
Knockdown of Simulated-Solar-Radiation-Sensitive miR-205-5p Does Not Induce Progression of Cutaneous Squamous Cell Carcinoma In Vitro

 , , and
, , and
Abstract
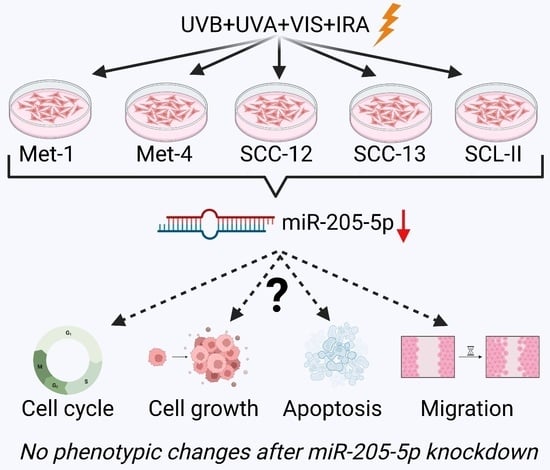
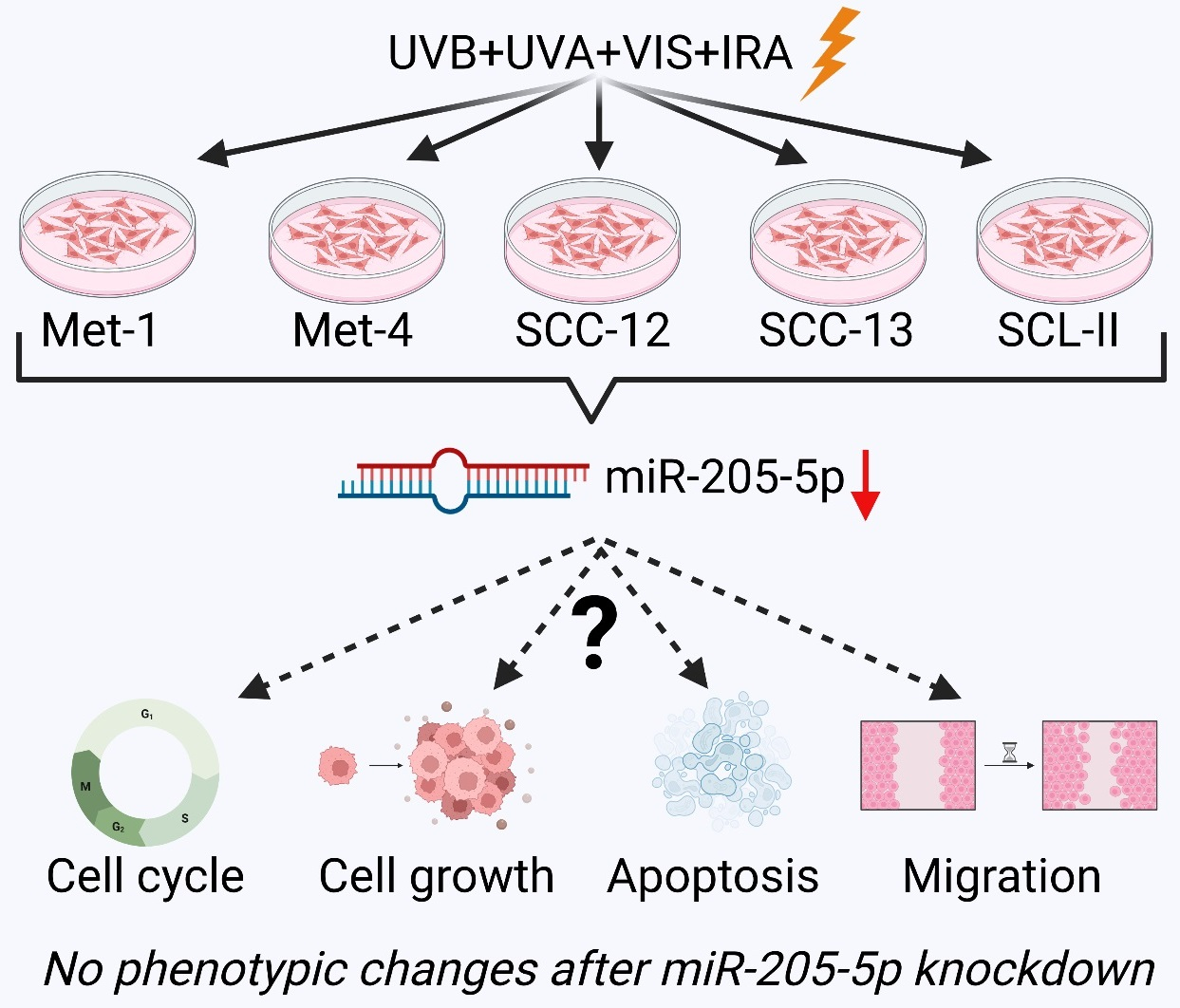
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
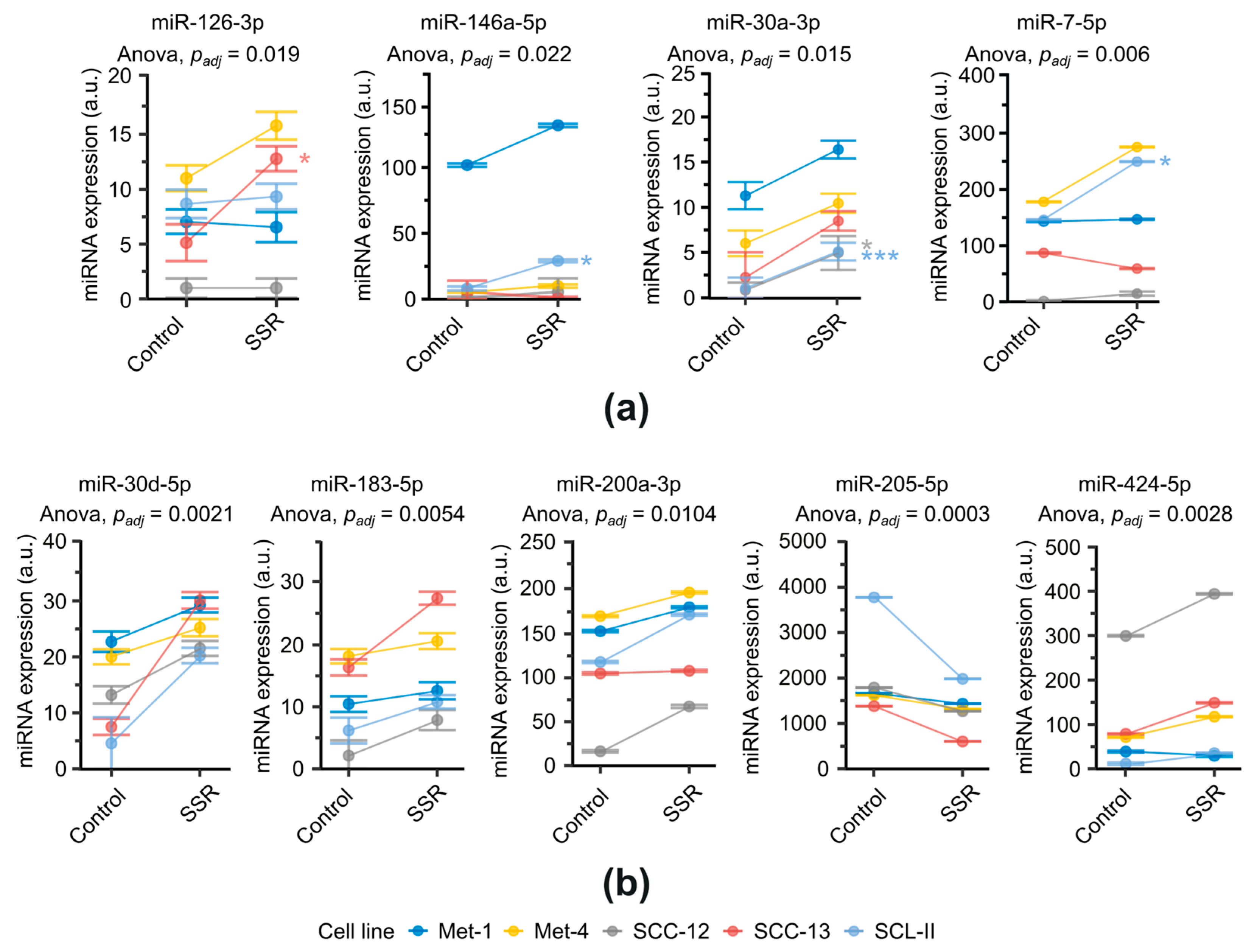
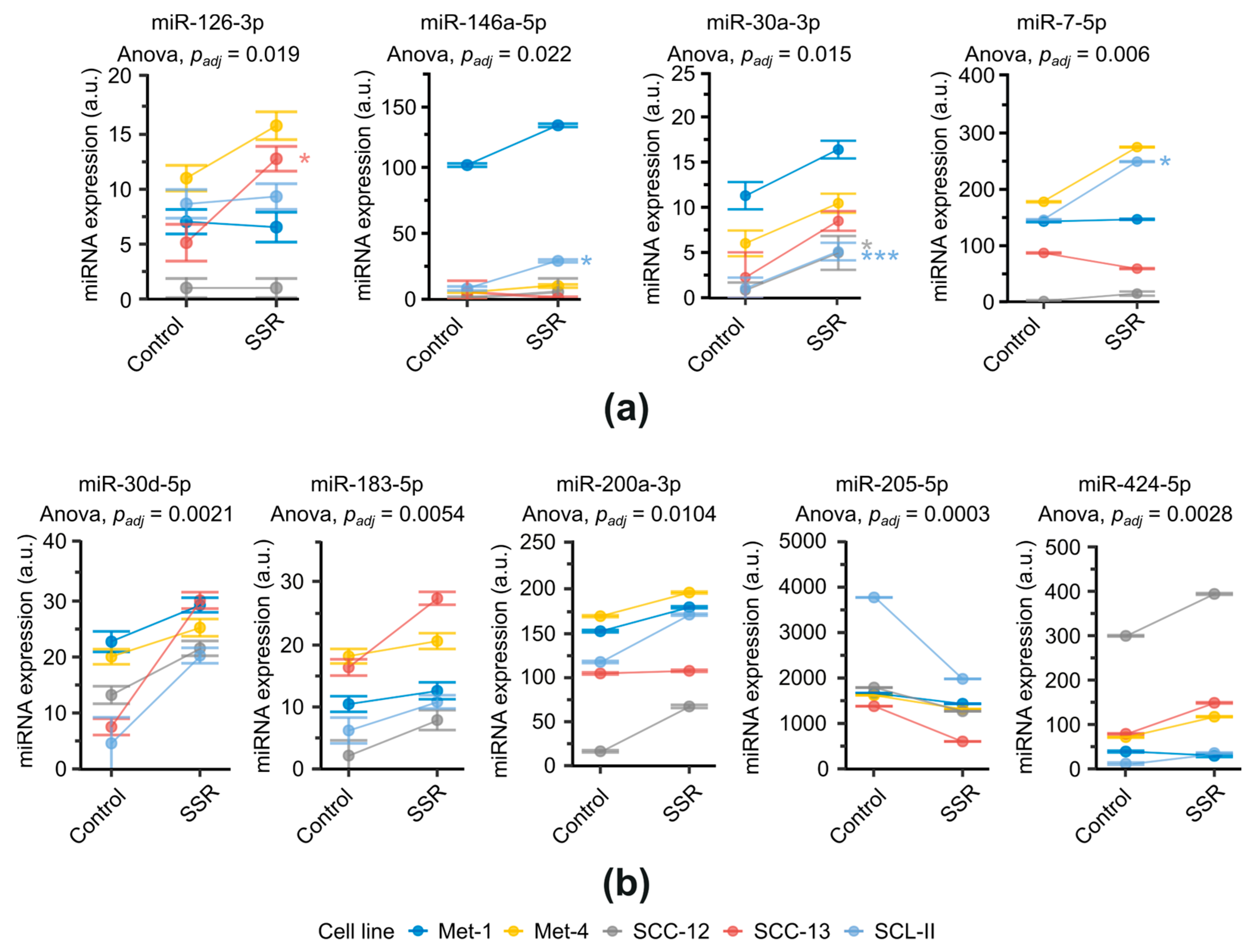
2.1. Differential miRNA Expression of cSCC Cell Lines after SSR-Exposure
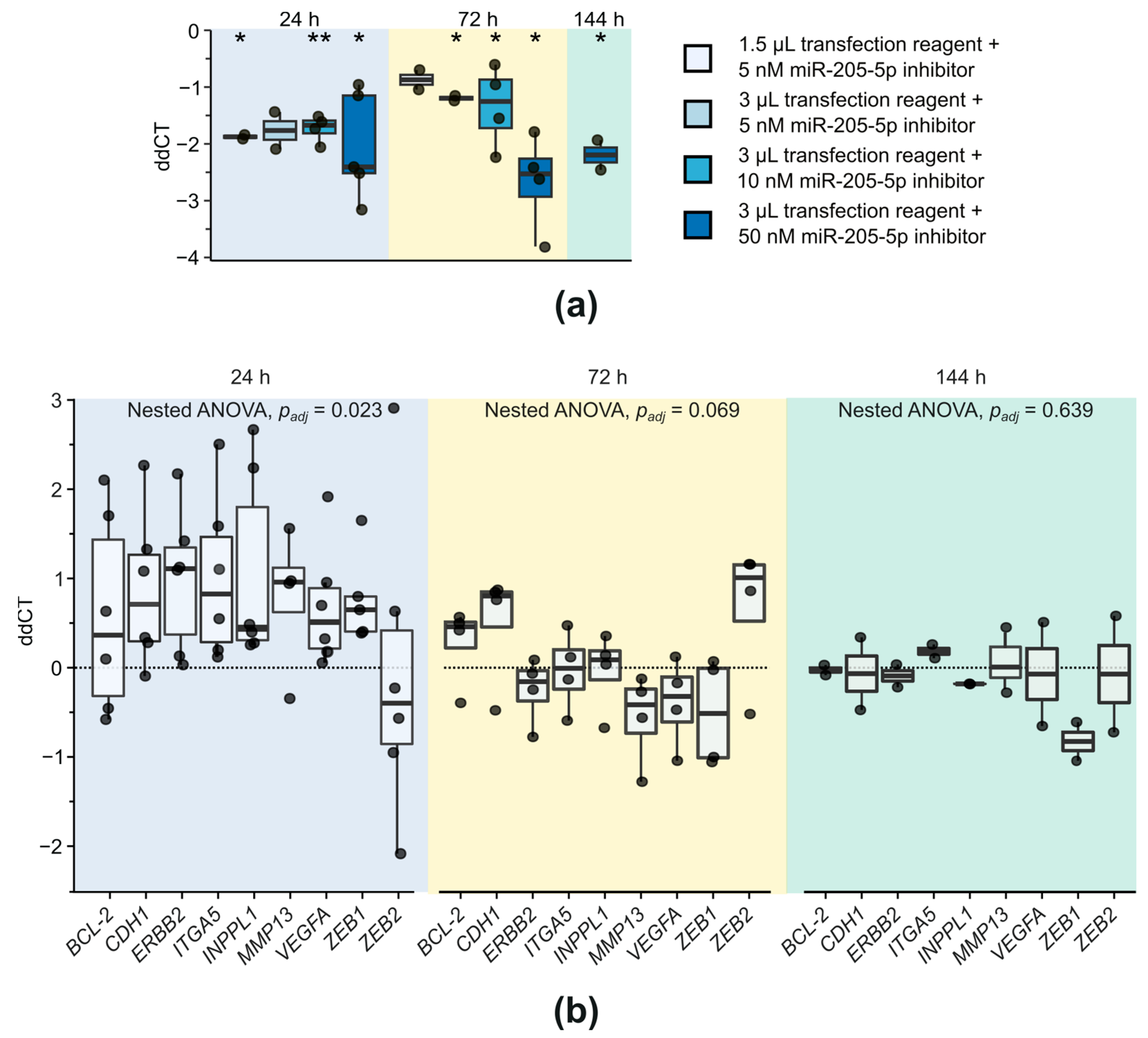
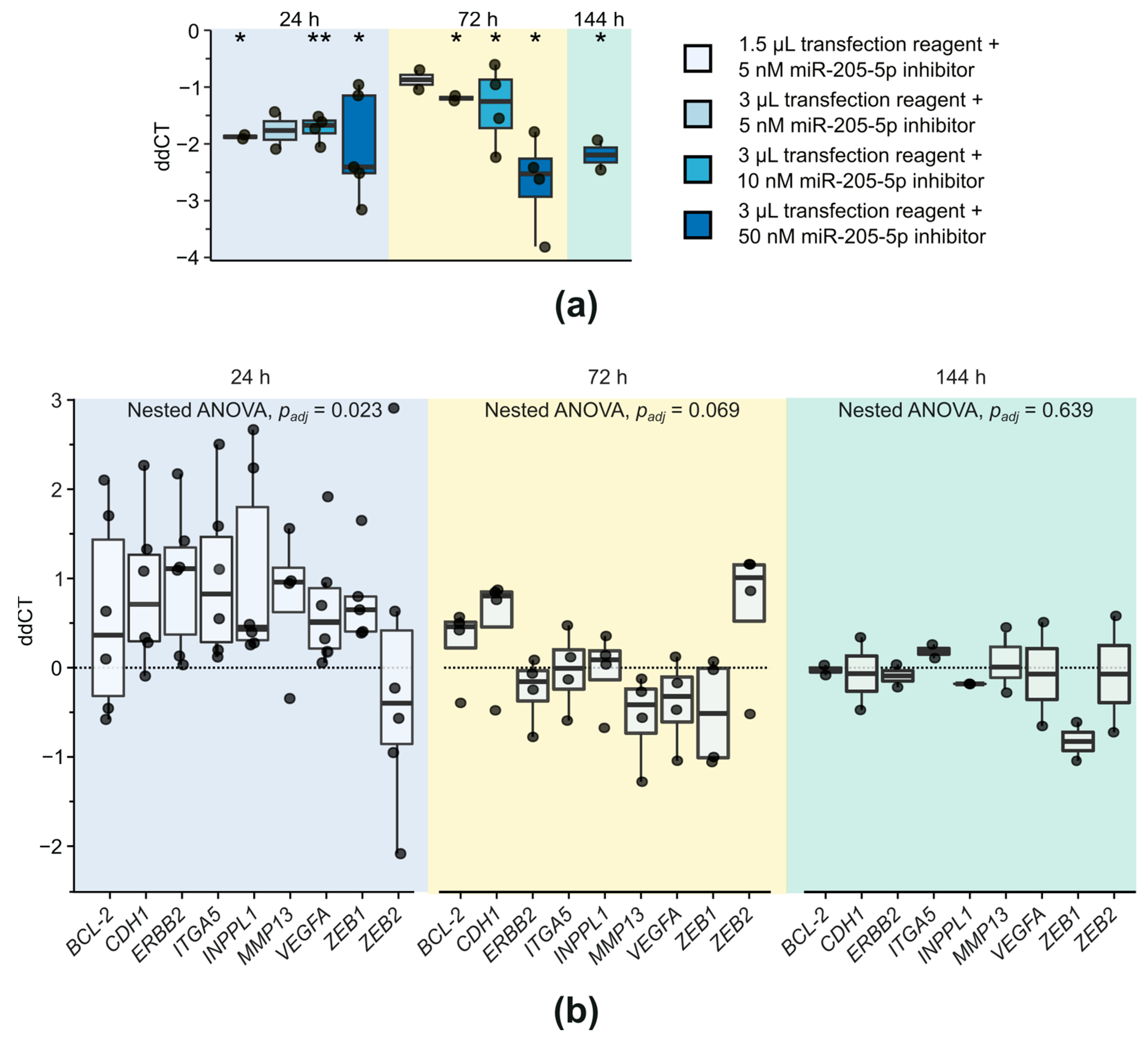
2.2. Investigation of Target Gene Expression after miR-205-5p Knockdown in SCC-12
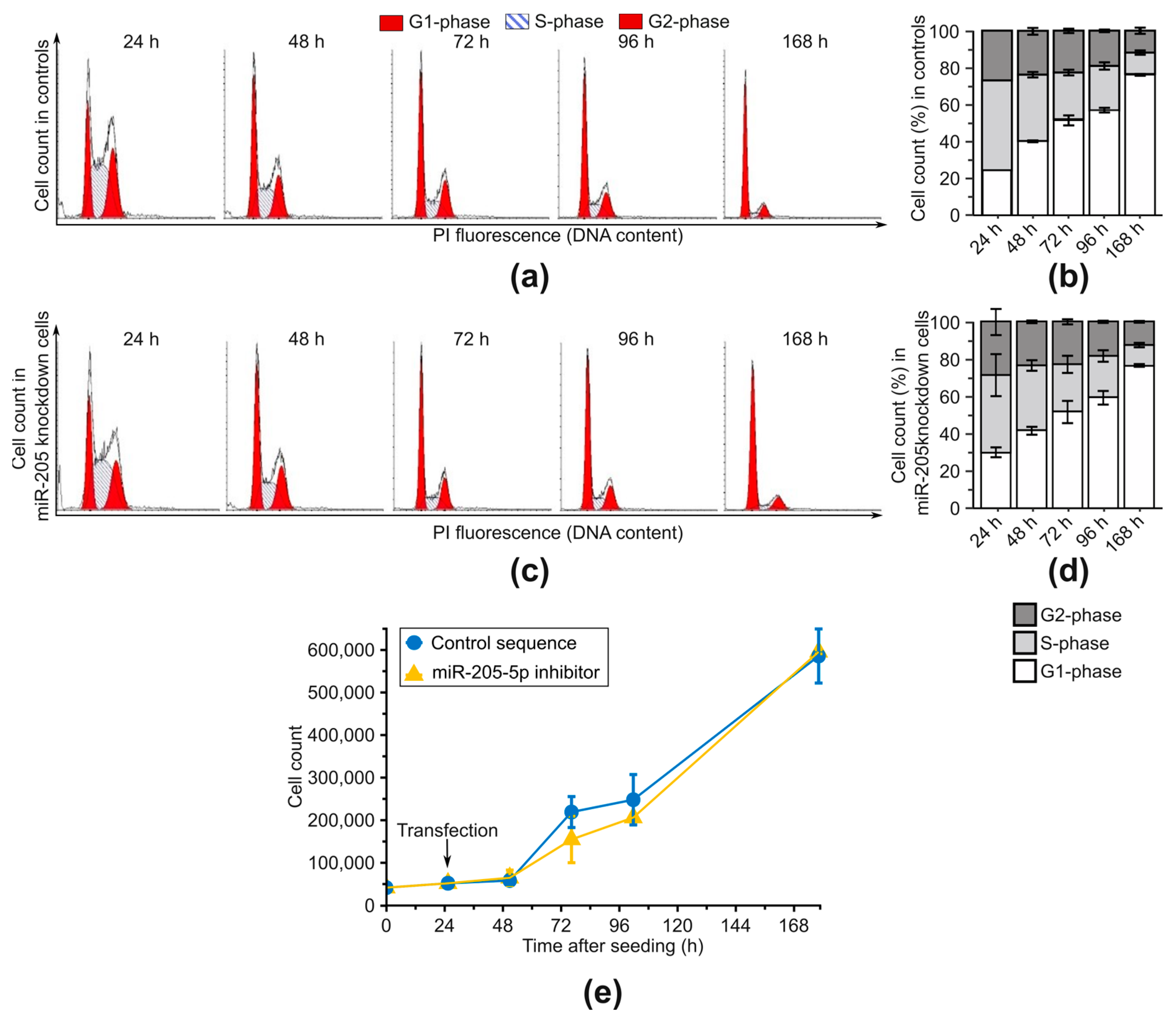
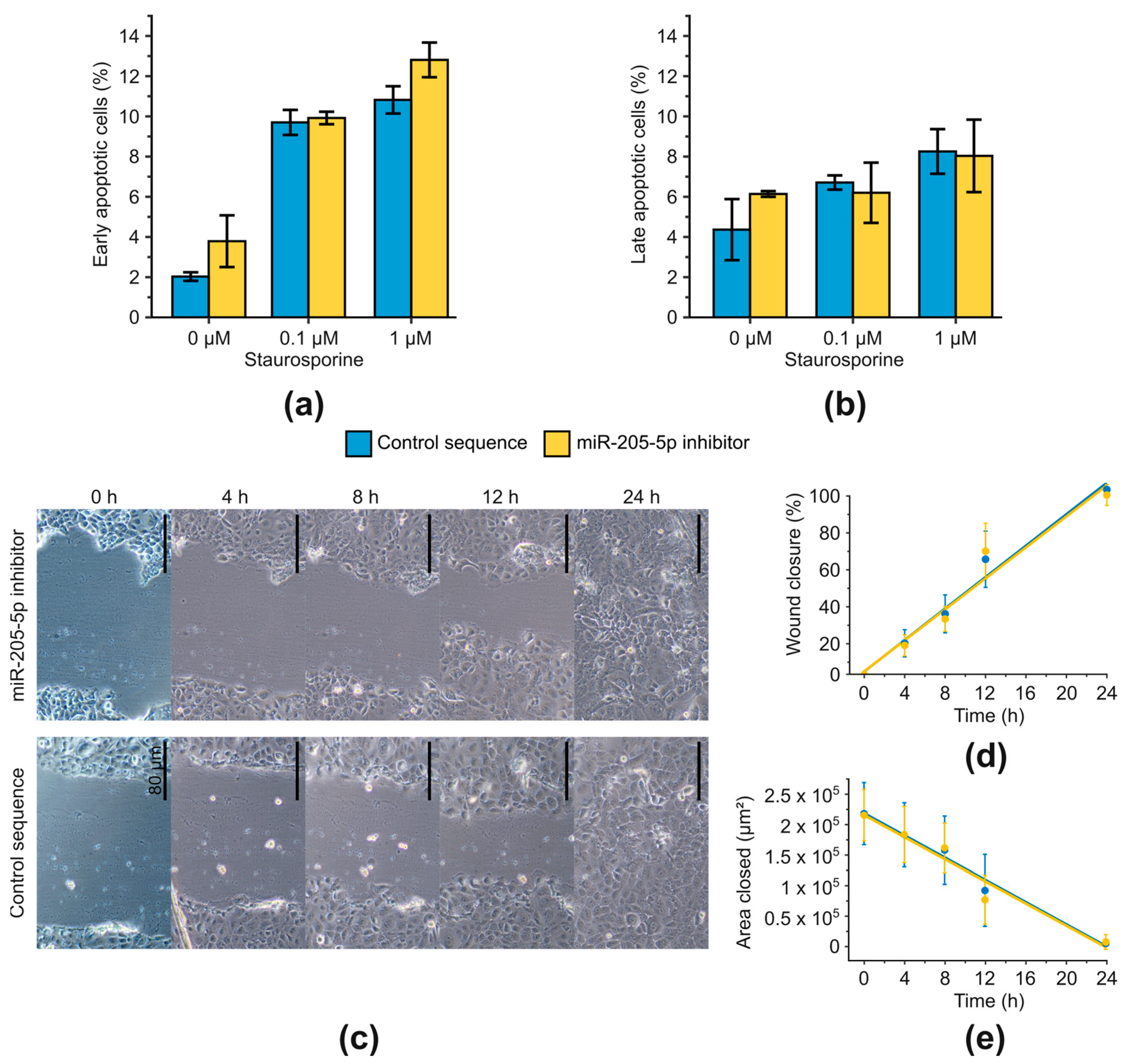
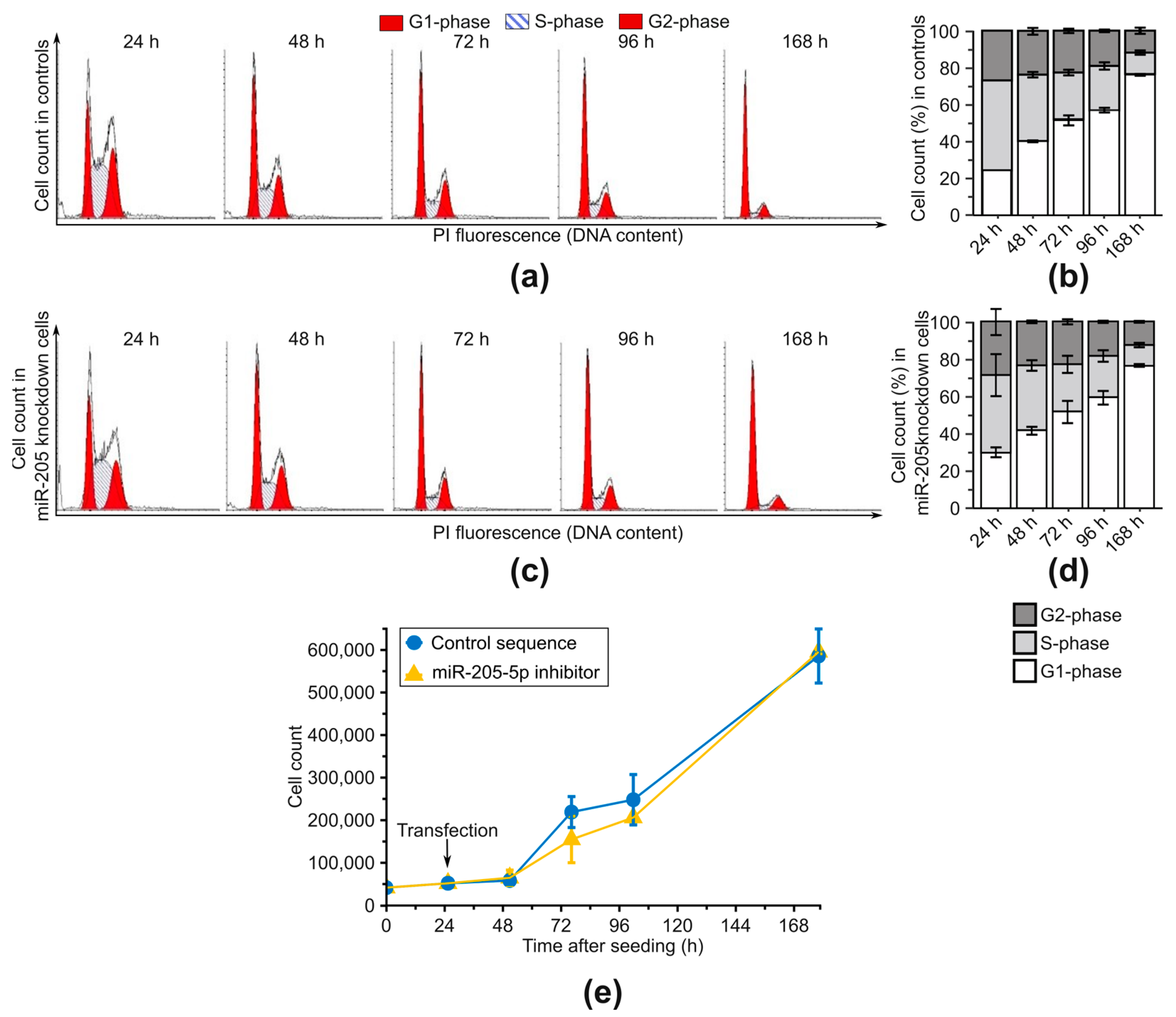
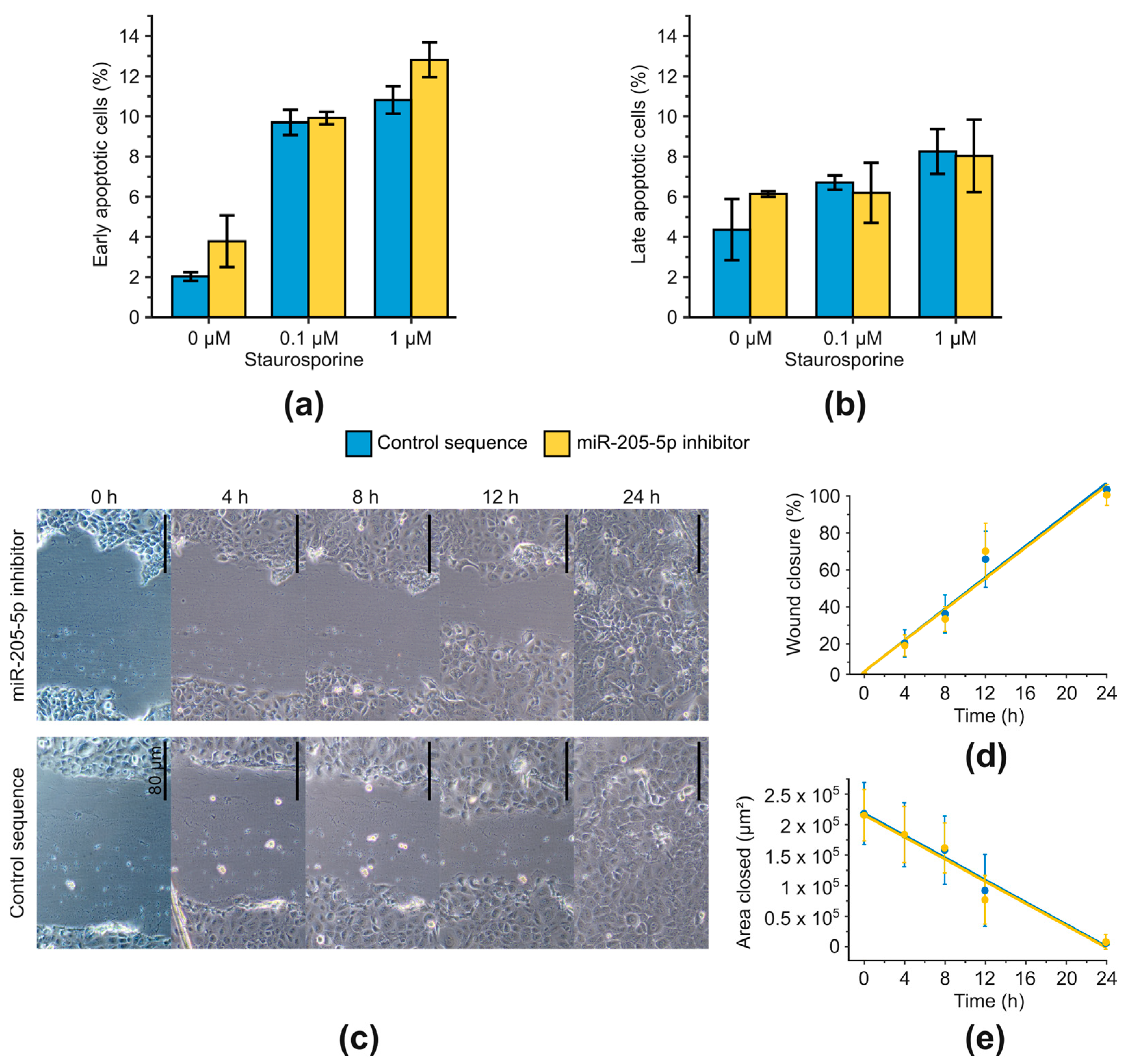
2.3. Examination of Phenotypic Changes after miR-205-5p Knockdown in SCC-12
3. Discussion
4. Materials and Methods
4.1. Cell Culture and SSR Irradiation
4.2. miRNA Profiling
4.3. Quantitative Real-Time Polymerase Chain Reaction (qPCR) for miRNA Detection
4.4. qPCR for Gene Expression Analysis
4.5. Knockdown of miR-205-5p
4.6. Cell Cycle Analysis
4.7. Cell Growth Assessment
4.8. Annexin V Assay
4.9. Scratch Assay
4.10. Statistical Analysis
4.11. Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karia, P.S.; Han, J.; Schmults, C.D. Cutaneous Squamous Cell Carcinoma: Estimated Incidence of Disease, Nodal Metastasis, and Deaths from Disease in the United States, 2012. J. Am. Acad. Dermatol. 2013, 68, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Mulvaney, P.M.; Schmults, C.D. Molecular Prediction of Metastasis in Cutaneous Squamous Cell Carcinoma. Curr. Opin. Oncol. 2020, 32, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Waldman, A.; Schmults, C. Cutaneous Squamous Cell Carcinoma. Hematol. Oncol. Clin. N. Am. 2019, 33, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Que, S.K.T.; Zwald, F.O.; Schmults, C.D. Cutaneous Squamous Cell Carcinoma. J. Am. Acad. Dermatol. 2018, 78, 237–247. [Google Scholar] [CrossRef]
- Ogata, D.; Tsuchida, T. Systemic Immunotherapy for Advanced Cutaneous Squamous Cell Carcinoma. Curr. Treat. Options Oncol. 2019, 20, 30. [Google Scholar] [CrossRef]
- Burton, K.A.; Ashack, K.A.; Khachemoune, A. Cutaneous Squamous Cell Carcinoma: A Review of High-Risk and Metastatic Disease. Am. J. Clin. Dermatol. 2016, 17, 491–508. [Google Scholar] [CrossRef]
- Migden, M.R.; Khushalani, N.I.; Chang, A.L.S.; Lewis, K.D.; Schmults, C.D.; Hernandez-Aya, L.; Meier, F.; Schadendorf, D.; Guminski, A.; Hauschild, A.; et al. Cemiplimab in Locally Advanced Cutaneous Squamous Cell Carcinoma: Results from an Open-Label, Phase 2, Single-Arm Trial. Lancet Oncol. 2020, 21, 294–305. [Google Scholar] [CrossRef]
- Fania, L.; Didona, D.; Di Pietro, F.R.; Verkhovskaia, S.; Morese, R.; Paolino, G.; Donati, M.; Ricci, F.; Coco, V.; Ricci, F.; et al. Cutaneous Squamous Cell Carcinoma: From Pathophysiology to Novel Therapeutic Approaches. Biomedicines 2021, 9, 171. [Google Scholar] [CrossRef]
- Syed, D.N.; Lall, R.K.; Mukhtar, H. MicroRNAs and Photocarcinogenesis. Photochem. Photobiol. 2015, 91, 173–187. [Google Scholar] [CrossRef]
- Kallini, J.R.; Hamed, N.; Khachemoune, A. Squamous Cell Carcinoma of the Skin: Epidemiology, Classification, Management, and Novel Trends. Int. J. Dermatol. 2015, 54, 130–140. [Google Scholar] [CrossRef]
- Mueller, S.A.; Gauthier, M.-E.A.; Ashford, B.; Gupta, R.; Gayevskiy, V.; Ch’ng, S.; Palme, C.E.; Shannon, K.; Clark, J.R.; Ranson, M.; et al. Mutational Patterns in Metastatic Cutaneous Squamous Cell Carcinoma. J. Investig. Dermatol. 2019, 139, 1449–1458.e1. [Google Scholar] [CrossRef] [PubMed]
- Rochette, P.J.; Lacoste, S.; Therrien, J.-P.; Bastien, N.; Brash, D.E.; Drouin, R. Influence of Cytosine Methylation on Ultraviolet-Induced Cyclobutane Pyrimidine Dimer Formation in Genomic DNA. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2009, 665, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-J.; Wu, Y.; Lv, Q.; Yang, X.-Y.; Jiang, M.-J.; Li, L.-M. Aberrant DNA Methylation in Cutaneous Squamous Cell Carcinoma. Int. J. Dermatol. Venereol. 2019, 2, 227–232. [Google Scholar] [CrossRef]
- Singh, A.; Willems, E.; Singh, A.; Ong, I.M.; Verma, A.K. Ultraviolet Radiation-Induced Differential MicroRNA Expression in the Skin of Hairless SKH1 Mice, a Widely Used Mouse Model for Dermatology Research. Oncotarget 2016, 7, 84924–84937. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- García-Sancha, N.; Corchado-Cobos, R.; Pérez-Losada, J.; Cañueto, J. MicroRNA Dysregulation in Cutaneous Squamous Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 2181. [Google Scholar] [CrossRef]
- Qiu, C.; Chen, G.; Cui, Q. Towards the Understanding of MicroRNA and Environmental Factor Interactions and Their Relationships to Human Diseases. Sci. Rep. 2012, 2, 318. [Google Scholar] [CrossRef]
- Kraemer, A.; Chen, I.-P.; Henning, S.; Faust, A.; Volkmer, B.; Atkinson, M.J.; Moertl, S.; Greinert, R. UVA and UVB Irradiation Differentially Regulate MicroRNA Expression in Human Primary Keratinocytes. PLoS ONE 2013, 8, e83392. [Google Scholar] [CrossRef]
- Tonolli, P.N.; Chiarelli-Neto, O.; Santacruz-Perez, C.; Junqueira, H.C.; Watanabe, I.-S.; Ravagnani, F.G.; Martins, W.K.; Baptista, M.S. Lipofuscin Generated by UVA Turns Keratinocytes Photosensitive to Visible Light. J. Investig. Dermatol. 2017, 137, 2447–2450. [Google Scholar] [CrossRef]
- Schroeder, P.; Calles, C.; Benesova, T.; Macaluso, F.; Krutmann, J. Photoprotection beyond Ultraviolet Radiation—Effective Sun Protection Has to Include Protection against Infrared A Radiation-Induced Skin Damage. Ski. Pharmacol. Physiol. 2010, 23, 15–17. [Google Scholar] [CrossRef]
- Bender, M.; Chen, I.-P.; Degenhardt, S.; Henning, S.; Volkmer, B.; Greinert, R. MicroRNA Dysregulation in Cutaneous Squamous Cell Carcinoma and Implication of UV Impact. 2023; Manuscript in preparation. [Google Scholar]
- Ferrari, E.; Gandellini, P. Unveiling the Ups and Downs of MiR-205 in Physiology and Cancer: Transcriptional and Post-Transcriptional Mechanisms. Cell Death Dis. 2020, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sendra, B.; Serna, E.; Navarro, L.; González-Muñoz, J.F.; Portero, J.; Ramos, A.; Murgui, A.; Monteagudo, C. Transcriptomic Identification of MiR-205 Target Genes Potentially Involved in Metastasis and Survival of Cutaneous Malignant Melanoma. Sci. Rep. 2020, 10, 4771. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Arnedo, A.; Torres Figueroa, F.; Clavijo, C.; Arbeláez, P.; Cruz, J.C.; Muñoz-Camargo, C. An Image J Plugin for the High Throughput Image Analysis of in Vitro Scratch Wound Healing Assays. PLoS ONE 2020, 15, e0232565. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Gazel, A.; Pintucci, G.; Shuck, A.; Shifteh, S.; Ginsburg, D.; Rao, L.S.; Kaneko, T.; Freedberg, I.M.; Tamaki, K.; et al. Specificity in Stress Response: Epidermal Keratinocytes Exhibit Specialized UV-Responsive Signal Transduction Pathways. DNA Cell Biol. 2003, 22, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, S.; Bil, P.; Gajda, K.; Poterala-Hejmo, A.; Hudy, D.; Rzeszowska-Wolny, J. Cell Type-Specific Differences in Redox Regulation and Proliferation after Low UVA Doses. PLoS ONE 2019, 14, e0205215. [Google Scholar] [CrossRef]
- Chen, I.-P.; Bender, M.; Spassova, I.; Henning, S.; Kubat, L.; Fan, K.; Degenhardt, S.; Mhamdi-Ghodbani, M.; Sriram, A.; Volkmer, B.; et al. UV-Type Specific Alteration of MiRNA Expression and Its Association with Tumor Progression and Metastasis in SCC Cell Lines. J. Cancer Res. Clin. Oncol. 2020, 146, 3215–3231. [Google Scholar] [CrossRef]
- McAbee, J.H.; Degorre-Kerbaul, C.; Valdez, K.; Wendler, A.; Shankavaram, U.T.; Watts, C.; Camphausen, K.; Tofilon, P.J. Detection of Glioblastoma Intratumor Heterogeneity in Radiosensitivity Using Patient-Derived Neurosphere Cultures. J. Neurooncol. 2020, 149, 383–390. [Google Scholar] [CrossRef]
- Alfonso, J.C.L.; Berk, L. Modeling the Effect of Intratumoral Heterogeneity of Radiosensitivity on Tumor Response over the Course of Fractionated Radiation Therapy. Radiat. Oncol. 2019, 14, 88. [Google Scholar] [CrossRef]
- Todorovic, V.; Prevc, A.; Zakelj, M.N.; Savarin, M.; Brozic, A.; Groselj, B.; Strojan, P.; Cemazar, M.; Sersa, G. Mechanisms of Different Response to Ionizing Irradiation in Isogenic Head and Neck Cancer Cell Lines. Radiat. Oncol. 2019, 14, 214. [Google Scholar] [CrossRef]
- Pickering, C.R.; Zhou, J.H.; Lee, J.J.; Drummond, J.A.; Peng, S.A.; Saade, R.E.; Tsai, K.Y.; Curry, J.L.; Tetzlaff, M.T.; Lai, S.Y.; et al. Mutational Landscape of Aggressive Cutaneous Squamous Cell Carcinoma. Clin. Cancer Res. 2014, 20, 6582–6592. [Google Scholar] [CrossRef]
- Hernandez-Onate, M.A.; Herrera-Estrella, A. Damage Response Involves Mechanisms Conserved across Plants, Animals and Fungi. Curr. Genet. 2015, 61, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Engelberg, D. The UV Response Involving the Ras Signaling Pathway and AP-1 Transcription Factors Is Conserved between Yeast and Mammals. Cell 1994, 77, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.I.L.; Roca, C.P.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Identifying Conserved UV Exposure Genes and Mechanisms. Sci. Rep. 2018, 8, 8605. [Google Scholar] [CrossRef] [PubMed]
- McFadden, D.G.; Politi, K.; Bhutkar, A.; Chen, F.K.; Song, X.; Pirun, M.; Santiago, P.M.; Kim-Kiselak, C.; Platt, J.T.; Lee, E.; et al. Mutational Landscape of EGFR-, MYC-, and Kras-Driven Genetically Engineered Mouse Models of Lung Adenocarcinoma. Proc. Natl. Acad. Sci. USA 2016, 113, E6409–E6417. [Google Scholar] [CrossRef] [PubMed]
- Nowell, C.; Radtke, F. Cutaneous Notch Signaling in Health and Disease. Cold Spring Harb. Perspect. Med. 2013, 3, a017772. [Google Scholar] [CrossRef]
- Mullany, L.K.; White, P.; Hanse, E.A.; Nelsen, C.J.; Goggin, M.M.; Mullany, J.E.; Anttila, C.K.; Greenbaum, L.E.; Kaestner, K.H.; Albrecht, J.H. Distinct Proliferative and Transcriptional Effects of the D-Type Cyclins in Vivo. Cell Cycle 2008, 7, 2215–2224. [Google Scholar] [CrossRef]
- Gillespie, J.; Skeeles, L.E.; Allain, D.C.; Kent, M.N.; Peters, S.B.; Nagarajan, P.; Yu, L.; Teknos, T.N.; Olencki, T.; Toland, A.E. MicroRNA Expression Profiling in Metastatic Cutaneous Squamous Cell Carcinoma. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1043–1045. [Google Scholar] [CrossRef]
- Mizrahi, A.; Barzilai, A.; Gur-Wahnon, D.; Ben-Dov, I.Z.; Glassberg, S.; Meningher, T.; Elharar, E.; Masalha, M.; Jacob-Hirsch, J.; Tabibian-Keissar, H.; et al. Alterations of MicroRNAs throughout the Malignant Evolution of Cutaneous Squamous Cell Carcinoma: The Role of MiR-497 in Epithelial to Mesenchymal Transition of Keratinocytes. Oncogene 2018, 37, 218–230. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczynski, A.; Romanowicz, H.; Szyllo, K.; Hogendorf, P. MiRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The MiR-200 Family and MiR-205 Regulate Epithelial to Mesenchymal Transition by Targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef]
- Yu, J.; Peng, H.; Ruan, Q.; Fatima, A.; Getsios, S.; Lavker, R.M. MicroRNA-205 Promotes Keratinocyte Migration via the Lipid Phosphatase SHIP2. FASEB J. 2010, 24, 3950–3959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, J.; Pang, X.; Chen, Z.; Zhang, Z.; Lei, L.; Xu, H.; Wen, L.; Zhu, J.; Jiang, Y.; et al. MiR-205–5p Suppresses Angiogenesis in Gastric Cancer by Downregulating the Expression of VEGFA and FGF1. Exp. Cell Res. 2021, 404, 112579. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Tan, S.; Yang, L.; Chen, X.; Yang, R.; Oyang, L.; Lin, J.; Xia, L.; Wu, N.; Han, Y.; et al. Exosomal MiR-205-5p Enhances Angiogenesis and Nasopharyngeal Carcinoma Metastasis by Targeting Desmocollin-2. Mol. Ther. Oncolytics 2022, 24, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Canueto, J.; Cardenoso-Alvarez, E.; Garcia-Hernandez, J.L.; Galindo-Villardón, P.; Vicente-Galindo, P.; Vicente-Villardon, J.L.; Alonso-Lopez, D.; De Las Rivas, J.; Valero, J.; Moyano-Sanz, E.; et al. Micro RNA (MiR)-203 and MiR-205 Expression Patterns Identify Subgroups of Prognosis in Cutaneous Squamous Cell Carcinoma. Br. J. Dermatol. 2017, 177, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Bruegger, C.; Kempf, W.; Spoerri, I.; Arnold, A.W.; Itin, P.H.; Burger, B. MicroRNA Expression Differs in Cutaneous Squamous Cell Carcinomas and Healthy Skin of Immunocompetent Individuals. Exp. Dermatol. 2013, 22, 426–428. [Google Scholar] [CrossRef] [PubMed]
- Skourti, E.; Logotheti, S.; Kontos, C.K.; Pavlopoulou, A.; Dimoragka, P.T.; Trougakos, I.P.; Gorgoulis, V.; Scorilas, A.; Michalopoulos, I.; Zoumpourlis, V. Progression of Mouse Skin Carcinogenesis Is Associated with the Orchestrated Deregulation of Mir-200 Family Members, Mir-205 and Their Common Targets. Mol. Carcinog. 2016, 55, 1229–1242. [Google Scholar] [CrossRef]
- Dziunycz, P.; Iotzova-Weiss, G.; Eloranta, J.J.; Läuchli, S.; Hafner, J.; French, L.E.; Hofbauer, G.F.L. Squamous Cell Carcinoma of the Skin Shows a Distinct MicroRNA Profile Modulated by UV Radiation. J. Investig. Dermatol. 2010, 130, 2686–2689. [Google Scholar] [CrossRef]
- Krutzfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of MicroRNAs in Vivo with ‘Antagomirs’ . Nature 2005, 438, 685–689. [Google Scholar] [CrossRef]
- De Cola, A.; Lamolinara, A.; Lanuti, P.; Rossi, C.; Iezzi, M.; Marchisio, M.; Todaro, M.; De Laurenzi, V. MiR-205-5p Inhibition by Locked Nucleic Acids Impairs Metastatic Potential of Breast Cancer Cells. Cell Death Dis. 2018, 9, 821. [Google Scholar] [CrossRef]
- Yang, F.; Ning, Z.; Ma, L.; Liu, W.; Shao, C.; Shu, Y.; Shen, H. Exosomal MiRNAs and MiRNA Dysregulation in Cancer-Associated Fibroblasts. Mol. Cancer 2017, 16, 148. [Google Scholar] [CrossRef]
- Tan, S.; Xia, L.; Yi, P.; Han, Y.; Tang, L.; Pan, Q.; Tian, Y.; Rao, S.; Oyang, L.; Liang, J.; et al. Exosomal MiRNAs in Tumor Microenvironment. J. Exp. Clin. Cancer Res. 2020, 39, 67. [Google Scholar] [CrossRef] [PubMed]
- Durendez-Saez, E.; Torres-Martinez, S.; Calabuig-Farinas, S.; Meri-Abad, M.; Ferrero-Gimeno, M.; Camps, C. Exosomal MicroRNAs in Non-Small Cell Lung Cancer. Transl. Cancer Res. 2021, 10, 3128–3139. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, F.; Xiao, Z.; Yao, L. Exosomal MicroRNA-205 Is Involved in Proliferation, Migration, Invasion, and Apoptosis of Ovarian Cancer Cells via Regulating VEGFA. Cancer Cell Int. 2019, 19, 281. [Google Scholar] [CrossRef] [PubMed]
- Popp, S.; Waltering, S.; Boukamp, P.; Holtgreve-Grez, H.; Jauch, A.; Proby, C.; Leigh, I.M. Genetic Characterization of a Human Skin Carcinoma Progression Model: From Primary Tumor to Metastasis. J. Investig. Dermatol. 2000, 115, 1095–1103. [Google Scholar] [CrossRef]
- Popp, S.; Waltering, S.; Herbst, C.; Moll, I.; Boukamp, P. UV-B-Type Mutations and Chromosomal Imbalances Indicate Common Pathways for the Development of Merkel and Skin Squamous Cell Carcinomas. Int. J. Cancer 2002, 99, 352–360. [Google Scholar] [CrossRef]
- Masters, J.R.W.; Palsson, B. Human Cell Culture; Springer: Dordrecht, The Netherlands, 1999; Volume 1, ISBN 978-0-7923-5143-6. [Google Scholar]
- Tackett, M.R.; Diwan, I. Using FirePlex Particle Technology for Multiplex MicroRNA Profiling without RNA Purification. Methods Mol. Biol. 2017, 1654, 209–219. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-DeltadeltaCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Beal, J. Biochemical Complexity Drives Log-normal Variation in Genetic Expression. Eng. Biol. 2017, 1, 55–60. [Google Scholar] [CrossRef]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.-P.; Meese, E.; et al. MiRTargetLink 2.0-Interactive MiRNA Target Gene and Target Pathway Networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Storey, K.B. RBiomirGS: An All-in-One MiRNA Gene Set Analysis Solution Featuring Target MRNA Mapping and Expression Profile Integration. PeerJ 2018, 6, e4262. [Google Scholar] [CrossRef]
- Ma, X.; Wu, D.; Zhang, X.; Shao, X.; Hu, G. MicroRNA-214 Prevents Traits of Cutaneous Squamous Cell Carcinoma via VEGFA and Bcl-2. Technol. Cancer Res. Treat. 2020, 19, 153303382098009. [Google Scholar] [CrossRef] [PubMed]
- Dahlhoff, M.; Muzumdar, S.; Schäfer, M.; Schneider, M.R. ERBB2 Is Essential for the Growth of Chemically Induced Skin Tumors in Mice. J. Investig. Dermatol. 2017, 137, 921–930. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, C.H.; Bae, J.T.; Nam, K.T.; Moon, D.B.; Hwang, O.K.; Choi, J.S.; Kim, T.H.; Jun, H.O.; Jung, Y.S.; et al. Inhibition of Skin Carcinogenesis by Suppression of NF-KappaB Dependent ITGAV and TIMP-1 Expression in IL-32gamma Overexpressed Condition. J. Exp. Clin. Cancer Res. 2018, 37, 293. [Google Scholar] [CrossRef]
- Xu, N.; Zhang, L.; Meisgen, F.; Harada, M.; Heilborn, J.; Homey, B.; Grandér, D.; Ståhle, M.; Sonkoly, E.; Pivarcsi, A. MicroRNA-125b Down-Regulates Matrix Metallopeptidase 13 and Inhibits Cutaneous Squamous Cell Carcinoma Cell Proliferation, Migration, and Invasion. J. Biol. Chem. 2012, 287, 29899–29908. [Google Scholar] [CrossRef]
- Genenger, B.; Perry, J.R.; Ashford, B.; Ranson, M. A TEMTing Target? Clinical and Experimental Evidence for Epithelial-Mesenchymal Transition in the Progression of Cutaneous Squamous Cell Carcinoma (a Scoping Systematic Review). Discov. Oncol. 2022, 13, 42. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bender, M.; Chen, I.-P.; Henning, S.; Degenhardt, S.; Mhamdi-Ghodbani, M.; Starzonek, C.; Volkmer, B.; Greinert, R. Knockdown of Simulated-Solar-Radiation-Sensitive miR-205-5p Does Not Induce Progression of Cutaneous Squamous Cell Carcinoma In Vitro. Int. J. Mol. Sci. 2023, 24, 16428. https://doi.org/10.3390/ijms242216428
Bender M, Chen I-P, Henning S, Degenhardt S, Mhamdi-Ghodbani M, Starzonek C, Volkmer B, Greinert R. Knockdown of Simulated-Solar-Radiation-Sensitive miR-205-5p Does Not Induce Progression of Cutaneous Squamous Cell Carcinoma In Vitro. International Journal of Molecular Sciences. 2023; 24(22):16428. https://doi.org/10.3390/ijms242216428
Chicago/Turabian StyleBender, Marc, I-Peng Chen, Stefan Henning, Sarah Degenhardt, Mouna Mhamdi-Ghodbani, Christin Starzonek, Beate Volkmer, and Rüdiger Greinert. 2023. "Knockdown of Simulated-Solar-Radiation-Sensitive miR-205-5p Does Not Induce Progression of Cutaneous Squamous Cell Carcinoma In Vitro" International Journal of Molecular Sciences 24, no. 22: 16428. https://doi.org/10.3390/ijms242216428
APA StyleBender, M., Chen, I.-P., Henning, S., Degenhardt, S., Mhamdi-Ghodbani, M., Starzonek, C., Volkmer, B., & Greinert, R. (2023). Knockdown of Simulated-Solar-Radiation-Sensitive miR-205-5p Does Not Induce Progression of Cutaneous Squamous Cell Carcinoma In Vitro. International Journal of Molecular Sciences, 24(22), 16428. https://doi.org/10.3390/ijms242216428