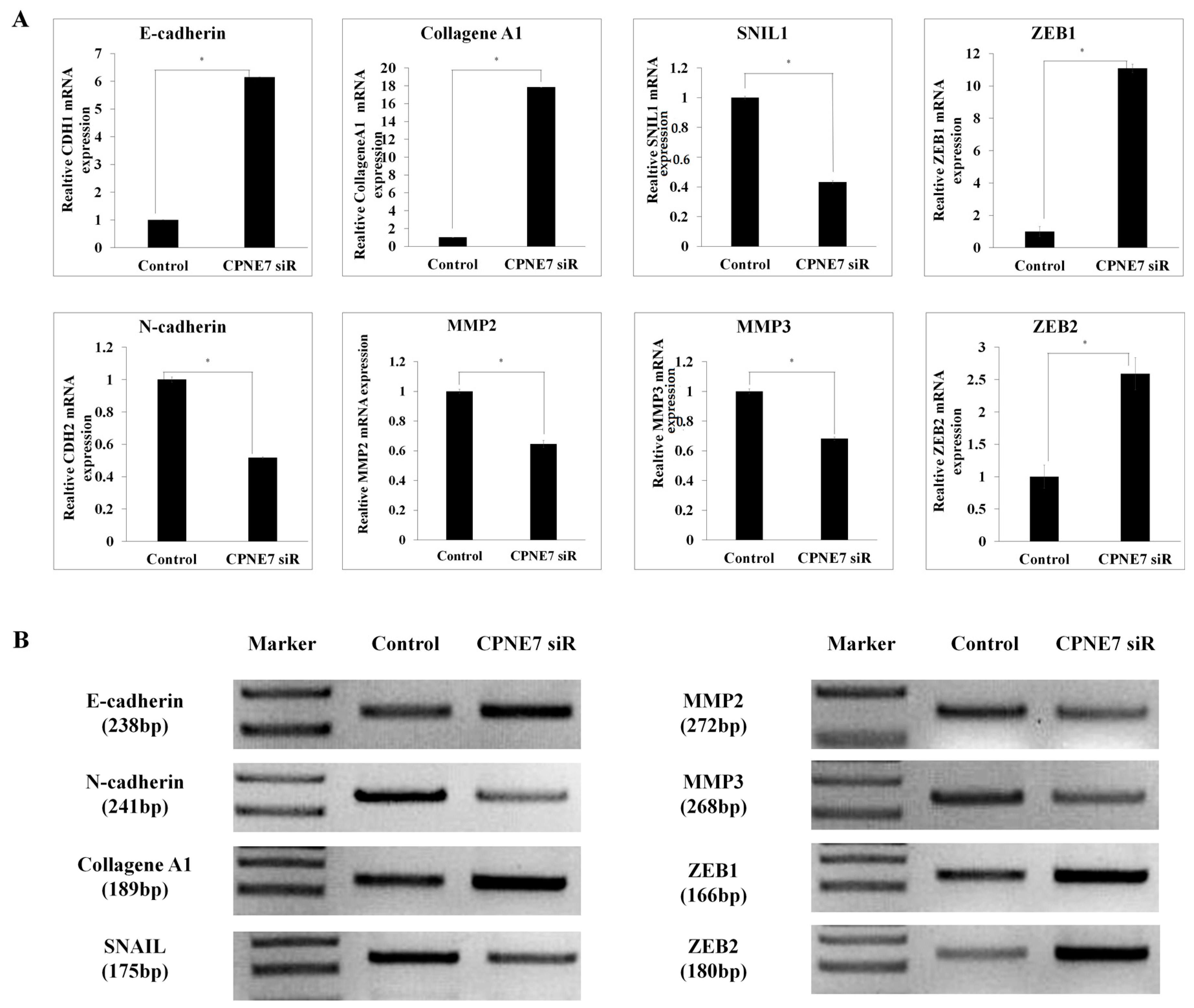
2.3. CPNE7 Inhibition Leads to the Dysregulation of EMT Signatures in CRC Cells
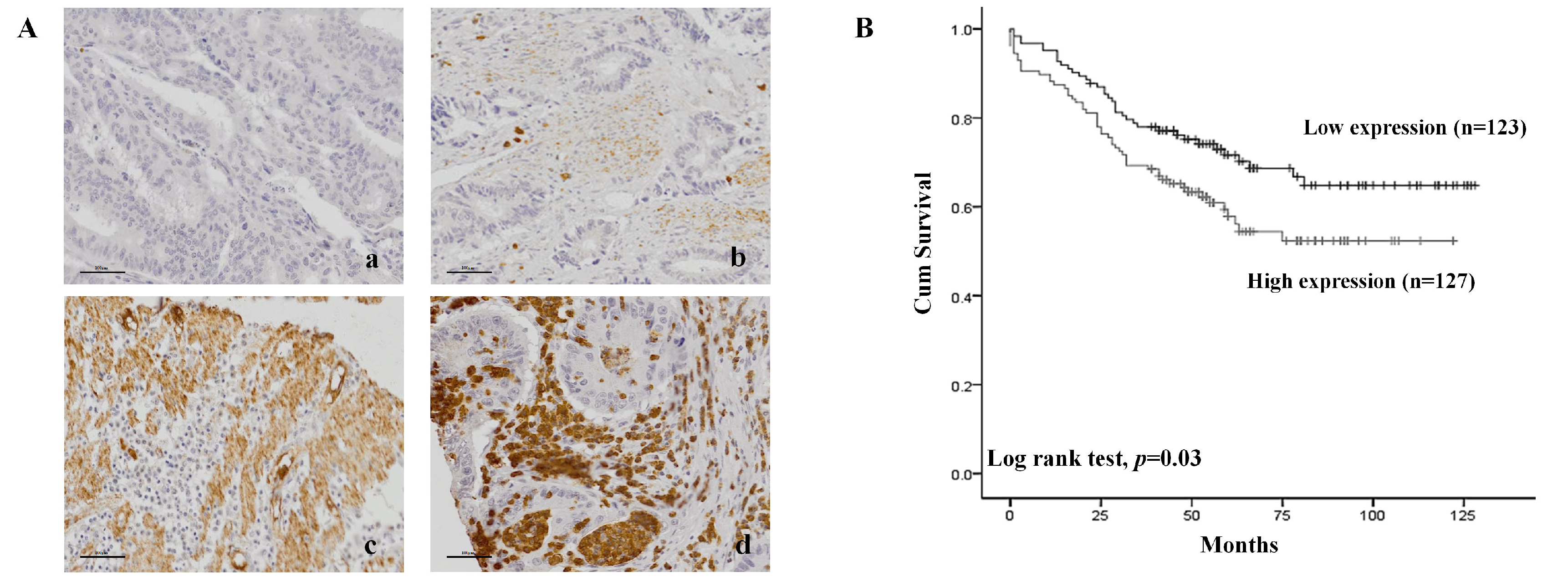
2.4. Patient Survival Rate According to CPNE7 Expression in FFPE Tissue in Colorectal Cancer Patients
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture
4.2. Tissue Specimens from Patients
4.3. Small Interfering RNA (siRNA) and Transient Transfection
4.4. Short Hairpin RNA (shRNA) and Transient Transfection
4.5. Real-Time Polymerase Chain Reaction
4.6. Cell Proliferation Analysis
4.7. Transwell Migration Assay
4.8. Transwell Invasion Assay
4.9. Semisolid Agar Colony-Forming Assay
4.10. Western Blot
4.11. Immunohistochemistry (IHC)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F.; Bsc, M.F.B.; Me, J.F.; Soerjomataram, M.I.; et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Won, Y.J.; Lee, J.J.; Jung, K.W.; Kong, H.J.; Im, J.S.; Seo, H.G. Cancer statistics in Korea: Incidence, mortality, survival, and prevalence in 2018. Cancer Res. Treat. Off. J. Korean Cancer Assoc. 2021, 53, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed]
- Muratore, A.; Zorzi, D.; Bouzari, H.; Amisano, M.; Massucco, P.; Sperti, E.; Capussotti, L. Asymptomatic colorectal cancer with un-resectable liver metastases: Immediate colorectal resection or up-front systemic chemotherapy? Ann. Surg. Oncol. 2007, 14, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Riihimäki, M.; Hemminki, A.; Sundquist, J.; Hemminki, K. Patterns of metastasis in colon and rectal cancer. Sci. Rep. 2016, 6, 29765. [Google Scholar] [CrossRef]
- Tang, H.; Pang, P.; Qin, Z.; Zhao, Z.; Wu, Q.; Song, S.; Li, F. The CPNE family and their role in cancers. Front. Genet. 2021, 12, 689097. [Google Scholar] [CrossRef] [PubMed]
- Damer, C.K.; Bayeva, M.; Hahn, E.S.; Rivera, J.I.; Socec, C. Copine A, a calcium-dependent membrane-binding protein, transiently localizes to the plasma membrane and intracellular vacuoles in Dictyostelium. BMC Cell Biol. 2005, 6, 46. [Google Scholar] [CrossRef]
- Ilacqua, A.N.; Price, J.E.; Graham, B.N.; Buccilli, M.J.; McKellar, D.R.; Damer, C.K. Cyclic AMP signaling in Dictyostelium promotes the translocation of the copine family of calcium-binding proteins to the plasma membrane. BMC Cell Biol. 2018, 19, 13. [Google Scholar] [CrossRef]
- Perestenko, P.V.; Pooler, A.M.; Noorbakhshnia, M.; Gray, A.; Bauccio, C.; McIlhinney, R.A.J. Copines-1, -2, -3, -6 and -7 show different calcium-dependent intracellular membrane translocation and targeting: Calcium-dependent translocation of copines. FEBS J. 2010, 277, 5174–5189. [Google Scholar] [CrossRef]
- Pan, H.; Xu, X.; Wu, D.; Qiu, Q.; Zhou, S.; He, X.; Zhou, Y.; Qu, P.; Hou, J.; He, J.; et al. Novel somatic mutations identified by whole-exome sequencing in muscle-invasive transitional cell carcinoma of the bladder. Oncol. Lett. 2016, 11, 1486–1492. [Google Scholar] [CrossRef]
- Ji, X.; Sun, T.; Xie, S.; Qian, H.; Song, L.; Wang, L.; Liu, L.; Feng, Q. Upregulation of CPNE7 in mesenchymal stromal cells promotes oral squamous cell carcinoma metastasis through the NF-κB pathway. Cell Death Discov. 2021, 7, 294. [Google Scholar] [CrossRef]
- Dekker, E.; Sanduleanu, S. Colorectal cancer: Strategies to minimize interval CRC in screening programmes: Colorectal cancer, Nature reviews. Gastroenterol. Hepatol. 2016, 13, 10–12. [Google Scholar]
- Kim, Y.J.; Kang, D.H.; Song, G.J.; Ahn, T.S.; Son, M.W.; Lee, M.S. Clinical relevance of Lgr5 expression in colorectal cancer patients. Korean J. Clin. Oncol. 2018, 14, 76–82. [Google Scholar] [CrossRef]
- Shin, E.J.; Kim, H.J.; Son, M.W.; Ahn, T.S.; Lee, H.Y.; Lim, D.R.; Bae, S.B.; Jeon, S.; Kim, H.; Jeong, D.; et al. Epigenetic inactivation of RUNX3 in colorectal cancer. Ann. Surg. Treat Res. 2018, 94, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Lobbes, L.A.; Schütze, M.A.; Droeser, R.; Arndt, M.; Pozios, I.; Lauscher, J.C.; Hering, N.A.; Weixler, B. Muscarinic Acetylcholine Receptor M3 Expression and Survival in Human Colorectal Carcinoma—An Unexpected Correlation to Guide Future Treatment? Int. J. Mol. Sci. 2023, 24, 8198. [Google Scholar] [CrossRef]
- Kikuchi, R.; Takano, M.; Takagi, K.; Fujimoto, N.; Nozaki, R.; Fujiyoshi, T.; Uchida, Y. Management of early invasive colorectal cancer. Risk of recurrence and clinical guidelines. Dis. Colon Rectum 1995, 38, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Wibe, A.; Syse, A.; Andersen, E.; Tretli, S.; Myrvold, H.E.; Søreide, O.; Norwegian Rectal Cancer Group. Oncological outcomes after total mesorectal excision for cure for cancer of the lower rectum: Anterior vs abdominoperineal resection. Dis. Colon Rectum 2004, 47, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, D.; Roh, S.; Hong, I.; Kim, H.; Ahn, T.S.; Kang, D.H.; Lee, M.S.; Baek, M.-J.; Kwak, H.J.; et al. Expression of Spermine Oxidase Is Associated with Colorectal Carcinogenesis and Prognosis of Patients. Bio-Medicines 2022, 10, 626. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, E.; Bösch, F.; Neumann, J.; Ganschow, P.; Bazhin, A.; Guba, M.; Werner, J.; Angele, M. Mechanisms of metastasis in colorectal cancer and metastatic organotropism: Hematogenous versus peritoneal spread. J. Oncol. 2019, 2019, 7407190. [Google Scholar] [CrossRef]
- Jaffe, L.F. A calcium-based theory of carcinogenesis. Adv. Cancer Res. 2005, 94, 231–263. [Google Scholar]
- Kadio, B.; Yaya, S.; Basak, A.; Djè, K.; Gomes, J.; Mesenge, C. Calcium role in human carcinogenesis: A comprehensive analysis and critical review of literature. Cancer Metastasis Rev. 2016, 35, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Stewart, T.A.; Yapa, K.T.; Monteith, G.R. Altered calcium signaling in cancer cells. Biochim. Et Biophys. Acta 2015, 1848 Pt B, 2502–2511. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, T.; Singh, R.; Sabharwal, L.; Bando, H.; Meng, J.; Arima, Y.; Yamada, M.; Harada, M.; Jiang, J.-J.; Kamimura, D.; et al. Inflammation amplifier, a new paradigm in cancer biology. Cancer Res. 2014, 74, 8–14. [Google Scholar] [CrossRef]
- Chen, J.; Wu, S.; Peng, Y.; Zhao, Y.; Dong, Y.; Ran, F.; Geng, H.; Zhang, K.; Li, J.; Huang, S.; et al. Constructing a cancer stem cell related prognostic model for predicting immune landscape and drug sensitivity in colorectal cancer. Front. Pharmacol. 2023, 14, 1200017. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.-A.; Kim, S.-H.; Hong, H.K.; Yun, S.H.; Kim, H.C.; Chun, H.-K.; Cho, Y.B.; Lee, W.Y. Loss of E-Cadherin expression is associated with a poor prognosis in stage III colorectal cancer. Oncology 2014, 86, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yan, L.; Liu, S.; Shan, Z.; Tian, Y.; Jin, Z. N-cadherin, a novel prognostic biomarker, drives malignant progression of colorectal cancer. Mol. Med. Rep. 2015, 12, 2999–3006. [Google Scholar] [CrossRef]
- Li, Y.Y.; Zhou, C.X.; Gao, Y. Podoplanin promotes the invasion of oral squamous cell carcinoma in coordination with MT1-MMP and Rho GTPases. Am. J. Cancer Res. 2015, 5, 514–529. [Google Scholar]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Factors | CPNE7 | Total | p Value | |
|---|---|---|---|---|
| High Expression (N = 127) | Low Expression (N = 123) | |||
| Age, years, mean (SD) | 63.1 (12.6) | 63.0 (12.7) | 63.10 (12.6) | 0.181 |
| Sex, N (%) | 0.124 | |||
| Male | 69 (27.6) | 79 (31.61) | 148 (59.2) | |
| Female | 58 (23.2) | 44 (17.6) | 102 (40.8) | |
| TMN stage, N (%) | 0.820 | |||
| I | 17 (6.8) | 20 (8.0) | 37 (14.8) | |
| II | 52 (20.8) | 47 (18.8) | 99 (39.6) | |
| III | 53 (21.2) | 49 (19.6) | 102 (40.8) | |
| IV | 5 (2.0) | 7 (2.8) | 12 (4.8) | |
| Vascular invasion, N (%) | 0.481 | |||
| 0 | 102 (40.8) | 103 (41.2) | 205 (82.0) | |
| 1 | 25 (10.0) | 20 (8.0) | 45 (18.0) | |
| Lymphatic invasion, N (%) | 0.230 | |||
| 0 | 93 (37.2) | 98 (39.2) | 191 (76.4) | |
| 1 | 34 (13.6) | 25 (10.0) | 59 (23.6) | |
| Clinicopathological Factors | Multivariate Analysis | Univariate Analysis | ||
|---|---|---|---|---|
| Hazard Ratio (95% CI) | p | Hazard Ratio (95% CI) | p | |
| Age, years, mean (SD) | 1.63(1.00–2.68) | 0.050 | 1.51 (0.94–2.42) | 0.087 |
| Sex, N (%) | ||||
| Female vs. Male | 0.98 (0.64–1.51) | 0.952 | 1.13 (0.74–1.72) | 0.548 |
| TNM stage, N (%) | 0.005 * | 0.001 * | ||
| I | 1.00 | 1.00 | ||
| II | 1.08 (0.50–2.30) | 0.838 | 1.16 (0.54–2.26) | 0.692 |
| III | 1.90 (0.91–3.95) | 0.085 | 2.01 (0.98–4.13) | 0.055 |
| IV | 4.05 (1.48–11.09) | 0.006 * | 4.60 (1.77–11.96) | 0.002 * |
| Venous Invasion | ||||
| High vs. Low | 0.87 (0.41–1.85) | 0.735 | 1.59 (0.97–2.59) | 0.062 |
| Lymphatic Invasion | ||||
| High vs. Low | 1.74 (0.89–3.37) | 0.102 | 1.94 (1.25–3.03) | 0.003 * |
| CPNE7 expression | ||||
| High vs. Low | 1.54 (1.00–2.38) | 0.048 * | 1.58 (1.03–2.41) | 0.032 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, H.-J.; Kang, D.-H.; Ahn, T.-S.; Kim, K.-S.; Kim, T.-W.; Lee, S.-H.; Lee, D.-W.; Ryu, J.-S.; Beak, M.-J. The Role of CPNE7 (Copine-7) in Colorectal Cancer Prognosis and Metastasis. Int. J. Mol. Sci. 2023, 24, 16704. https://doi.org/10.3390/ijms242316704
Kong H-J, Kang D-H, Ahn T-S, Kim K-S, Kim T-W, Lee S-H, Lee D-W, Ryu J-S, Beak M-J. The Role of CPNE7 (Copine-7) in Colorectal Cancer Prognosis and Metastasis. International Journal of Molecular Sciences. 2023; 24(23):16704. https://doi.org/10.3390/ijms242316704
Chicago/Turabian StyleKong, Hye-Jeong, Dong-Hyun Kang, Tae-Sung Ahn, Kwang-Seock Kim, Tae-Wan Kim, Soo-Hyeon Lee, Dong-Woo Lee, Jae-Sung Ryu, and Moo-Jun Beak. 2023. "The Role of CPNE7 (Copine-7) in Colorectal Cancer Prognosis and Metastasis" International Journal of Molecular Sciences 24, no. 23: 16704. https://doi.org/10.3390/ijms242316704
APA StyleKong, H.-J., Kang, D.-H., Ahn, T.-S., Kim, K.-S., Kim, T.-W., Lee, S.-H., Lee, D.-W., Ryu, J.-S., & Beak, M.-J. (2023). The Role of CPNE7 (Copine-7) in Colorectal Cancer Prognosis and Metastasis. International Journal of Molecular Sciences, 24(23), 16704. https://doi.org/10.3390/ijms242316704