
Genome-Wide Differential Transcription of Long Noncoding RNAs in Psoriatic Skin


Abstract
:1. Introduction
2. Results
2.1. GSE54456 Analysis
2.2. GSE121212 Analysis
2.3. General Analysis
3. Discussion
4. Materials and Methods
4.1. Data Collection
4.2. Computational Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raharja, A.; Mahil, S.K.; Barker, J.N. Psoriasis: A brief overview. Clin. Med. 2021, 21, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Mehrmal, S.; Uppal, P.; Nedley, N.; Giesey, R.L.; Delost, G.R. The global, regional, and national burden of psoriasis in 195 countries and territories, 1990 to 2017: A systematic analysis from the Global Burden of Disease Study 2017. J. Am. Acad. Dermatol. 2021, 84, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Rendon, A.; Schakel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef] [PubMed]
- Evans, C. Managed care aspects of psoriasis and psoriatic arthritis. Am. J. Manag. Care 2016, 22 (Suppl. S8), s238–s243. [Google Scholar]
- Adesanya, E.I.; Matthewman, J.; Schonmann, Y.; Hayes, J.F.; Henderson, A.; Mathur, R.; Mulick, A.R.; Smith, C.H.; Langan, S.M.; Mansfield, K.E. Factors associated with depression, anxiety and severe mental illness among adults with atopic eczema or psoriasis: A systematic review and meta-analysis. Br. J. Dermatol. 2023, 188, 460–470. [Google Scholar] [CrossRef]
- Koks, S.; Keermann, M.; Reimann, E.; Prans, E.; Abram, K.; Silm, H.; Koks, G.; Kingo, K. Psoriasis-Specific RNA Isoforms Identified by RNA-Seq Analysis of 173,446 Transcripts. Front. Med. 2016, 3, 46. [Google Scholar] [CrossRef]
- Vandenbroeck, K. (Ed.) Cytokine Gene Polymorphisms in Multifactorial Conditions, 1st ed.; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Dand, N.; Stuart, P.E.; Bowes, J.; Ellinghaus, D.; Nititham, J.; Saklatvala, J.R.; Teder-Laving, M.; Thomas, L.F.; Traks, T.; Uebe, S.; et al. GWAS meta-analysis of psoriasis identifies new susceptibility alleles impacting disease mechanisms and therapeutic targets. medRxiv 2023. [Google Scholar] [CrossRef]
- Moltrasio, C.; Romagnuolo, M.; Marzano, A.V. Epigenetic Mechanisms of Epidermal Differentiation. Int. J. Mol. Sci. 2022, 23, 4874. [Google Scholar] [CrossRef]
- Liu, J.; Chang, H.W.; Huang, Z.M.; Nakamura, M.; Sekhon, S.; Ahn, R.; Munoz-Sandoval, P.; Bhattarai, S.; Beck, K.M.; Sanchez, I.M.; et al. Single-cell RNA sequencing of psoriatic skin identifies pathogenic Tc17 cell subsets and reveals distinctions between CD8+ T cells in autoimmunity and cancer. J. Allergy Clin. Immunol. 2021, 147, 2370–2380. [Google Scholar] [CrossRef]
- Krishnan, V.S.; Koks, S. Transcriptional Basis of Psoriasis from Large Scale Gene Expression Studies: The Importance of Moving towards a Precision Medicine Approach. Int. J. Mol. Sci. 2022, 23, 6130. [Google Scholar] [CrossRef]
- Rioux, G.; Ridha, Z.; Simard, M.; Turgeon, F.; Guerin, S.L.; Pouliot, R. Transcriptome Profiling Analyses in Psoriasis: A Dynamic Contribution of Keratinocytes to the Pathogenesis. Genes 2020, 11, 1155. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef]
- Clark, M.B.; Mattick, J.S. Long noncoding RNAs in cell biology. Semin. Cell Dev. Biol. 2011, 22, 366–376. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef]
- Chen, X.; Guo, G.; Lu, Y.; Wang, S.; Zhang, Y.; Huang, Q. Mechanisms and functions of long non-coding RNAs in glioma (Review). Oncol. Rep. 2021, 45, 9. [Google Scholar] [CrossRef]
- Watanabe, Y.; Yamamoto, M.S. pombe mei2+ encodes an RNA-binding protein essential for premeiotic DNA synthesis and meiosis I, which cooperates with a novel RNA species meiRNA. Cell 1994, 78, 487–498. [Google Scholar] [CrossRef]
- Campalans, A.; Kondorosi, A.; Crespi, M. Enod40, a short open reading frame-containing mRNA, induces cytoplasmic localization of a nuclear RNA binding protein in Medicago truncatula. Plant Cell 2004, 16, 1047–1059. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Nadhan, R.; Isidoro, C.; Song, Y.S.; Dhanasekaran, D.N. Signaling by LncRNAs: Structure, Cellular Homeostasis, and Disease Pathology. Cells 2022, 11, 2517. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, M.; Kitagawa, K.; Kotake, Y.; Niida, H.; Ohhata, T. Cell cycle regulation by long non-coding RNAs. Cell. Mol. Life Sci. 2013, 70, 4785–4794. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Aghabalazade, A.; Shoorei, H.; Majidpoor, J.; Taheri, M.; Mokhtari, M. The Impact of lncRNAs and miRNAs on Apoptosis in Lung Cancer. Front. Oncol. 2021, 11, 714795. [Google Scholar] [CrossRef]
- DiStefano, J.K. The Emerging Role of Long Noncoding RNAs in Human Disease. Methods Mol. Biol. 2018, 1706, 91–110. [Google Scholar]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging role of non-coding RNA in health and disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef]
- Yan, J.; Song, J.; Qiao, M.; Zhao, X.; Li, R.; Jiao, J.; Sun, Q. Long noncoding RNA expression profile and functional analysis in psoriasis. Mol. Med. Rep. 2019, 19, 3421–3430. [Google Scholar] [CrossRef]
- Song, J.K.; Yin, S.Y.; Li, W.; Li, X.D.; Luo, Y.; Luo, Y.; Xing, M.; Li, B.; Kuai, L. An update on the role of long non-coding RNAs in psoriasis. Chin. Med. J. 2020, 134, 379–389. [Google Scholar] [CrossRef]
- Shefler, A.; Patrick, M.T.; Wasikowski, R.; Chen, J.; Sarkar, M.K.; Gudjonsson, J.E.; Tsoi, L.C. Skin-Expressing lncRNAs in Inflammatory Responses. Front. Genet. 2022, 13, 835740. [Google Scholar] [CrossRef]
- Lin, J.; Li, X.; Zhang, F.; Zhu, L.; Chen, Y. Transcriptome wide analysis of long non-coding RNA-associated ceRNA regulatory circuits in psoriasis. J. Cell. Mol. Med. 2021, 25, 6925–6935. [Google Scholar] [CrossRef] [PubMed]
- Dand, N.; Mahil, S.K.; Capon, F.; Smith, C.H.; Simpson, M.A.; Barker, J.N. Psoriasis and Genetics. Acta Derm. Venereol. 2020, 100, adv00030. [Google Scholar] [CrossRef]
- Ogawa, K.; Okada, Y. The current landscape of psoriasis genetics in 2020. J. Dermatol. Sci. 2020, 99, 2–8. [Google Scholar] [CrossRef]
- Qiao, M.; Li, R.; Zhao, X.; Yan, J.; Sun, Q. Up-regulated lncRNA-MSX2P1 promotes the growth of IL-22-stimulated keratinocytes by inhibiting miR-6731-5p and activating S100A7. Exp. Cell Res. 2018, 363, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Ahn, R.; Lai, K.; Mullins, E.; Debbaneh, M.; Dimon, M.; Arron, S.; Liao, W. Landscape of long non-coding RNAs in psoriatic and healthy skin. J. Investig. Dermatol. 2016, 136, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Wu, T.; Yu, L.; Zhou, N.; Zhang, Z.; Pu, Y.; Wu, J.; Shu, H. Exploration of Biomarkers of Psoriasis through Combined Multiomics Analysis. Mediat. Inflamm. 2022, 2022, 7731082. [Google Scholar] [CrossRef]
- Hangauer, M.J.; Vaughn, I.W.; McManus, M.T. Pervasive transcription of the human genome produces thousands of previously unidentified long intergenic noncoding RNAs. PLoS Genet. 2013, 9, e1003569. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H.; Taylor, M.; Cook, C.; Martinez-Berdeja, A.; North, J.P.; Harirchian, P.; Hailer, A.A.; Zhao, Z.; Ghadially, R.; et al. Classification of human chronic inflammatory skin disease based on single-cell immune profiling. Sci. Immunol. 2022, 7, eabl9165. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, Z.; Chen, F. Association of Key Genes and Pathways with Atopic Dermatitis by Bioinformatics Analysis. Med. Sci. Monit. 2019, 25, 4353–4361. [Google Scholar] [CrossRef]
- Frazier, W.; Bhardwaj, N. Atopic Dermatitis: Diagnosis and Treatment. Am. Fam. Physician 2020, 101, 590–598. [Google Scholar]
- Liu, W.; Tan, S.; Bai, X.; Ma, S.; Chen, X. Long non-coding RNA LINC01215 promotes epithelial-mesenchymal transition and lymph node metastasis in epithelial ovarian cancer through RUNX3 promoter methylation. Transl. Oncol. 2021, 14, 101135. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhang, F.; Zhang, Z.; Zhong, Y.; Chen, Y. lncRNA LINC01206 regulates the cell cycles in psoriasis. J. Diagn. Ther. Derm.Venereol. 2021, 28, 343–347. [Google Scholar]
- Yang, Z.; Chen, Z.; Wang, C.; Huang, P.; Luo, M.; Zhou, R. STAT3/SH3PXD2A-AS1/miR-125b/STAT3 positive feedback loop affects psoriasis pathogenesis via regulating human keratinocyte proliferation. Cytokine 2021, 144, 155535. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Tsoi, L.C.; Swindell, W.R.; Gudjonsson, J.E.; Tejasvi, T.; Johnston, A.; Ding, J.; Stuart, P.E.; Xing, X.; Kochkodan, J.J.; et al. Transcriptome analysis of psoriasis in a large case-control sample: RNA-seq provides insights into disease mechanisms. J. Investig. Dermatol. 2014, 134, 1828–1838. [Google Scholar] [CrossRef]
- Song, J.; Kim, D.; Lee, S.; Jung, J.; Joo, J.W.J.; Jang, W. Integrative transcriptome-wide analysis of atopic dermatitis for drug repositioning. Commun. Biol. 2022, 5, 615. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Smedley, D.; Haider, S.; Ballester, B.; Holland, R.; London, D.; Thorisson, G.; Kasprzyk, A. BioMart–biological queries made easy. BMC Genom. 2009, 10, 22. [Google Scholar] [CrossRef]
- Severin, J.; Lizio, M.; Harshbarger, J.; Kawaji, H.; Daub, C.O.; Hayashizaki, Y.; Consortium, F.; Bertin, N.; Forrest, A.R. Interactive visualization and analysis of large-scale sequencing datasets using ZENBU. Nat. Biotechnol. 2014, 32, 217–219. [Google Scholar] [CrossRef]
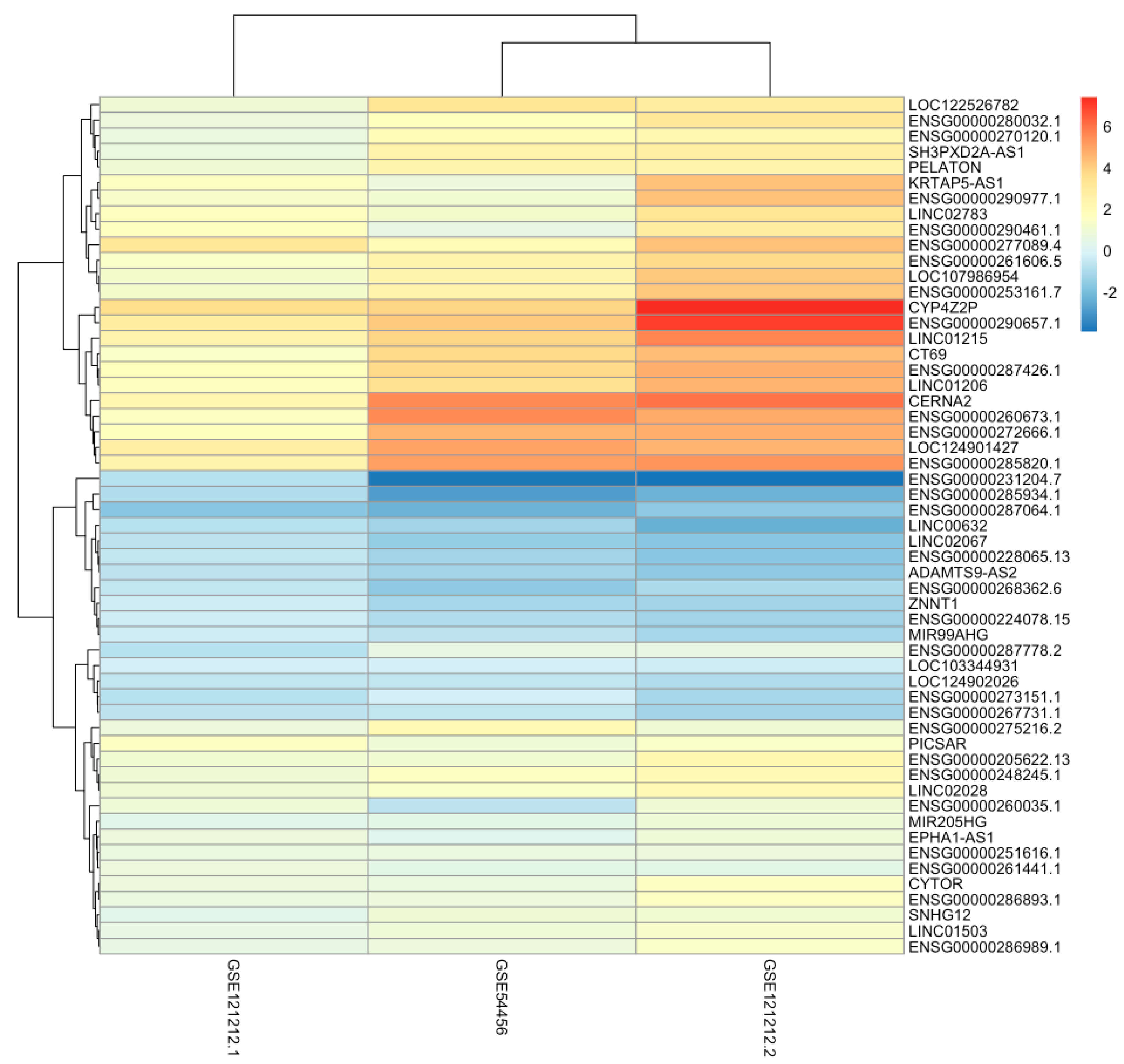
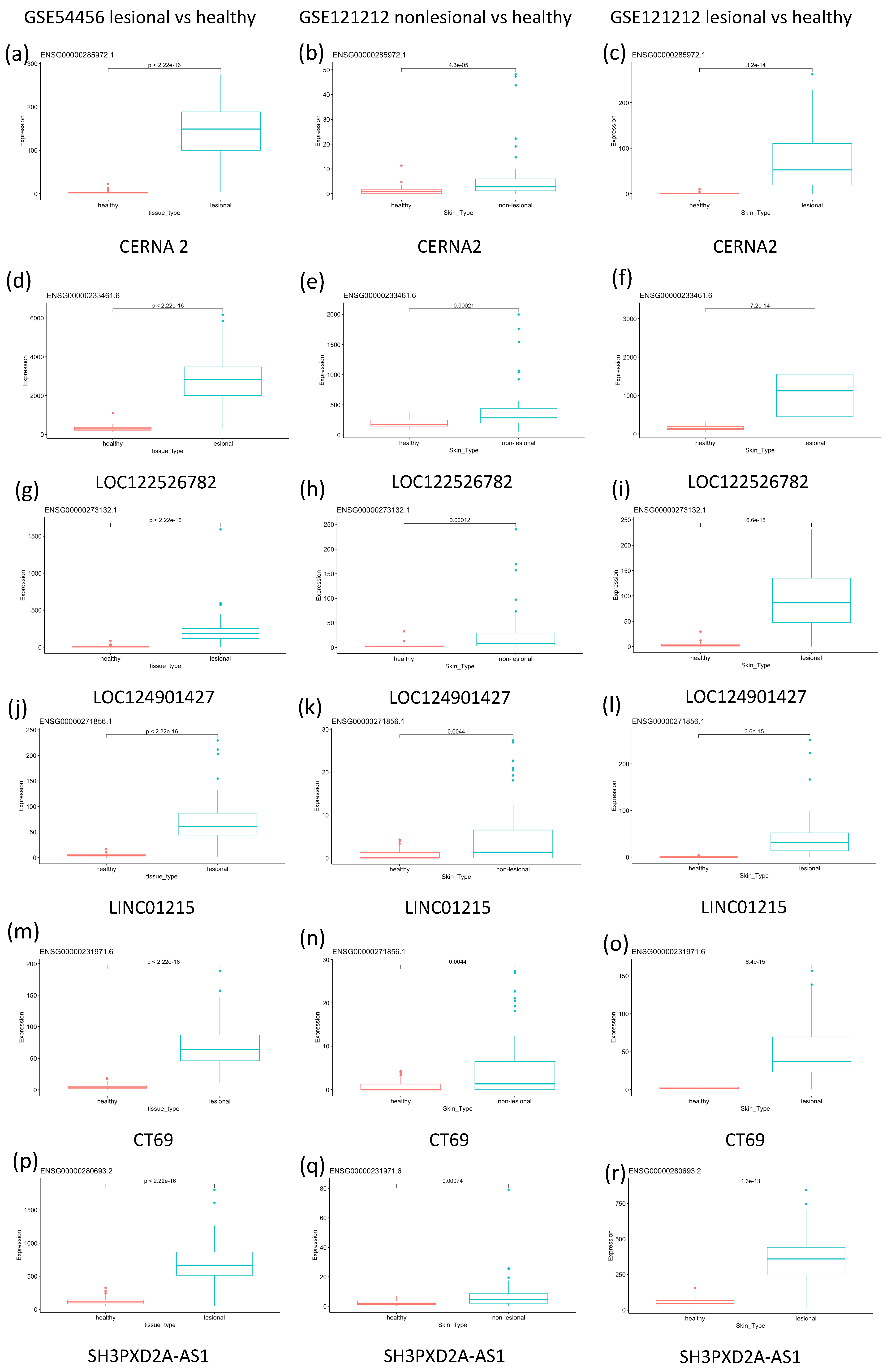

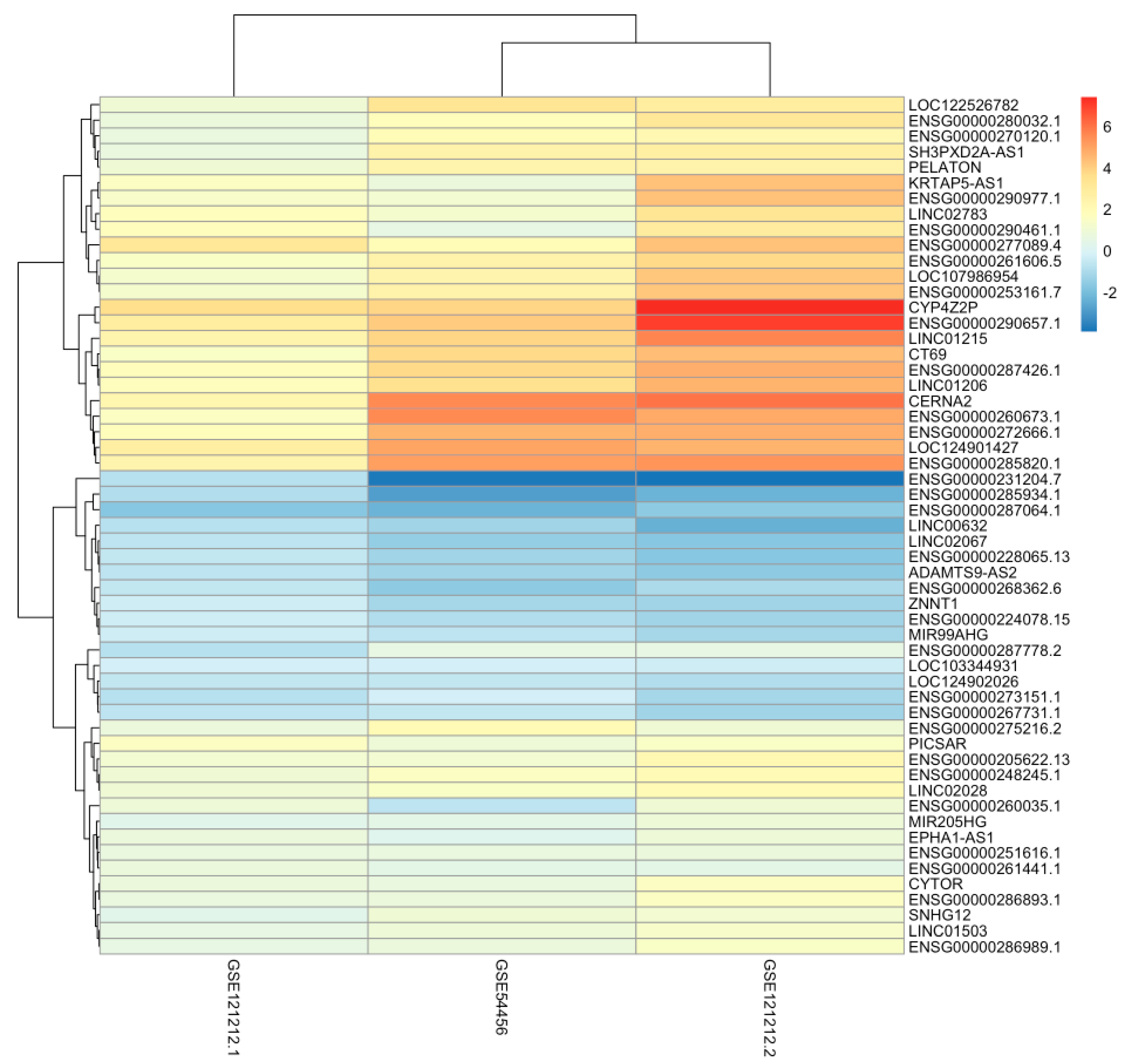{kind=link}
{kind=link}
| Gene | logFC | p-Value | FDR | Symbol | Gene Name |
|---|---|---|---|---|---|
| ENSG00000285972.1 | 5.54 | 0 | 0 | CERNA2 | competing endogenous lncRNA 2 for microRNA let-7b |
| ENSG00000233461.6 | 3.26 | 1.72 × 10−271 | 9.03 × 10−268 | LOC122526782 | uncharacterised LOC122526782 |
| ENSG00000260673.1 | 5.60 | 4.93 × 10−245 | 1.73 × 10−241 | lnc-RPP40-3 | NA |
| ENSG00000214999.3 | 2.56 | 1.32 × 10−237 | 3.47 × 10−234 | ALOX12-AS1 | ALOX12 antisense 1 |
| ENSG00000280693.2 | 2.54 | 6.90 × 10−197 | 1.45 × 10−193 | SH3PXD2A-AS1 | SH3PXD2A antisense RNA 1 |
| ENSG00000271856.1 | 3.87 | 3.29 × 10−195 | 5.76 × 10−192 | LINC01215 | long intergenic non-protein-coding RNA 1215 |
| ENSG00000272666.1 | 4.61 | 1.20 × 10−189 | 1.80 × 10−186 | KLHDC7B-DT | KLHDC7B Divergent Transcript |
| ENSG00000231971.6 | 3.73 | 4.45 × 10−187 | 5.84 × 10−184 | CT69 | cancer/testis associated transcript 69 |
| ENSG00000273272.2 | 4.41 | 4.97 × 10−164 | 5.81 × 10−161 | NA | NA |
| ENSG00000286962.1 | 4.34 | 1.61 × 10−162 | 1.70 × 10−159 | SLC6A11-AS1 | antisense to SLC6A11 |
| ENSG00000253417.5 | 3.69 | 3.41 × 10−142 | 3.26 × 10−139 | LINC02159 | long intergenic non-protein-coding RNA 2159 |
| ENSG00000273132.1 | 5.01 | 1.59 × 10−135 | 1.39 × 10−132 | LOC124901427 | uncharacterised LOC124901427 |
| ENSG00000285820.1 | 5.05 | 6.36 × 10−127 | 5.14 × 10−124 | ASTN2-AS1 | antisense to ASTN2 |
| ENSG00000251320.1 | −2.78 | 2.97 × 10−126 | 2.23 × 10−123 | NA | NA |
| ENSG00000275880.1 | 1.68 | 1.19 × 10−123 | 8.35 × 10−121 | NAXD-AS1 | NAXD antisense RNA 1 |
| ENSG00000272711.1 | 1.97 | 1.89 × 10−119 | 1.24 × 10−116 | HK2-DT | HK2 Divergent Transcript |
| ENSG00000254027.1 | 3.32 | 4.02 × 10−117 | 2.49 × 10−114 | LNMICC | NA |
| ENSG00000258689.2 | 5.27 | 1.11 × 10−115 | 6.45 × 10−113 | LINC01269 | long intergenic non-protein-coding RNA 1269 |
| ENSG00000245648.2 | 2.41 | 7.96 × 10−113 | 4.40 × 10−110 | KLRK1-AS1 | KLRK1 antisense RNA 1 |
| ENSG00000288575.1 | 3.62 | 1.16 × 10−110 | 6.08 × 10−108 | NA | NA |
| Gene | logFC | p-Value | FDR | Symbol | Gene Name |
|---|---|---|---|---|---|
| ENSG00000273132.1 | 2.67 | 2.56 × 10−10 | 1.24 × 10−6 | LOC124901427 | uncharacterised LOC124901427 |
| ENSG00000229153.7 | 0.81 | 1.09 × 10−8 | 2.63 × 10−5 | EPHA1-AS1 | EPHA1 antisense RNA 1 |
| ENSG00000233461.6 | 1.03 | 4.20 × 10−8 | 5.32 × 10−5 | LOC122526782 | uncharacterised LOC122526782 |
| ENSG00000285972.1 | 2.33 | 4.40 × 10−8 | 5.32 × 10−5 | CERNA2 | competing endogenous lncRNA 2 for microRNA let-7b |
| ENSG00000277089.4 | 3.13 | 1.89 × 10−7 | 1.83 × 10−4 | CCL3-AS1 | CCL3 antisense RNA 1 |
| ENSG00000231971.6 | 1.52 | 6.17 × 10−7 | 4.97 × 10−4 | CT69 | cancer/testis associated transcript 69 |
| ENSG00000204362.6 | 1.68 | 9.28 × 10−6 | 3.53 × 10−3 | LINC02783 | long intergenic non-protein-coding RNA 2783 |
| ENSG00000234678.2 | 1.41 | 9.89 × 10−6 | 3.53 × 10−3 | ELF3-AS1 | ELF3 antisense RNA 1 |
| ENSG00000234862.1 | 2.30 | 1.02 × 10−5 | 3.53 × 10−3 | NA | NA |
| ENSG00000240401.9 | 1.12 | 7.07 × 10−6 | 3.53 × 10−3 | NA | NA |
| ENSG00000260673.1 | 1.65 | 6.99 × 10−6 | 3.53 × 10−3 | NA | NA |
| ENSG00000270120.1 | 0.71 | 9.73 × 10−6 | 3.53 × 10−3 | NA | NA |
| ENSG00000271856.1 | 2.34 | 6.35 × 10−6 | 3.53 × 10−3 | LINC01215 | long intergenic non-protein-coding RNA 1215 |
| ENSG00000272666.1 | 1.78 | 8.06 × 10−6 | 3.53 × 10−3 | KLHDC7B-DT | KLHDC7B divergent transcript |
| ENSG00000287426.1 | 1.81 | 1.13 × 10−5 | 3.65 × 10−3 | NA | NA |
| ENSG00000222041.13 | 0.82 | 1.29 × 10−5 | 3.91 × 10−3 | CYTOR | cytoskeleton regulator RNA |
| ENSG00000203930.13 | −0.76 | 1.43 × 10−5 | 4.01 × 10−3 | LINC00632 | long intergenic non-protein-coding RNA 632 |
| ENSG00000287927.1 | 1.28 | 2.22 × 10−5 | 5.96 × 10−3 | LOC107986954 | uncharacterised LOC107986954 |
| ENSG00000231204.7 | −0.75 | 2.64 × 10−5 | 6.72 × 10−3 | NA | NA |
| ENSG00000253161.7 | 1.24 | 2.87 × 10−5 | 6.95 × 10−3 | LINC01605 | long intergenic non-protein-coding RNA 1605 |
| Gene | logFC | p-Value | FDR | Symbol | Gene Name |
|---|---|---|---|---|---|
| ENSG00000253161.7 | 4.24 | 7.46 × 10−82 | 5.54 × 10−78 | LINC01605 | long intergenic non-protein-coding RNA 1605 |
| ENSG00000273132.1 | 4.66 | 2.92 × 10−70 | 1.09 × 10−66 | LOC124901427 | uncharacterised LOC124901427 |
| ENSG00000287426.1 | 4.74 | 2.26 × 10−65 | 5.59 × 10−62 | LOC124902793 | NA |
| ENSG00000255774.2 | −2.60 | 3.26 × 10−65 | 6.06 × 10−62 | LINC02747 | long intergenic non-protein-coding RNA 2747 |
| ENSG00000272666.1 | 4.73 | 1.03 × 10−63 | 1.53 × 10−60 | KLHDC7B-DT | KLHDC7B divergent transcript |
| ENSG00000231204.7 | −3.89 | 1.15 × 10−61 | 1.27 × 10−58 | NA | NA |
| ENSG00000260673.1 | 4.83 | 1.20 × 10−61 | 1.27 × 10−58 | NA | NA |
| ENSG00000280693.2 | 2.70 | 3.47 × 10−59 | 3.22 × 10−56 | SH3PXD2A-AS1 | SH3PXD2A antisense RNA 1 |
| ENSG00000285972.1 | 5.97 | 1.74 × 10−58 | 1.43 × 10−55 | CERNA2 | competing endogenous lncRNA 2 for microRNA let-7b |
| ENSG00000231971.6 | 4.44 | 3.93 × 10−57 | 2.92 × 10−54 | CT69 | cancer/testis associated transcript 69 |
| ENSG00000287563.1 | −3.75 | 1.35 × 10−56 | 9.14 × 10−54 | NA | NA |
| ENSG00000249859.14 | 1.66 | 1.12 × 10−55 | 6.91 × 10−53 | PVT1 | Pvt1 oncogene |
| ENSG00000280032.1 | 3.23 | 4.09 × 10−54 | 2.34 × 10−51 | NA | NA |
| ENSG00000233461.6 | 2.85 | 8.99 × 10−54 | 4.77 × 10−51 | LOC122526782 | uncharacterised LOC122526782 |
| ENSG00000233452.9 | 2.50 | 2.63 × 10−51 | 1.30 × 10−48 | STXBP5-AS1 | STXBP5 antisense RNA 1 |
| ENSG00000271856.1 | 5.64 | 6.34 × 10−51 | 2.94 × 10−48 | LINC01215 | long intergenic non-protein-coding RNA 1215 |
| ENSG00000253746.1 | 4.19 | 2.23 × 10−49 | 9.75 × 10−47 | NA | NA |
| ENSG00000290657.1 | 7.08 | 8.04 × 10−49 | 3.32 × 10−46 | NA | NA |
| ENSG00000223863.1 | −1.79 | 2.64 × 10−48 | 1.03 × 10−45 | LINC01805 | long intergenic non-protein-coding RNA 1805 |
| ENSG00000234678.2 | 3.45 | 5.81 × 10−44 | 2.1 × 10−41 | ELF3-AS1 | ELF3 antisense RNA 1 |
| Gene | Symbol | Gene Name | Regulation |
|---|---|---|---|
| ENSG00000273132.1 | LOC124901427 | uncharacterised LOC124901427 | Upregulated |
| ENSG00000229153.7 | EPHA1-AS1 | EPHA1 antisense RNA 1 | Upregulated |
| ENSG00000233461.6 | LOC122526782 | uncharacterised LOC122526782 | Upregulated |
| ENSG00000285972.1 | CERNA2 | competing endogenous lncRNA 2 for microRNA let-7b | Upregulated |
| ENSG00000277089.4 | CCL3-AS1 | CCL3 antisense RNA 1 | Upregulated |
| ENSG00000231971.6 | CT69 | cancer/testis associated transcript 69 | Upregulated |
| ENSG00000204362.6 | LINC02783 | long intergenic non-protein-coding RNA 2783 | Upregulated |
| ENSG00000260673.1 | NA | NA | Upregulated |
| ENSG00000270120.1 | NA | NA | Upregulated |
| ENSG00000271856.1 | LINC01215 | long intergenic non-protein-coding RNA 1215 | Upregulated |
| ENSG00000272666.1 | KLHDC7B-DT | KLHDC7B divergent transcript | Upregulated |
| ENSG00000287426.1 | LOC124902793 | uncharacterised LOC124902793 | Upregulated |
| ENSG00000222041.13 | CYTOR | cytoskeleton regulator RNA | Upregulated |
| ENSG00000203930.13 | LINC00632 | long intergenic non-protein-coding RNA 632 | Downregulated |
| ENSG00000287927.1 | LOC107986954 | uncharacterised LOC107986954 | Upregulated |
| ENSG00000231204.7 | NA | NA | Downregulated |
| ENSG00000253161.7 | LINC01605 | long intergenic non-protein-coding RNA 1605 | Upregulated |
| ENSG00000251616.1 | RP11-485M7.3 | NA | Upregulated |
| ENSG00000275874.1 | PICSAR | P38 inhibited cutaneous squamous cell carcinoma associated lincRNA | Upregulated |
| ENSG00000228065.13 | LINC01515 | long intergenic non-protein-coding RNA 1515 | Downregulated |
| ENSG00000230937.13 | MIR205HG | MIR205 host gene | Upregulated |
| ENSG00000233901.8 | LINC01503 | long intergenic non-protein-coding RNA 1503 | Upregulated |
| ENSG00000240567.1 | LINC02067 | long intergenic non-protein-coding RNA 2067 | Downregulated |
| ENSG00000275216.2 | LINC03061 | long intergenic non-protein-coding RNA 3061 | Upregulated |
| ENSG00000280693.2 | SH3PXD2A-AS1 | SH3PXD2A antisense RNA 1 | Upregulated |
| ENSG00000248245.1 | NA | NA | Upregulated |
| ENSG00000286893.1 | NA | NA | Upregulated |
| ENSG00000286989.1 | NA | NA | Upregulated |
| ENSG00000285820.1 | ASTN2-AS1 | antisense to ASTN2 | Upregulated |
| ENSG00000261441.1 | POLG-DT | POLG Divergent Transcript | Upregulated |
| ENSG00000290461.1 | NA | NA | Upregulated |
| ENSG00000224397.9 | PELATON | plaque enriched lncRNA in atherosclerotic and inflammatory bowel macrophage regulation | Upregulated |
| ENSG00000273151.1 | NA | NA | Downregulated |
| ENSG00000290456.1 | CYP4Z2P | cytochrome P450 family 4 subfamily Z member 2, pseudogene | Upregulated |
| ENSG00000267731.1 | NA | NA | Downregulated |
| ENSG00000224078.15 | SNHG14 | small nucleolar RNA host gene 14 | Downregulated |
| ENSG00000280032.1 | NA | NA | Upregulated |
| ENSG00000215386.15 | MIR99AHG | mir-99a-let-7c cluster host gene | Downregulated |
| ENSG00000261087.1 | ZNNT1 | ZNF706 neighbouring transcript 1 | Downregulated |
| ENSG00000290657.1 | NA | NA | Upregulated |
| ENSG00000268362.6 | NA | NA | Downregulated |
| ENSG00000285934.1 | NA | NA | Downregulated |
| ENSG00000230102.8 | LINC02028 | long intergenic non-protein-coding RNA 2028 | Upregulated |
| ENSG00000260917.1 | LOC103344931 | uncharacterised LOC103344931 | Downregulated |
| ENSG00000233930.4 | KRTAP5-AS1 | KRTAP5-1/KRTAP5-2 antisense RNA 1 | Upregulated |
| ENSG00000287778.2 | NA | NA | Downregulated |
| ENSG00000197989.15 | SNHG12 | small nucleolar RNA host gene 12 | Upregulated |
| ENSG00000241684.8 | ADAMTS9-AS2 | ADAMTS9 antisense RNA 2 | Downregulated |
| ENSG00000261606.5 | NA | NA | Upregulated |
| ENSG00000287064.1 | NA | NA | Downregulated |
| ENSG00000286535.1 | LOC124902026 | uncharacterised LOC124902026 | Downregulated |
| ENSG00000205622.13 | NA | NA | Upregulated |
| ENSG00000242512.9 | LINC01206 | long intergenic non-protein-coding RNA 1206 | Upregulated |
| ENSG00000290977.1 | NA | NA | Upregulated |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stacey, V.M.; Kõks, S. Genome-Wide Differential Transcription of Long Noncoding RNAs in Psoriatic Skin. Int. J. Mol. Sci. 2023, 24, 16344. https://doi.org/10.3390/ijms242216344
Stacey VM, Kõks S. Genome-Wide Differential Transcription of Long Noncoding RNAs in Psoriatic Skin. International Journal of Molecular Sciences. 2023; 24(22):16344. https://doi.org/10.3390/ijms242216344
Chicago/Turabian StyleStacey, Valerie M., and Sulev Kõks. 2023. "Genome-Wide Differential Transcription of Long Noncoding RNAs in Psoriatic Skin" International Journal of Molecular Sciences 24, no. 22: 16344. https://doi.org/10.3390/ijms242216344
APA StyleStacey, V. M., & Kõks, S. (2023). Genome-Wide Differential Transcription of Long Noncoding RNAs in Psoriatic Skin. International Journal of Molecular Sciences, 24(22), 16344. https://doi.org/10.3390/ijms242216344