

Ouabain’s Influence on TRPV4 Channels of Epithelial Cells: An Exploration of TRPV4 Activity, Expression, and Signaling Pathways
 ,
,
Abstract
:1. Introduction
2. Results
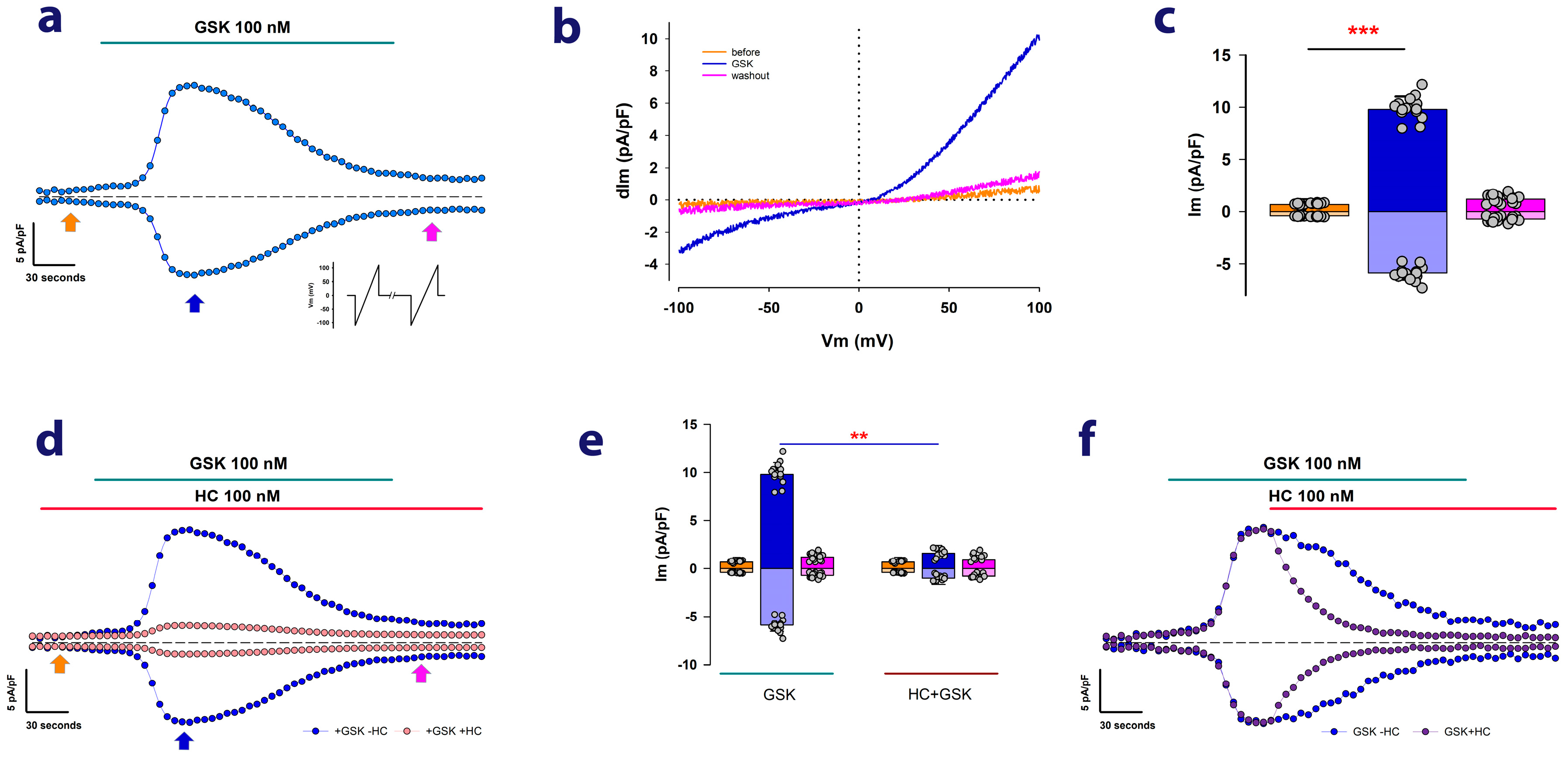
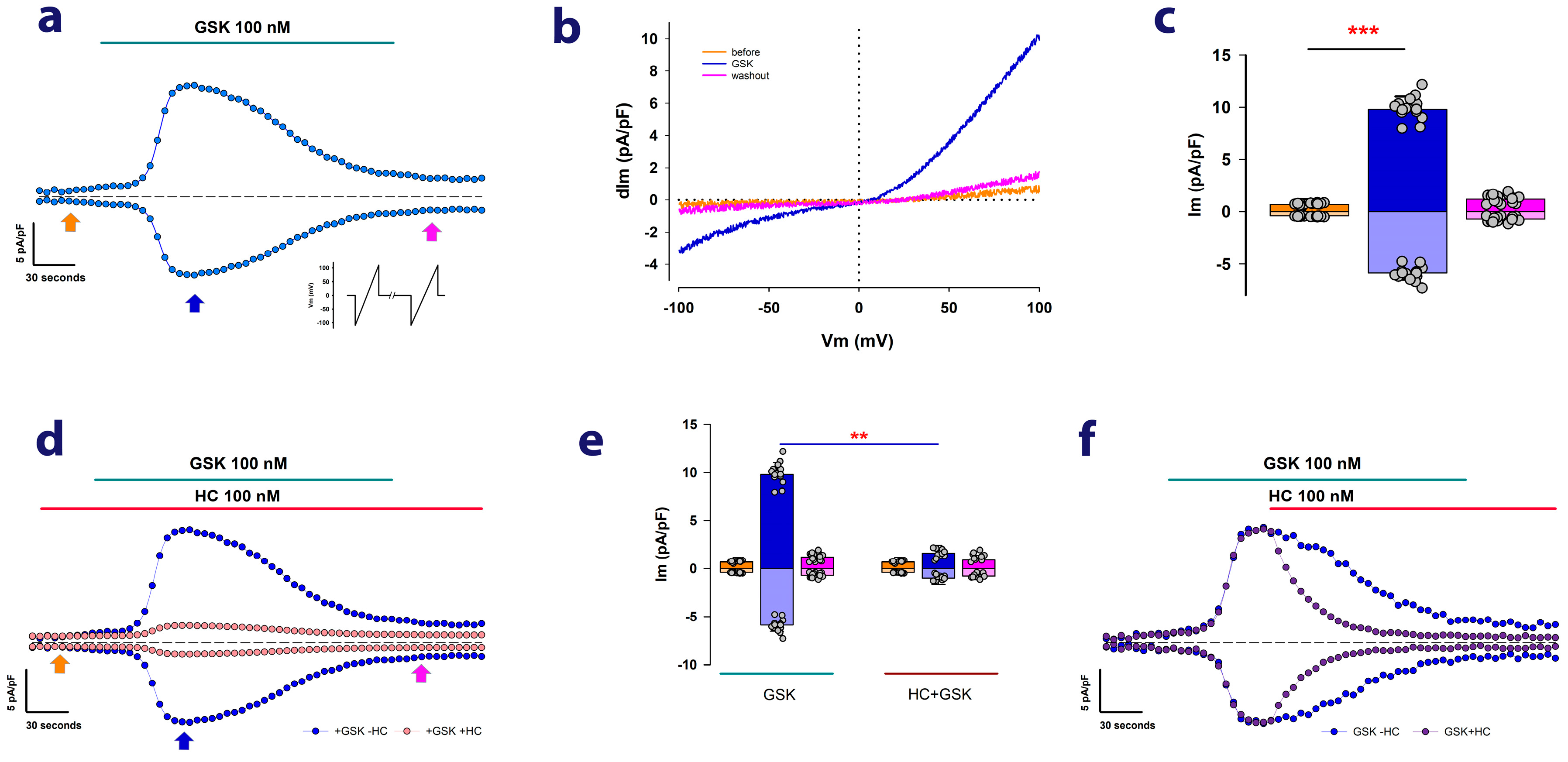
2.1. MDCK Cells Cultured as Mature Epithelial Monolayers Exhibit Ion Currents Attributable to TRPV4 Channels
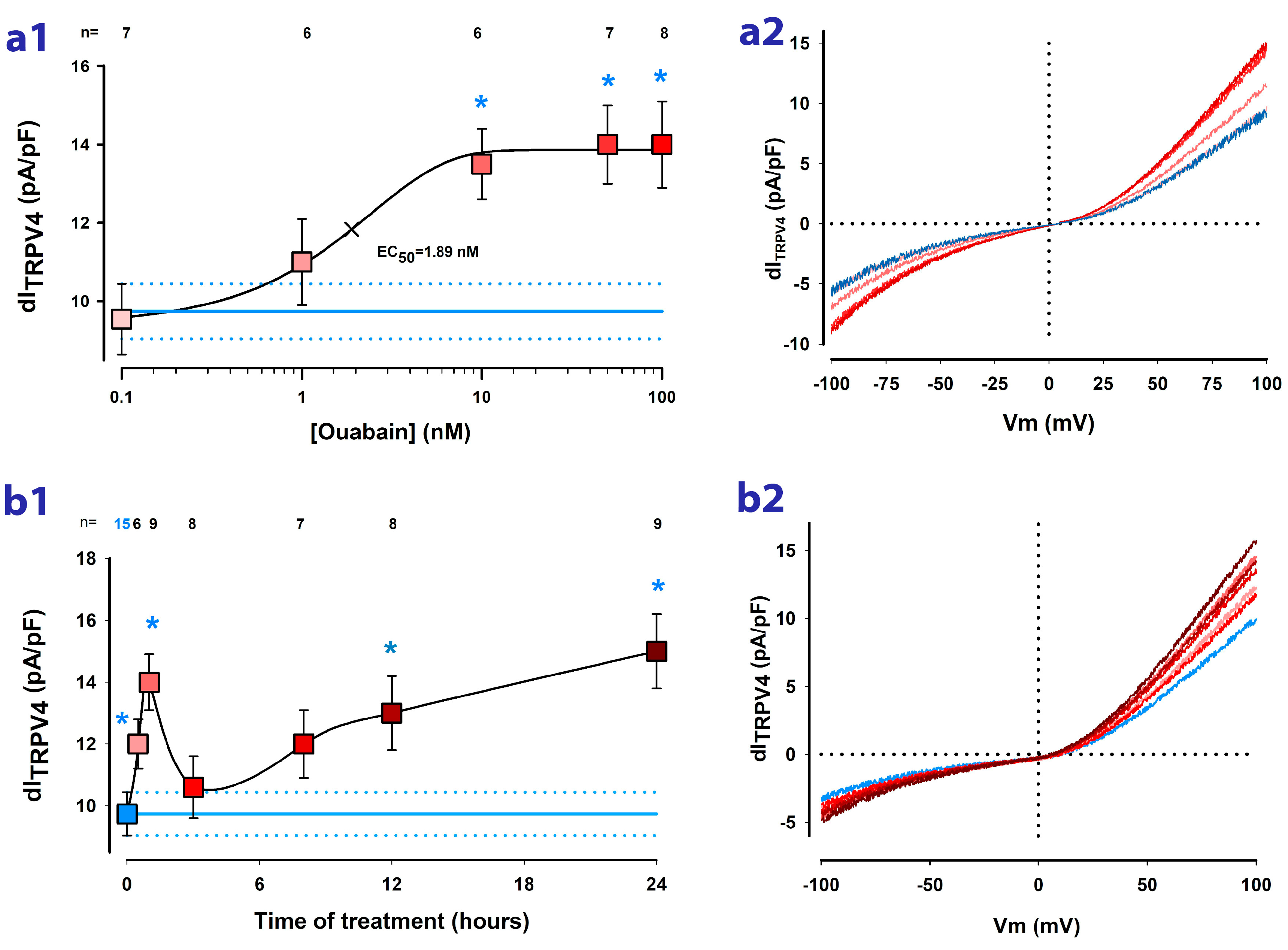
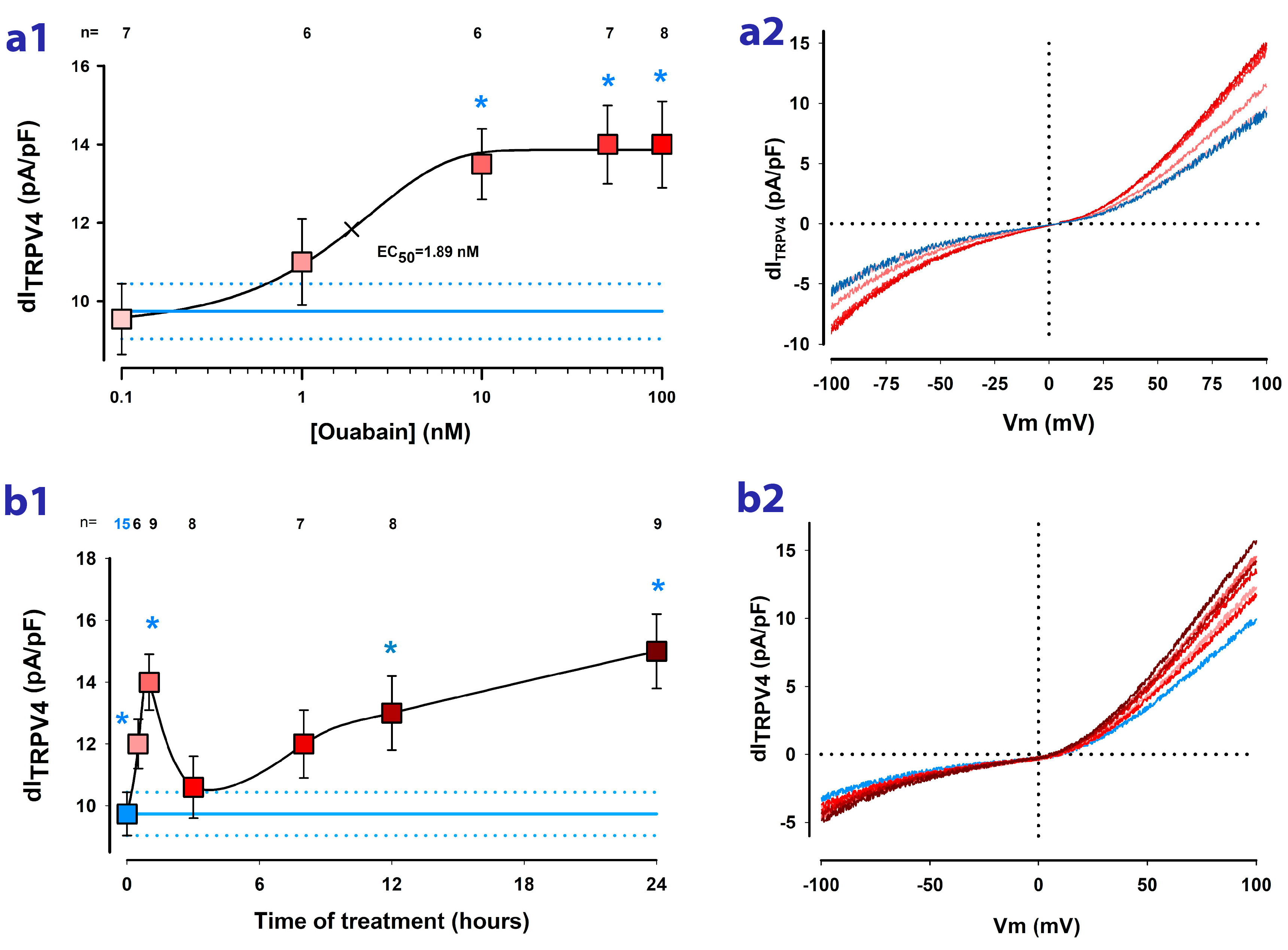
2.2. Ouabain Enhances the dITRPV4 of Cells Cultured as Mature Epithelial Monolayers
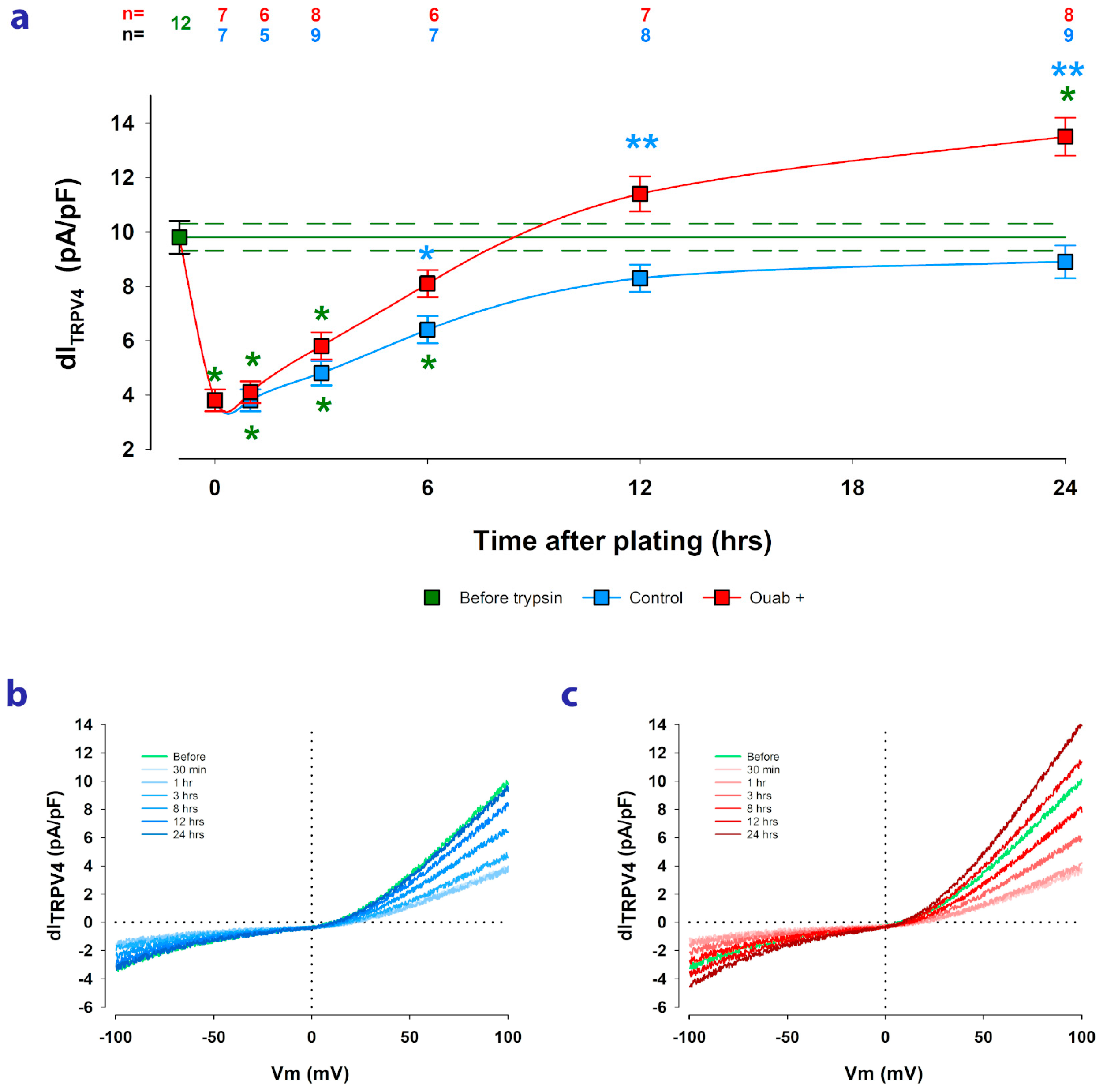
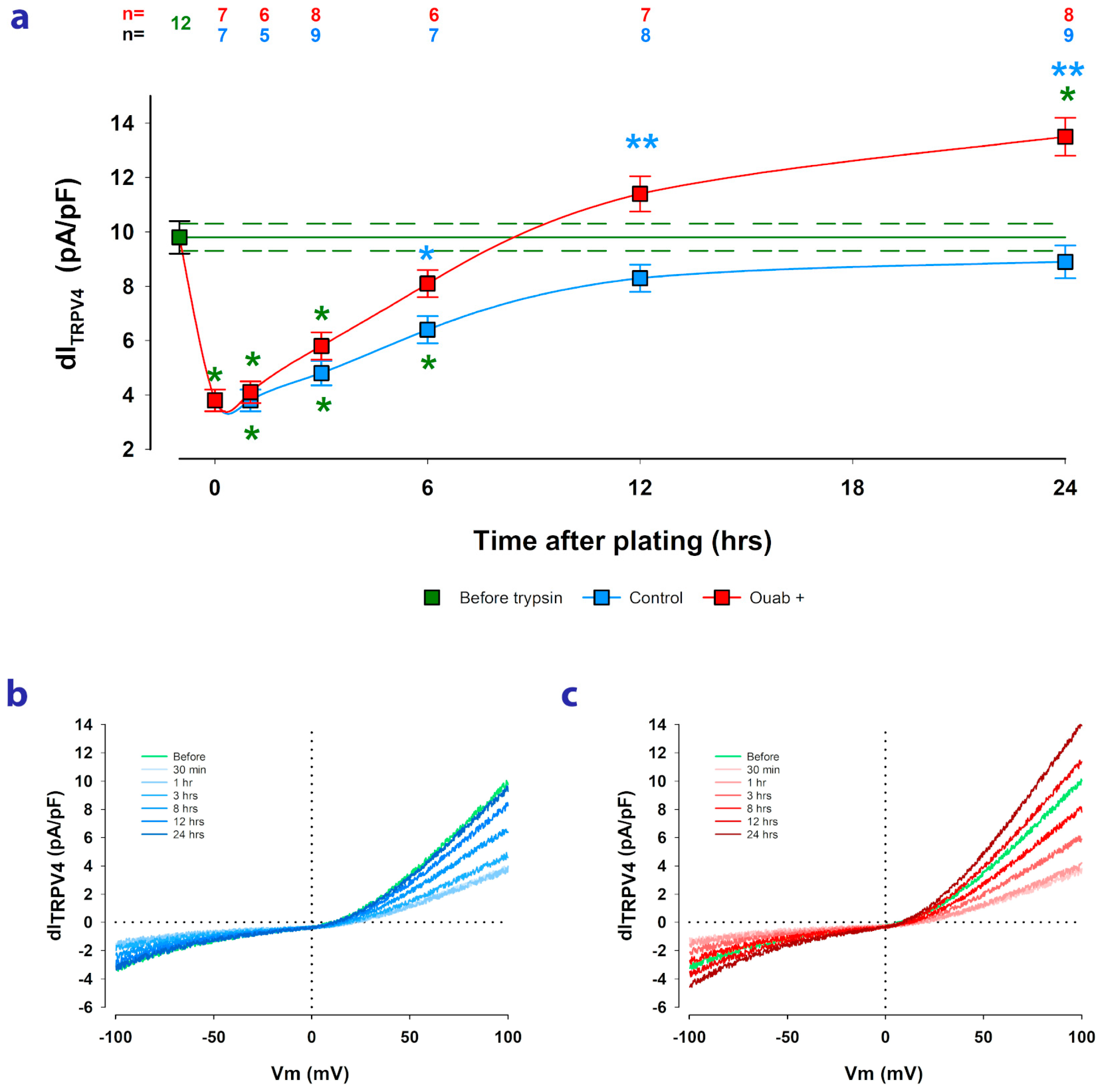
2.3. Ouabain Speeds up the Recovery of TRPV4 Channels Lost by Trypsinization
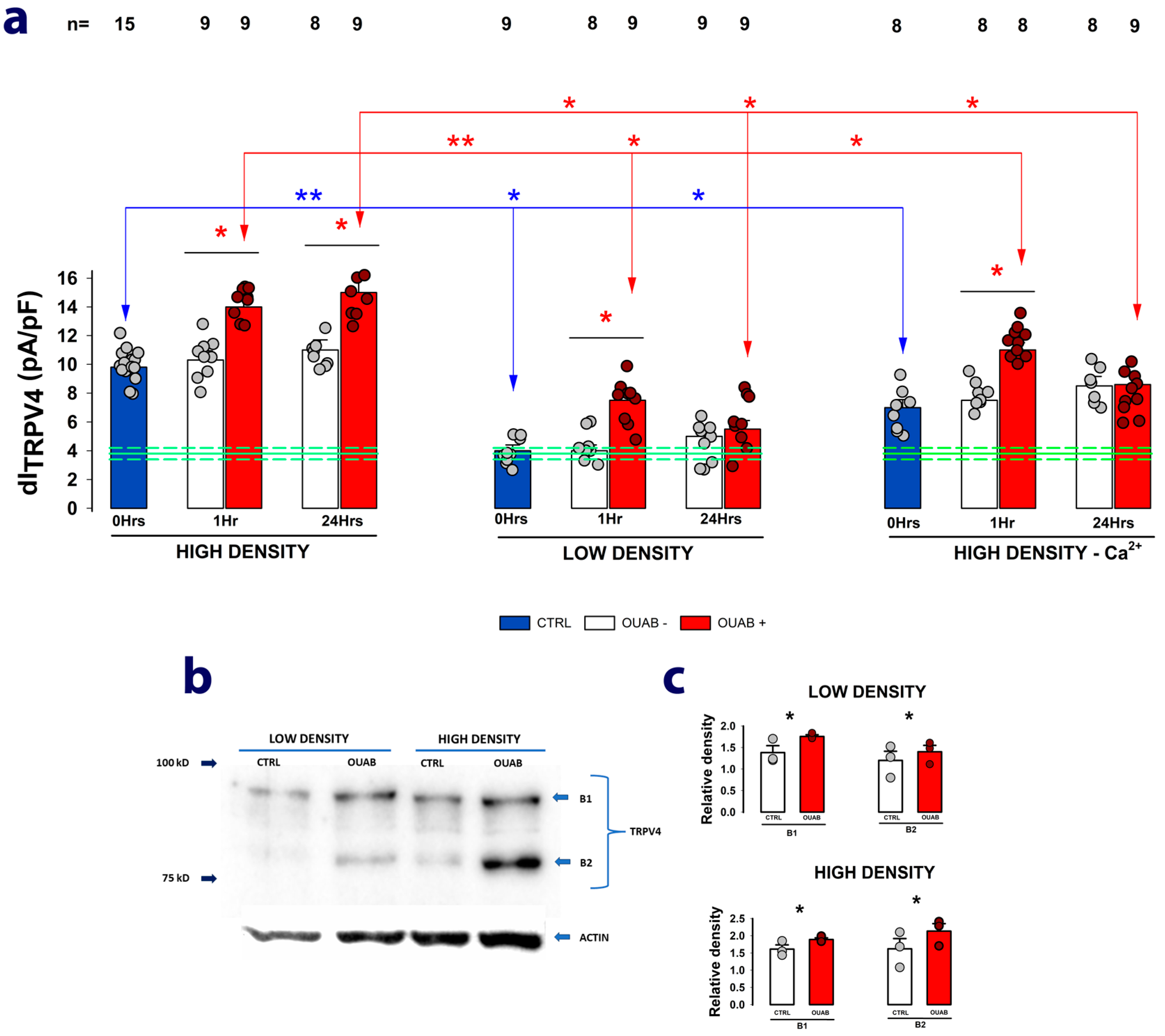
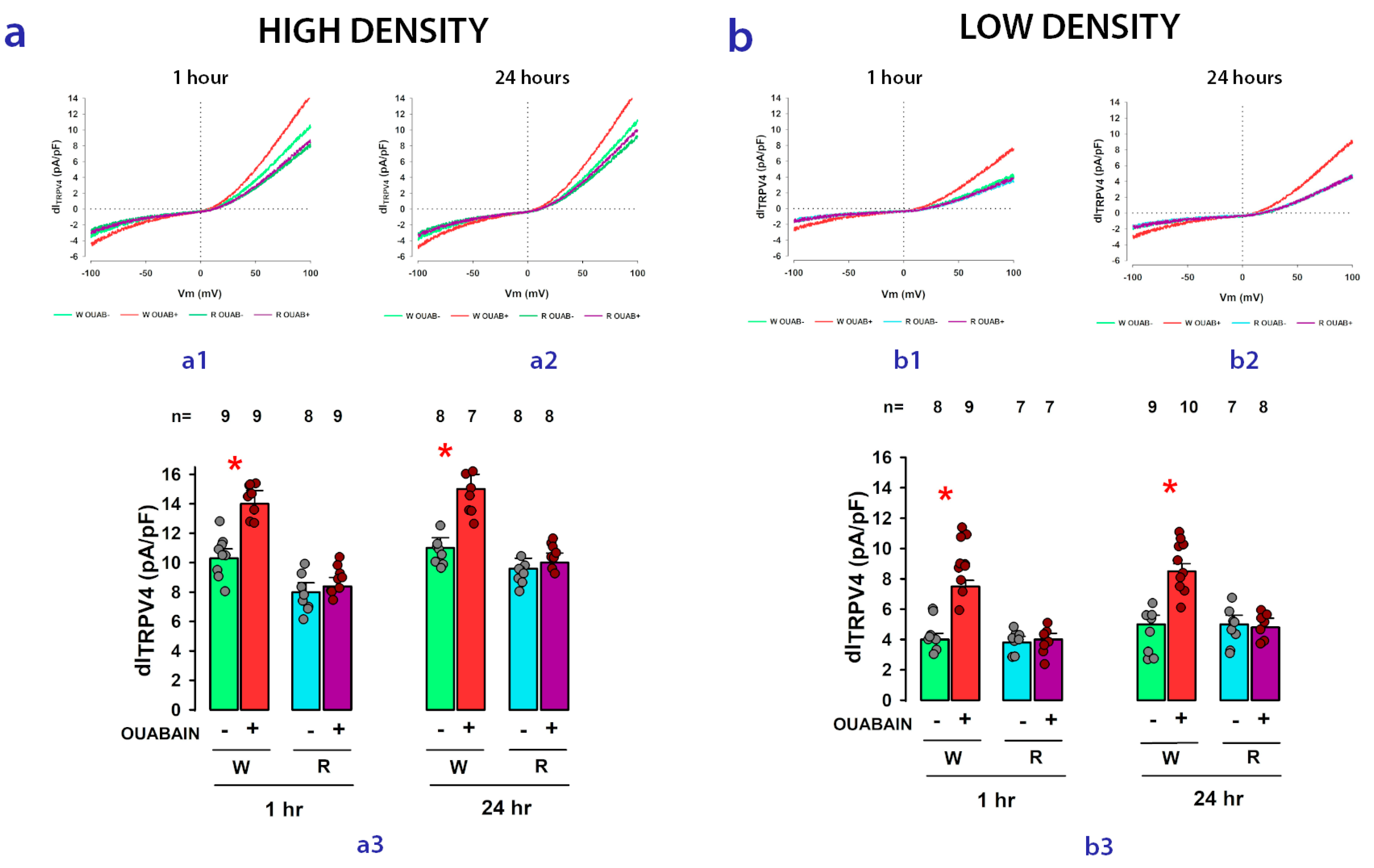
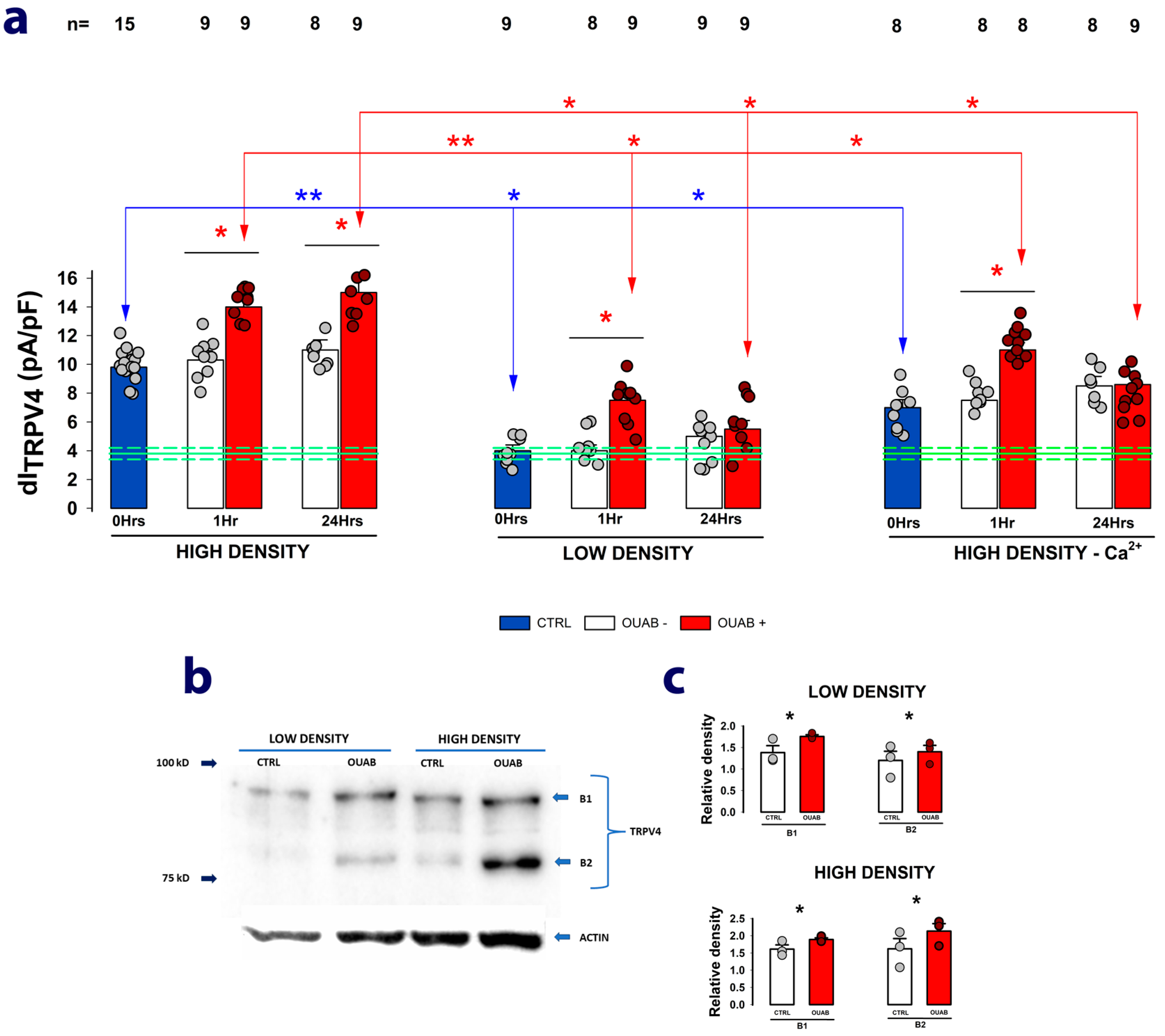
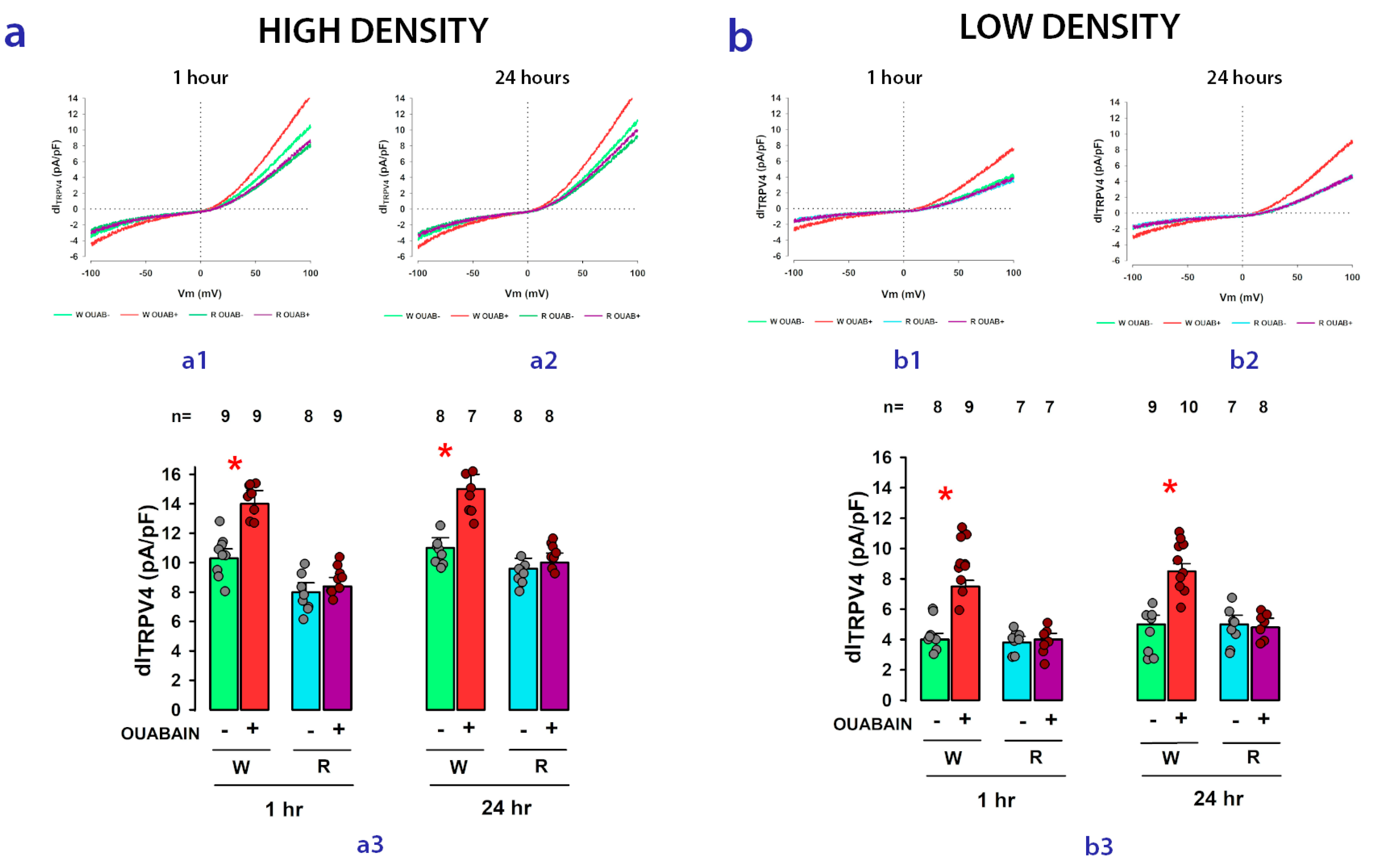
2.4. The Enhancing Effect of Ouabain on dITRPV4 Relies on Contact between Cells
2.5. Na+/K+-ATPase Acts as the Receptor for Ouabain-Mediated Modulation of TRPV4 Channel Activity and Expression
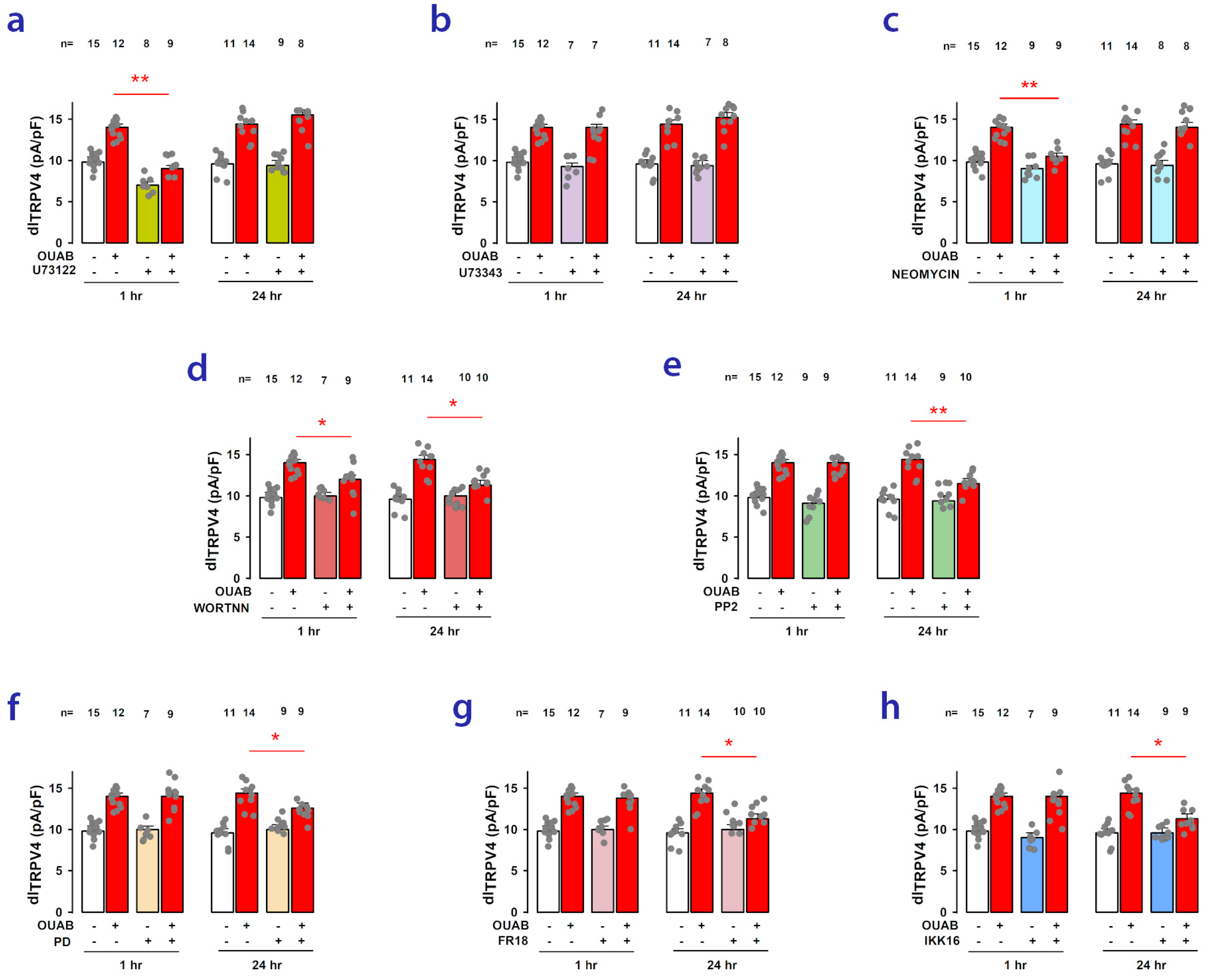
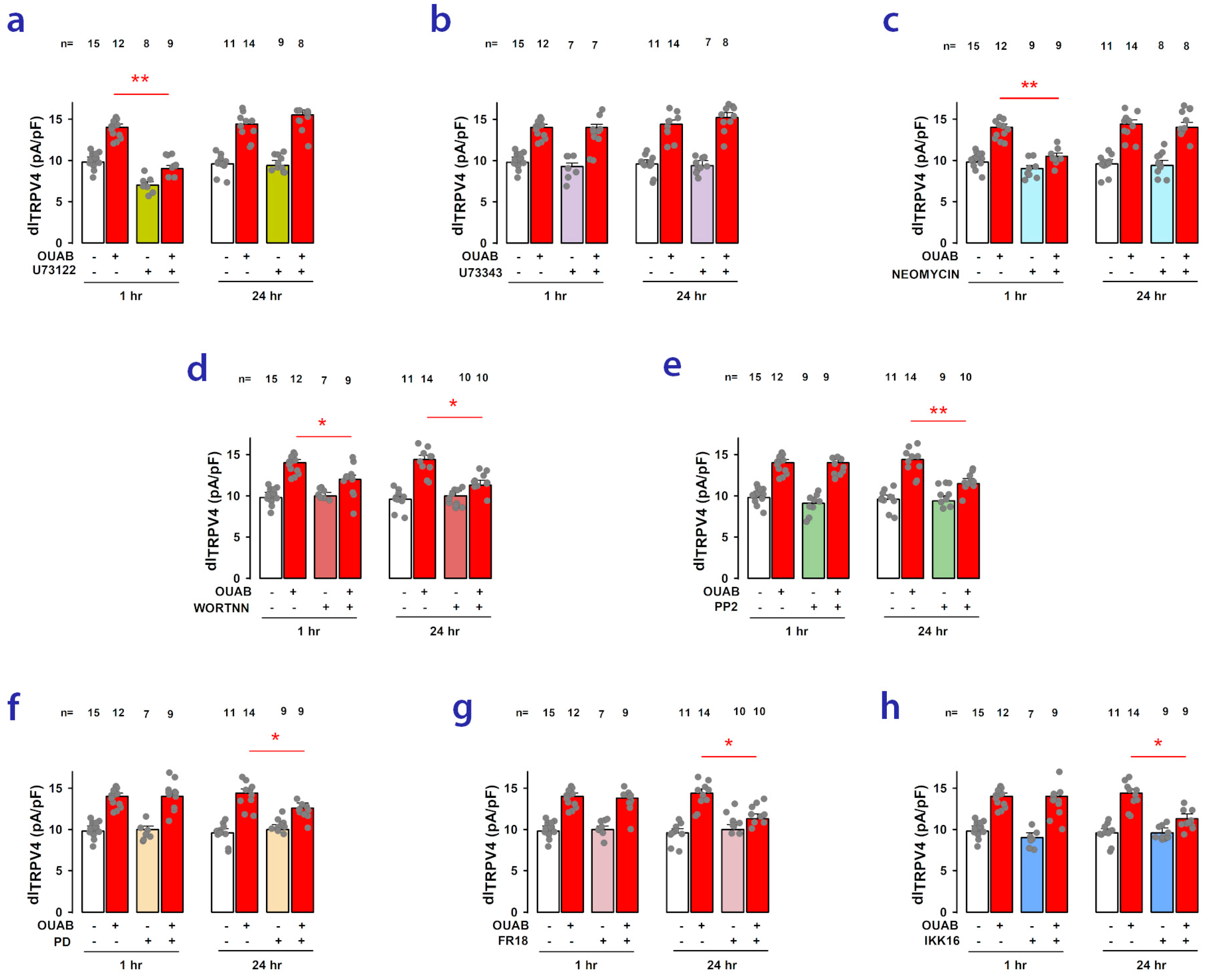
2.6. Signaling Pathways Involved in Short- and Long-Term Ouabain-Stimulated Enhancement of TRPV4 Activity
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Electrophysiological Recording of Cells
4.3. Measurement of Membrane Area
4.4. Western Blotting and Densitometry Analyses
4.5. Solutions
4.6. Chemicals and Drugs
4.7. Analysis of Data
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hollman, A. Plants and cardiac glycosides. Br. Heart J. 1985, 54, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Bejček, J.; Jurášek, M.; Spiwok, V.; Rimpelová, S. Quo vadis Cardiac Glycoside Research? Toxins 2021, 13, 344. [Google Scholar] [CrossRef] [PubMed]
- Botelho, A.F.M.; Pierezan, F.; Soto-Blanco, B.; Melo, M.M. A review of cardiac glycosides: Structure, toxicokinetics, clinical signs, diagnosis and antineoplastic potential. Toxicon 2018, 158, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Plant-derived cardiac glycosides: Role in heart ailments and cancer management. Biomed. Pharmacother. 2016, 84, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Fuerstenwerth, H. Ouabain—The key to cardioprotection? Am. J. Ther. 2014, 21, 395–402. [Google Scholar] [CrossRef]
- Zhang, H.; Sridhar Reddy, M.; Phoenix, S.; Deslongchamps, P. Total synthesis of ouabagenin and ouabain. Angew. Chem. Int. Ed. Engl. 2008, 47, 1272–1275. [Google Scholar] [CrossRef]
- Haustein, K.O. Therapeutic range of cardiac glycosides. Basic Res. Cardiol. 1984, 79, 147–153. [Google Scholar] [CrossRef]
- Schoner, W. Ouabain, a new steroid hormone of adrenal gland and hypothalamus. Exp. Clin. Endocrinol. Diabetes 2000, 108, 449–454. [Google Scholar] [CrossRef]
- Schoner, W. Endogenous cardiac glycosides, a new class of steroid hormones. Eur. J. Biochem. 2002, 269, 2440–2448. [Google Scholar] [CrossRef]
- Cho, M.J.; Thompson, D.P.; Cramer, C.T.; Vidmar, T.J.; Scieszka, J.F. The Madin Darby canine kidney (MDCK) epithelial cell monolayer as a model cellular transport barrier. Pharm. Res. 1989, 6, 71–77. [Google Scholar] [CrossRef]
- Simmons, N.L. Cultured monolayers of MDCK cells: A novel model system for the study of epithelial development and function. Gen. Pharmacol. 1982, 13, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Cereijido, M.; Robbins, E.S.; Dolan, W.J.; Rotunno, C.A.; Sabatini, D.D. Polarized monolayers formed by epithelial cells on a permeable and translucent support. J. Cell Biol. 1978, 77, 853–880. [Google Scholar] [CrossRef] [PubMed]
- Larre, I.; Lazaro, A.; Contreras, R.G.; Balda, M.S.; Matter, K.; Flores-Maldonado, C.; Ponce, A.; Flores-Benitez, D.; Rincon-Heredia, R.; Padilla-Benavides, T.; et al. Ouabain modulates epithelial cell tight junction. Proc. Natl. Acad. Sci. USA 2010, 107, 11387–11392. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.; Ortuño-Pineda, C.; Flores-Maldonado, C.; Larre, I.; Rendón, J.M.; Hinojosa, L.; Ponce, A.; Ogazón, A.; Serrano, M.; Valdes, J.; et al. Ouabain Modulates the Adherens Junction in Renal Epithelial Cells. Cell Physiol. Biochem. 2019, 52, 1381–1397. [Google Scholar] [PubMed]
- Ponce, A.; Larre, I.; Castillo, A.; Garcia-Villegas, R.; Romero, A.; Flores-Maldonado, C.; Martinez-Rendón, J.; Contreras, R.G.; Cereijido, M. Ouabain Increases Gap Junctional Communication in Epithelial Cells. Cell Physiol. Biochem. 2014, 34, 2081–2090. [Google Scholar] [CrossRef]
- Ponce, A.; Larre, I.; Castillo, A.; Flores-Maldonado, C.; Verdejo-Torres, O.; Contreras, R.G.; Cereijido, M. Ouabain Modulates the Distribution of Connexin 43 in Epithelial Cells. Cell Physiol. Biochem. 2016, 39, 1329–1338. [Google Scholar] [CrossRef]
- Larre, I.; Castillo, A.; Flores-Maldonado, C.; Contreras, R.G.; Galvan, I.; Muñoz-Estrada, J.; Cereijido, M. Ouabain modulates ciliogenesis in epithelial cells. Proc. Natl. Acad. Sci. USA 2011, 108, 20591–20596. [Google Scholar] [CrossRef]
- Cereijido, M.; Jimenez, L.; Hinojosa, L.; Castillo, A.; Martínez-Rendon, J.; Ponce, A. Ouabain-Induced Changes in the Expression of Voltage-Gated Potassium Channels in Epithelial Cells Depend on Cell-Cell Contacts. Int. J. Mol. Sci. 2022, 23, 13257. [Google Scholar] [CrossRef]
- Mukaiyama, M.; Yamasaki, Y.; Usui, T.; Nagumo, Y. Transient receptor potential V4 channel stimulation induces reversible epithelial cell permeability in MDCK cell monolayers. FEBS Lett. 2019, 593, 2250–2260. [Google Scholar] [CrossRef]
- Espadas-Álvarez, H.; Martínez-Rendón, J.; Larre, I.; Matamoros-Volante, A.; Romero-García, T.; Rosenbaum, T.; Rueda, A.; García-Villegas, R. TRPV4 activity regulates nuclear Ca2+ and transcriptional functions of β-catenin in a renal epithelial cell model. J. Cell Physiol. 2021, 236, 3599–3614. [Google Scholar] [CrossRef]
- Plant, T.D.; Strotmann, R. TRPV4. In Handbook of Experimental Pharmacology; Springer: New York, NY, USA, 2007; Volume 179, pp. 189–205. [Google Scholar] [CrossRef]
- Becker, D.; Blase, C.; Bereiter-Hahn, J.; Jendrach, M. TRPV4 exhibits a functional role in cell-volume regulation. J. Cell Sci. 2005, 118 Pt 11, 2435–2440. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Bereiter-Hahn, J.; Jendrach, M. Functional interaction of the cation channel transient receptor potential vanilloid 4 (TRPV4) and actin in volume regulation. Eur. J. Cell Biol. 2009, 88, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W. TRPV4 plays an evolutionary conserved role in the transduction of osmotic and mechanical stimuli in live animals. J. Physiol. 2005, 567 Pt 1, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Watanabe, H.; Vriens, J. The TRPV4 channel: Structure-function relationship and promiscuous gating behaviour. Pflugers. Arch. 2003, 446, 298–303. [Google Scholar] [CrossRef]
- Thorneloe, K.S.; Sulpizio, A.C.; Lin, Z.; Figueroa, D.J.; Clouse, A.K.; McCafferty, G.P.; Chendrimada, T.P.; Lashinger, E.S.; Gordon, E.; Evans, L.; et al. N-((1S)-1-{[4-((2S)-2-{[(2,4-dichlorophenyl)sulfonyl]amino}-3-hydroxypropanoyl)-1-piperazinyl]carbonyl}-3-methylbutyl)-1-benzothiophene-2-carboxamide (GSK1016790A), a novel and potent transient receptor potential vanilloid 4 channel agonist induces urinary bladder contraction and hyperactivity: Part I. J. Pharmacol. Exp. Ther. 2008, 326, 432–442. [Google Scholar]
- Willette, R.N.; Bao, W.; Nerurkar, S.; Yue, T.L.; Doe, C.P.; Stankus, G.; Turner, G.H.; Ju, H.; Thomas, H.; Fishman, C.E.; et al. Systemic activation of the transient receptor potential vanilloid subtype 4 channel causes endothelial failure and circulatory collapse: Part 2. J. Pharmacol. Exp. Ther. 2008, 326, 443–452, Erratum in: J. Pharmacol. Exp. Ther. 2011, 338, 408–409. [Google Scholar] [CrossRef]
- Nilius, B.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T. TRPV4 calcium entry channel: A paradigm for gating diversity. Am. J. Physiol. Cell Physiol. 2004, 286, C195–C205. [Google Scholar] [CrossRef]
- Everaerts, W.; Zhen, X.; Ghosh, D.; Vriens, J.; Gevaert, T.; Gilbert, J.P.; Hayward, N.J.; McNamara, C.R.; Xue, F.; Moran, M.M.; et al. Inhibition of the cation channel TRPV4 improves bladder function in mice and rats with cyclophosphamide-induced cystitis. Proc. Natl. Acad. Sci. USA 2010, 107, 19084–19089. [Google Scholar] [CrossRef]
- Merrill, L.; Vizzard, M.A. Intravesical TRPV4 blockade reduces repeated variate stress-induced bladder dysfunction by increasing bladder capacity and decreasing voiding frequency in male rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R471–R480. [Google Scholar] [CrossRef]
- Gaush, C.R.; Hard, W.L.; Smith, T.F. Characterization of an Established Line of Canine Kidney Cells (MDCK). Proc. Soc. Exp. Biol. Med. 1966, 122, 931–935. [Google Scholar] [CrossRef]
- Rindler, M.J.; Chuman, L.M.; Shaffer, L.; Saier, M.H., Jr. Retention of differentiated properties in an established dog kidney epithelial cell line (MDCK). J. Cell Biol. 1979, 81, 635–648. [Google Scholar] [CrossRef]
- Lever, J.E. Expression of differentiated functions in kidney epithelial cell lines. Miner. Electrolyte Metab. 1986, 12, 14–19. [Google Scholar] [PubMed]
- Taub, M.; Saier, M.H., Jr. An established but differentiated kidney epithelial cell line (MDCK). Methods Enzymol. 1979, 58, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Ponce, A.; Bolivar, J.J.; Vega, J.; Cereijido, M. Synthesis of plasma membrane and potassium channels in epithelial (MDCK) cells. Cell. Physiol. Biochem. 1991, 1, 195–204. [Google Scholar] [CrossRef]
- Talavera, D.; Ponce, A.; Fiorentino, R.; González-Mariscal, L.; Contreras, R.G.; Sánchez, S.H.; García-Villegas, M.R.; Valdés, J.; Cereijido, M. Expression of potassium channels in epithelial cells depends on calcium-activated cell-cell contacts. J. Membr. Biol. 1995, 143, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Boulan, E.; Macara, I.G. Organization and execution of the epithelial polarity programme. Nat. Rev. Mol. Cell Biol. 2014, 15, 225–242. [Google Scholar] [CrossRef]
- Cereijido, M.; Contreras, R.G.; Shoshani, L. Cell adhesion, polarity, and epithelia in the dawn of metazoans. Physiol. Rev. 2004, 84, 1229–1262. [Google Scholar] [CrossRef]
- Cereijido, M.; Contreras, R.G.; Shoshani, L.; Flores-Benitez, D.; Larre, I. Tight junction and polarity interaction in the transporting epithelial phenotype. Biochim. Biophys. Acta. 2008, 1778, 770–793. [Google Scholar] [CrossRef]
- Willenborg, C.; Prekeris, R. Apical protein transport and lumen morphogenesis in polarized epithelial cells. Biosci. Rep. 2011, 31, 245–256. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, L.; Contreras, R.G.; Bolívar, J.J.; Ponce, A.; Chávez De Ramirez, B.; Cereijido, M. Role of calcium in tight junction formation between epithelial cells. Am. J. Physiol. 1990, 259 Pt 1, C978–C986. [Google Scholar] [CrossRef]
- Contreras, R.G.; Miller, J.H.; Zamora, M.; González-Mariscal, L.; Cereijido, M. Interaction of calcium with plasma membrane of epithelial (MDCK) cells during junction formation. Am. J Physiol. 1992, 263 Pt 1, C313–C318. [Google Scholar] [CrossRef] [PubMed]
- Soderberg, K.; Rossi, B.; Lazdunski, M.; Louvard, D. Characterization of ouabain-resistant mutants of a canine kidney cell line, MDCK. J. Biol. Chem. 1983, 258, 12300–12307. [Google Scholar] [CrossRef] [PubMed]
- Canessa, C.M.; Horisberger, J.D.; Rossier, B.C. Mutation of a tyrosine in the H3-H4 ectodomain of Na,K-ATPase alpha subunit confers ouabain resistance. J. Biol. Chem. 1993, 268, 17722–17726. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Mohammadi, K.; Aynafshar, B.; Wang, H.; Li, D.; Liu, J.; Ivanov, A.V.; Xie, Z.; Askari, A. Role of caveolae in signal-transducing function of cardiac Na+/K+-ATPase. Am. J. Physiol. Cell Physiol. 2003, 284, C1550–C1560. [Google Scholar] [CrossRef] [PubMed]
- Juhaszova, M.; Blaustein, M.P. Na+ pump low and high ouabain affinity alpha subunit isoforms are differently distributed in cells. Proc. Natl. Acad. Sci. USA 1997, 94, 1800–1805. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z. Molecular mechanisms of Na/K-ATPase-mediated signal transduction. Ann. N. Y. Acad. Sci. 2003, 986, 497–503. [Google Scholar] [CrossRef]
- Kadamur, G.; Ross, E.M. Mammalian phospholipase C. Annu. Rev. Physiol. 2013, 75, 127–154. [Google Scholar] [CrossRef]
- Gresset, A.; Sondek, J.; Harden, T.K. The phospholipase C isozymes and their regulation. Subcell Biochem. 2012, 58, 61–94. [Google Scholar] [CrossRef]
- Harraz, O.F.; Longden, T.A.; Hill-Eubanks, D.; Nelson, M.T. PIP2 depletion promotes TRPV4 channel activity in mouse brain capillary endothelial cells. Elife 2018, 7, e38689. [Google Scholar] [CrossRef]
- Bleasdale, J.E.; Thakur, N.R.; Gremban, R.S.; Bundy, G.L.; Fitzpatrick, F.A.; Smith, R.J.; Bunting, S.T. Selective inhibition of receptor-coupled phospholipase C-dependent processes in human platelets and polymorphonuclear neutrophils. J. Pharmacol. Exp. Ther. 1990, 255, 756–768. [Google Scholar]
- Liscovitch, M.; Chalifa, V.; Danin, M.; Eli, Y. Inhibition of neural phospholipase D activity by aminoglycoside antibiotics. Biochem. J. 1991, 279, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, J.P.; Plant, T.D.; Meves, H. The effects of bradykinin on K+ currents in NG108-15 cells treated with U73122, a phospholipase C inhibitor, or neomycin. Br. J. Pharmacol. 1997, 120, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.J.; Sam, L.M.; Justen, J.M.; Bundy, G.L.; Bala, G.A.; Bleasdale, J.E. Receptor-coupled signal transduction in human polymorphonuclear neutrophils: Effects of a novel inhibitor of phospholipase C-dependent processes on cell responsiveness. J. Pharmacol. Exp. Ther. 1990, 253, 688–697. [Google Scholar] [PubMed]
- Fruman, D.A.; Meyers, R.E.; Cantley, L.C. Phosphoinositide kinases. Annu. Rev. Biochem. 1998, 67, 481–507. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shreder, K.R.; Gai, W.; Corral, S.; Ferris, D.K.; Rosenblum, J.S. Wortmannin, a widely used phosphoinositide 3-kinase inhibitor, also potently inhibits mammalian polo-like kinase. Chem. Biol. 2005, 12, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Kapeller, R.; Cantley, L.C. Phosphatidylinositol 3-kinase. Bioessays 1994, 16, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. Phosphatidylinositol 4-kinases and the role of polyphosphoinositides in cellular regulation. Endocr. Rev. 1992, 13, 692–706. [Google Scholar] [CrossRef]
- Hanke, J.H.; Gardner, J.P.; Dow, R.L.; Changelian, P.S.; Brissette, W.H.; Weringer, E.J.; Pollok, B.A.; Connelly, P.A. Discovery of a novel, potent, and Src family-selective tyrosine kinase inhibitor. Study of Lck- and FynT-dependent T cell activation. J. Biol. Chem. 1996, 271, 695–701. [Google Scholar] [CrossRef]
- Chong, Y.P.; Ia, K.K.; Mulhern, T.D.; Cheng, H.C. Endogenous and synthetic inhibitors of the Src-family protein tyrosine kinases. Biochim. Biophys. Acta 2005, 1754, 210–220. [Google Scholar] [CrossRef]
- Barrett, S.D.; Bridges, A.J.; Dudley, D.T.; Saltiel, A.R.; Fergus, J.H.; Flamme, C.M.; Delaney, A.M.; Kaufman, M.; LePage, S.; Leopold, W.R.; et al. The discovery of the benzhydroxamate MEK inhibitors CI-1040 and PD 0325901. Bioorg. Med. Chem. Lett. 2008, 18, 6501–6504. [Google Scholar] [CrossRef]
- Ohori, M.; Takeuchi, M.; Maruki, R.; Nakajima, H.; Miyake, H. FR180204, a novel and selective inhibitor of extracellular signal-regulated kinase, ameliorates collagen-induced arthritis in mice. Naunyn Schmiedebergs Arch. Pharmacol. 2007, 374, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Ohori, M.; Kinoshita, T.; Okubo, M.; Sato, K.; Yamazaki, A.; Arakawa, H.; Nishimura, S.; Inamura, N.; Nakajima, H.; Neya, M.; et al. Identification of a selective ERK inhibitor and structural determination of the inhibitor-ERK2 complex. Biochem. Biophys. Res. Commun. 2005, 336, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.R. The NF-kappaB regulatory network. Cardiovasc. Toxicol. 2006, 6, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Häcker, H.; Karin, M. Regulation and function of IKK and IKK-related kinases. Sci. STKE 2006, 2006, re13. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed]
- Waelchli, R.; Bollbuck, B.; Bruns, C.; Buhl, T.; Eder, J.; Feifel, R.; Hersperger, R.; Janser, P.; Revesz, L.; Zerwes, H.G.; et al. Design and preparation of 2-benzamido-pyrimidines as inhibitors of IKK. Bioorg. Med. Chem. Lett. 2006, 16, 108–112. [Google Scholar] [CrossRef]
- Coldewey, S.M.; Rogazzo, M.; Collino, M.; Patel, N.S.; Thiemermann, C. Inhibition of IᴋB kinase reduces the multiple organ dysfunction caused by sepsis in the mouse. Dis. Model. Mech. 2013, 6, 1031–1042. [Google Scholar]
- Deepak, D.; Srivastava, S.; Khare, N.K.; Khare, A. Cardiac glycosides. Fortschr. Chem. Org. Naturst. 1996, 69, 71–155. [Google Scholar] [CrossRef]
- Hauptman, P.J.; Garg, R.; Kelly, R.A. Cardiac glycosides in the next millennium. Prog. Cardiovasc. Dis. 1999, 41, 247–254. [Google Scholar] [CrossRef]
- Pierre, S.V.; Xie, Z. The Na,K-ATPase receptor complex: Its organization and membership. Cell Biochem. Biophys. 2006, 46, 303–316. [Google Scholar] [CrossRef]
- Cereijido, M.; Contreras, R.G.; Shoshani, L.; Larre, I. The Na+-K+-ATPase as self-adhesion molecule and hormone receptor. Am. J. Physiol. Cell Physiol. 2012, 302, C473–C481. [Google Scholar] [CrossRef] [PubMed]
- Aperia, A.; Akkuratov, E.E.; Fontana, J.M.; Brismar, H. Na+-K+-ATPase, a new class of plasma membrane receptors. Am. J. Physiol. Cell Physiol. 2016, 310, C491–C495. [Google Scholar] [CrossRef] [PubMed]
- Bejček, J.; Spiwok, V.; Kmoníčková, E.; Rimpelová, S. Na+/K+-ATPase Revisited: On Its Mechanism of Action, Role in Cancer, and Activity Modulation. Molecules 2021, 26, 1905. [Google Scholar] [CrossRef] [PubMed]
- Alevizopoulos, K.; Calogeropoulou, T.; Lang, F.; Stournaras, C. Na+/K+ ATPase inhibitors in cancer. Curr. Drug Targets 2014, 15, 988–1000. [Google Scholar] [CrossRef]
- Mijatovic, T.; Dufrasne, F.; Kiss, R. Na+/K+-ATPase and cancer. Pharm. Pat. Anal. 2012, 1, 91–106. [Google Scholar] [CrossRef]
- Prassas, I.; Diamandis, E.P. Novel therapeutic applications of cardiac glycosides. Nat. Rev. Drug Discov. 2008, 7, 926–935. [Google Scholar] [CrossRef]
- Diederich, M.; Muller, F.; Cerella, C. Cardiac glycosides: From molecular targets to immunogenic cell death. Biochem. Pharmacol. 2017, 125, 1–11. [Google Scholar] [CrossRef]
- Harwood, S.; Yaqoob, M.M. Ouabain-induced cell signaling. Front. Biosci. 2005, 10, 2011–2017. [Google Scholar] [CrossRef]
- Xie, Z. Ouabain interaction with cardiac Na/K-ATPase reveals that the enzyme can act as a pump and as a signal transducer. Cell Mol. Biol. 2001, 47, 383–390. [Google Scholar]
- Fontana, J.M.; Burlaka, I.; Khodus, G.; Brismar, H.; Aperia, A. Calcium oscillations triggered by cardiotonic steroids. FEBS J. 2013, 280, 5450–5455. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T. Diversity of TRP channel activation. Novartis Found. Symp. 2004, 258, 140–149, discussion 149–159, 263–266. [Google Scholar] [PubMed]
- Montell, C. The TRP superfamily of cation channels. Sci. STKE 2005, 2005, re3. [Google Scholar] [CrossRef] [PubMed]
- Toft-Bertelsen, T.L.; MacAulay, N. TRPing to the Point of Clarity: Understanding the Function of the Complex TRPV4 Ion Channel. Cells 2021, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, I.M.; Liedtke, W.; Sanderson, M.J.; Valverde, M.A. TRPV4 channel participates in receptor-operated calcium entry and ciliary beat frequency regulation in mouse airway epithelial cells. Proc. Natl. Acad. Sci. USA 2008, 105, 12611–12616. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Yang, H.; Mergler, S.; Liu, H.; Tachado, S.D.; Zhang, F.; Kao, W.W.Y.; Koziel, H.; Pleyer, U.; Reinach, P. Dependence of regulatory volume decrease on transient receptor potential vanilloid 4 (TRPV4) expression in human corneal epithelial cells. Cell Calcium. 2008, 44, 374–385. [Google Scholar] [CrossRef]
- Tian, W.; Salanova, M.; Xu, H.; Lindsley, J.N.; Oyama, T.T.; Anderson, S.; Bachmann, S.; Cohen, D.M. Renal expression of osmotically responsive cation channel TRPV4 is restricted to water-impermeant nephron segments. Am. J. Physiol. Physiol. 2004, 287, F17–F24. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; Guo, H.; Wang, Y. Na+/K+-ATPase-mediated signal transduction and Na+/K+-ATPase regulation. Fundam. Clin. Pharmacol. 2008, 22, 615–621. [Google Scholar] [CrossRef]
- Xie, Z.; Cai, T. Na+-K+--ATPase-mediated signal transduction: From protein interaction to cellular function. Mol. Interv. 2003, 3, 157–168. [Google Scholar] [CrossRef]
- Plakhova, V.B.; Penniyaynen, V.A.; Rogachevskii, I.V.; Podzorova, S.A.; Khalisov, M.M.; Ankudinov, A.V.; Krylov, B.V. Dual mechanism of modulation of NaV1.8 sodium channels by ouabain. Can. J. Physiol. Pharmacol. 2020, 98, 785–802. [Google Scholar] [CrossRef]
- Valentich, J.D. Morphological similarities between the dog kidney cell line MDCK and the mammalian cortical collecting tubule. Ann. N. Y. Acad. Sci. 1981, 372, 384–405. [Google Scholar] [CrossRef]
- Subramanya, A.R.; Ellison, D.H. Distal convoluted tubule. Clin. J. Am. Soc. Nephrol. 2014, 9, 2147–2163. [Google Scholar] [CrossRef] [PubMed]
- Pochynyuk, O.; Zaika, O.; O’Neil, R.G.; Mamenko, M. Novel insights into TRPV4 function in the kidney. Pflugers. Arch. 2013, 465, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Stavniichuk, A.; Pyrshev, K.; Zaika, O.; Tomilin, V.N.; Kordysh, M.; Lakk, M.; Križaj, D.; Pochynyuk, O. TRPV4 expression in the renal tubule is necessary for maintaining whole body K+ homeostasis. Am. J. Physiol. Renal. Physiol. 2023, 324, F603–F616. [Google Scholar] [CrossRef] [PubMed]
- Mamenko, M.V.; Boukelmoune, N.; Tomilin, V.N.; Zaika, O.L.; Jensen, V.B.; O’Neil, R.G.; Pochynyuk, O.M. The renal TRPV4 channel is essential for adaptation to increased dietary potassium. Kidney Int. 2017, 91, 1398–1409. [Google Scholar] [CrossRef]
- Mamenko, M.; Zaika, O.; Boukelmoune, N.; O’Neil, R.G.; Pochynyuk, O. Deciphering physiological role of the mechanosensitive TRPV4 channel in the distal nephron. Am. J. Physiol. Renal Physiol. 2015, 308, F275–F286. [Google Scholar] [CrossRef]
- Conforti, L. Patch-Clamp Techniques. In Cell Physiology Source Book, 4th ed.; Sperelakis, N., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 369–381. ISBN 9780123877383. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Cat. No. | Stock mg/mL | Dose |
|---|---|---|---|
| GSK1016790A | G0798 | 15 | 20 nM |
| HC-067047 | SML0143 | 15 | 150 nM |
| U73122 | U6756 | 0.5 | 2 µM |
| U73343 | U6881 | 0.5 | 2 µM |
| Neomycin | 1405-10-3 | 10 | 50 µM |
| Wortmannin | W1628 | 14 | 10 nM |
| PP2 | P0042 | 1.4 | 100 nM |
| PD 098,059 | P215 | 30 | 7 µM |
| FR180204 | SML0320 | 25 | 400 nM |
| IKK-16 | SML1138 | 10 | 200 nM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponce, A.; Larre, I.; Jimenez, L.; Roldán, M.L.; Shoshani, L.; Cereijido, M. Ouabain’s Influence on TRPV4 Channels of Epithelial Cells: An Exploration of TRPV4 Activity, Expression, and Signaling Pathways. Int. J. Mol. Sci. 2023, 24, 16687. https://doi.org/10.3390/ijms242316687
Ponce A, Larre I, Jimenez L, Roldán ML, Shoshani L, Cereijido M. Ouabain’s Influence on TRPV4 Channels of Epithelial Cells: An Exploration of TRPV4 Activity, Expression, and Signaling Pathways. International Journal of Molecular Sciences. 2023; 24(23):16687. https://doi.org/10.3390/ijms242316687
Chicago/Turabian StylePonce, Arturo, Isabel Larre, Lidia Jimenez, Maria Luisa Roldán, Liora Shoshani, and Marcelino Cereijido. 2023. "Ouabain’s Influence on TRPV4 Channels of Epithelial Cells: An Exploration of TRPV4 Activity, Expression, and Signaling Pathways" International Journal of Molecular Sciences 24, no. 23: 16687. https://doi.org/10.3390/ijms242316687
APA StylePonce, A., Larre, I., Jimenez, L., Roldán, M. L., Shoshani, L., & Cereijido, M. (2023). Ouabain’s Influence on TRPV4 Channels of Epithelial Cells: An Exploration of TRPV4 Activity, Expression, and Signaling Pathways. International Journal of Molecular Sciences, 24(23), 16687. https://doi.org/10.3390/ijms242316687