
Host Genetic Background Effect on Body Weight Changes Influenced by Heterozygous Smad4 Knockout Using Collaborative Cross Mouse Population
 , ,
, , 

Abstract
:1. Introduction
2. Results
2.1. Generation of F1 (Smad4x CC) Mice
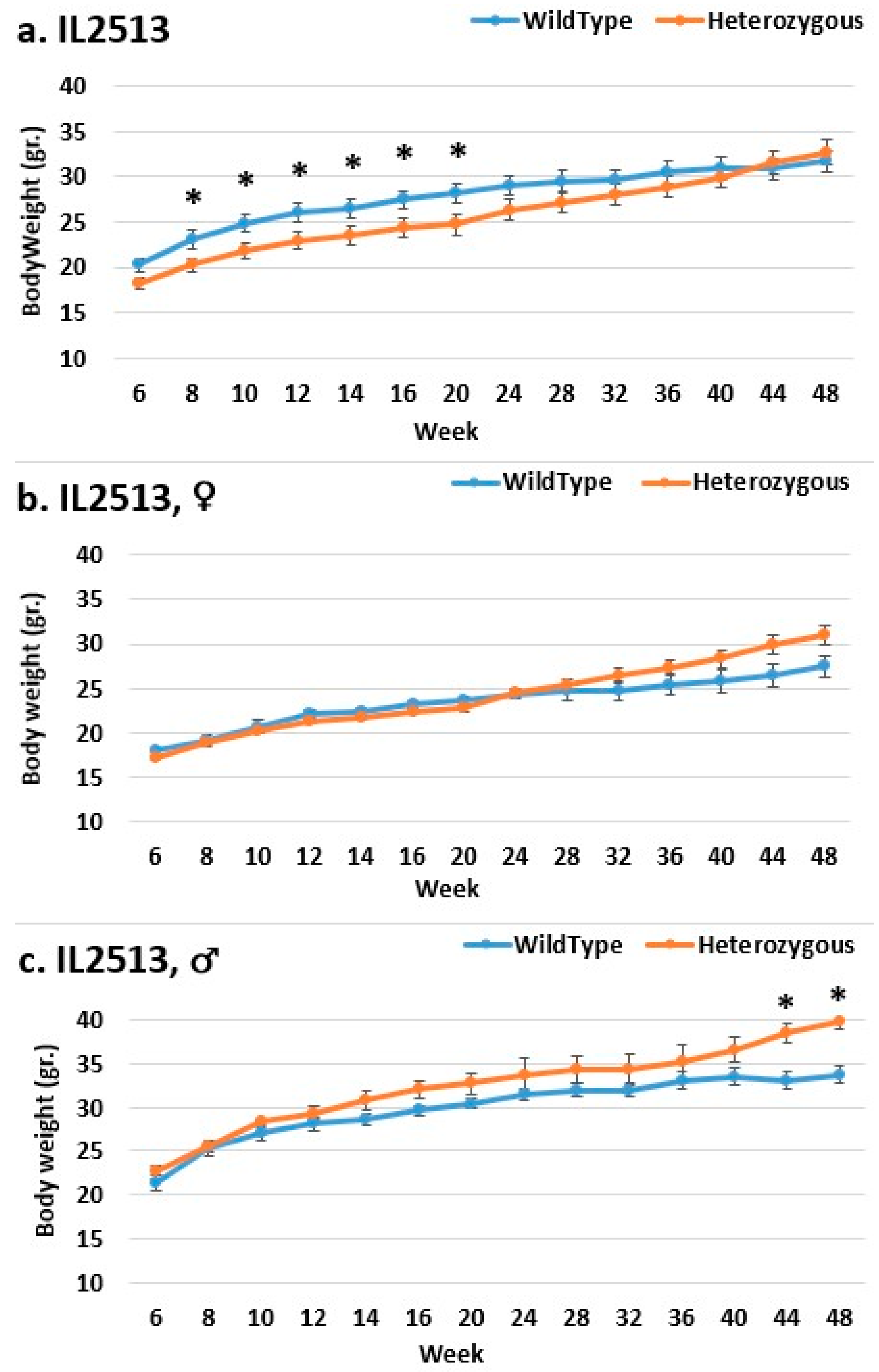
2.2. Dynamics of Body Weight (BW) Changes during the Experimental Period for F1 (Smad4x CC) Mice
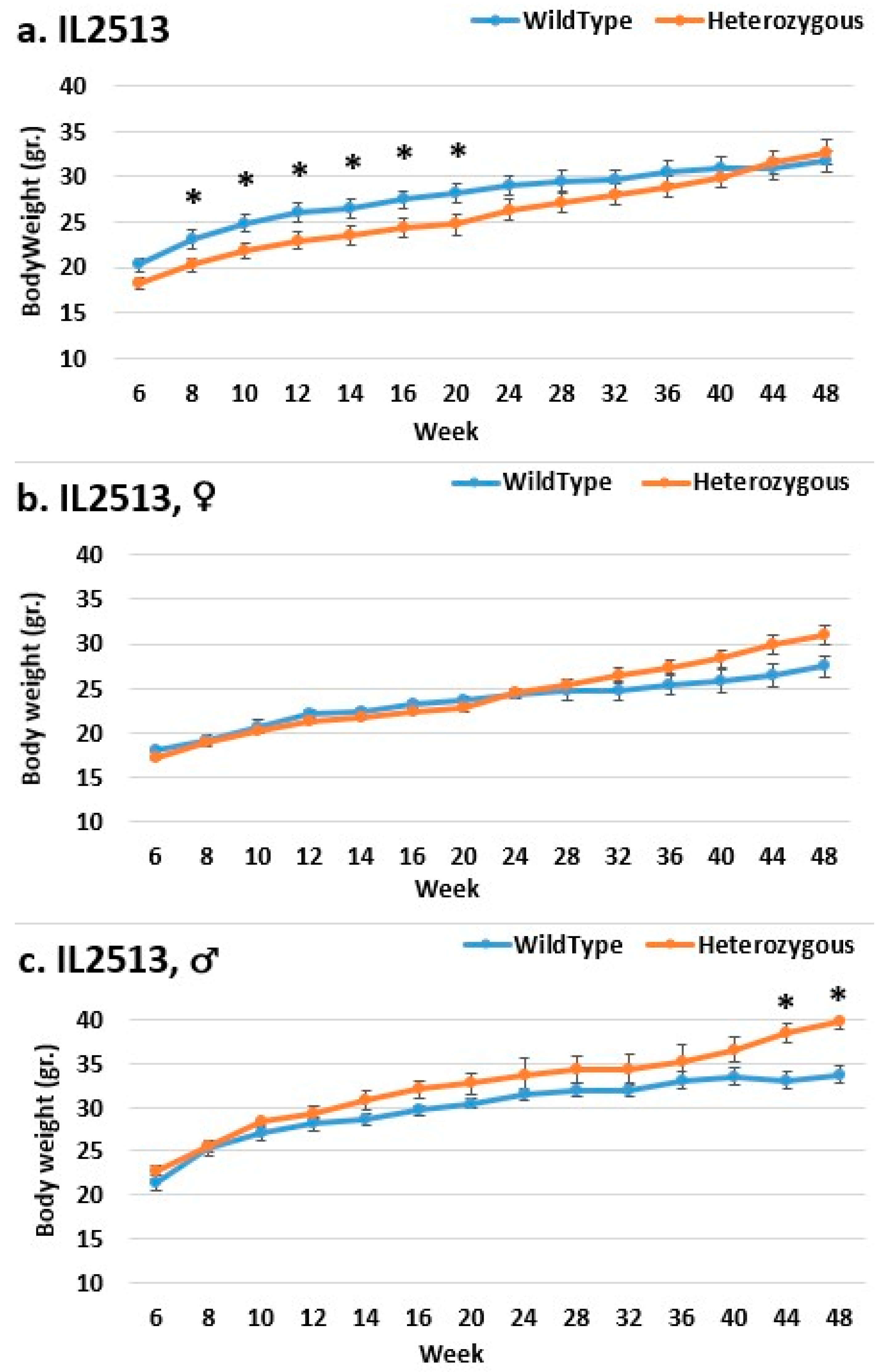
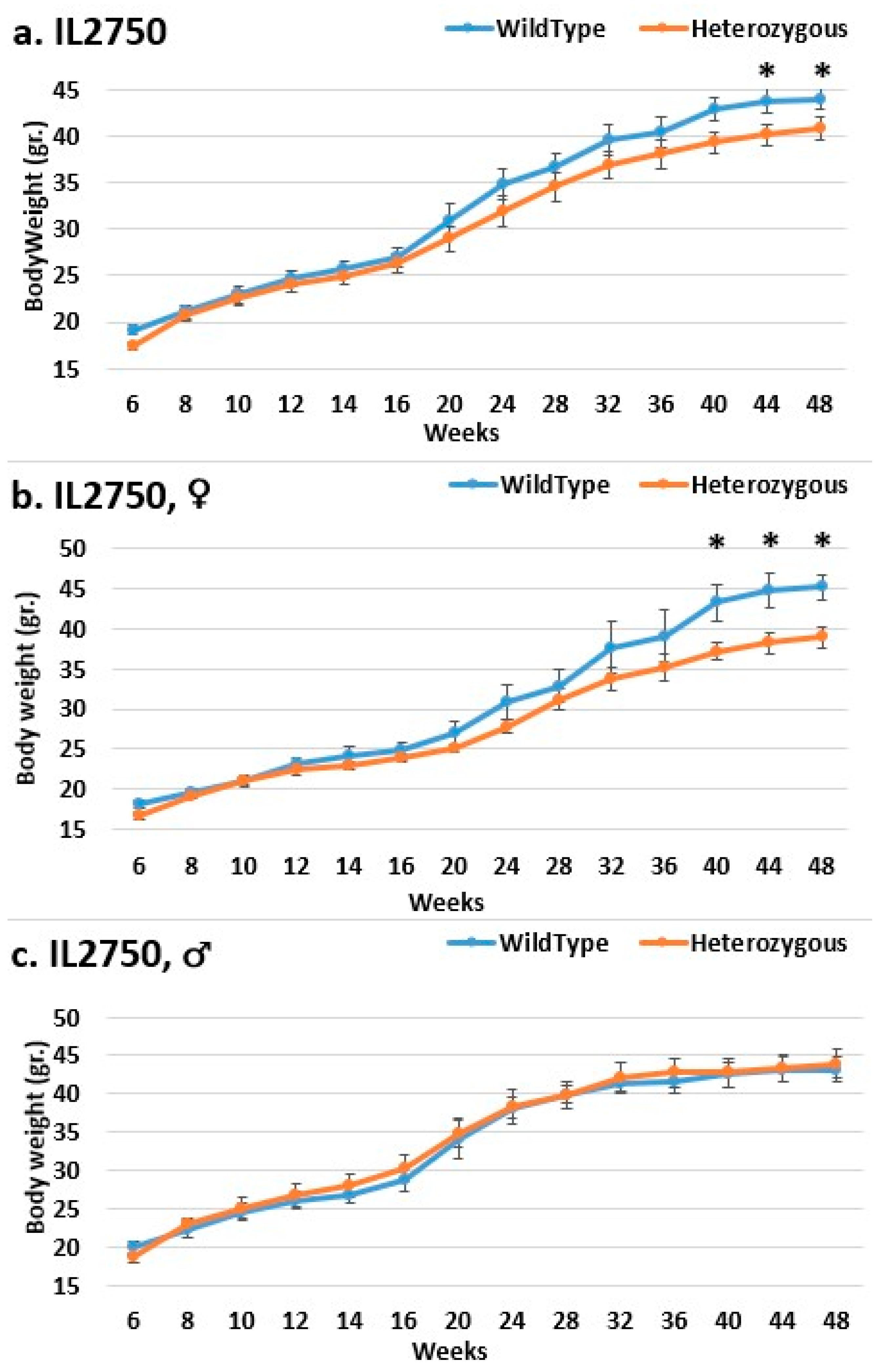
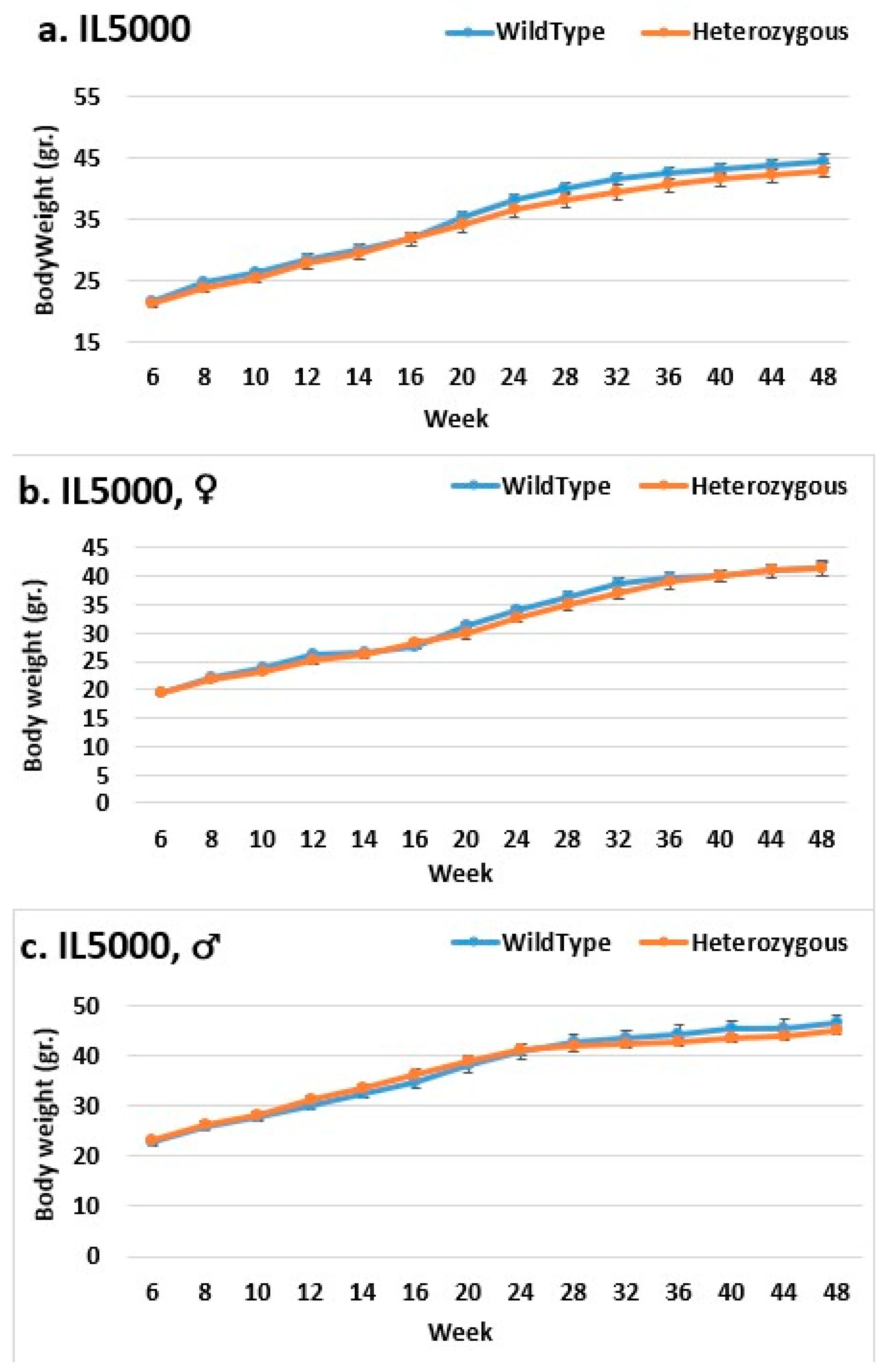
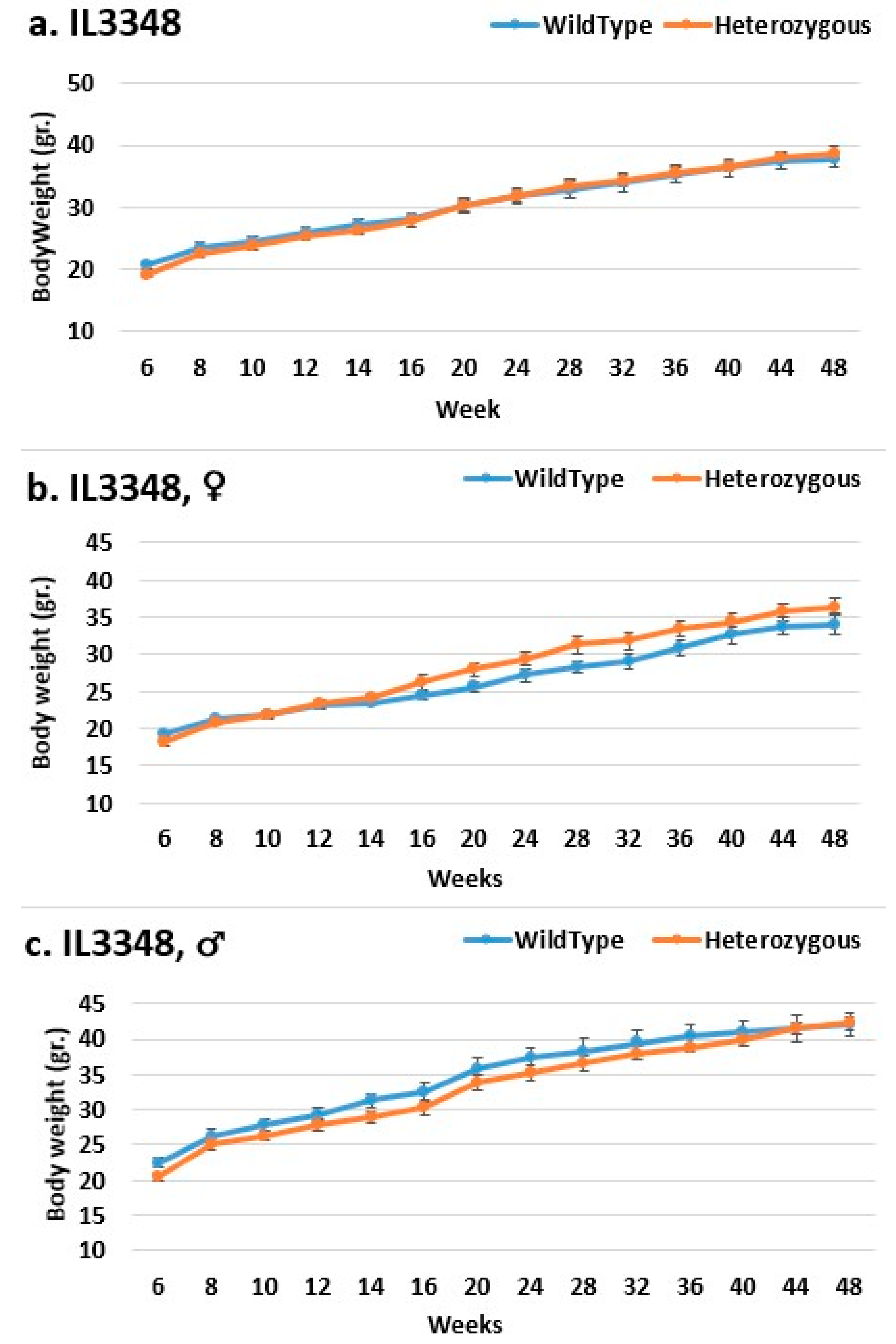
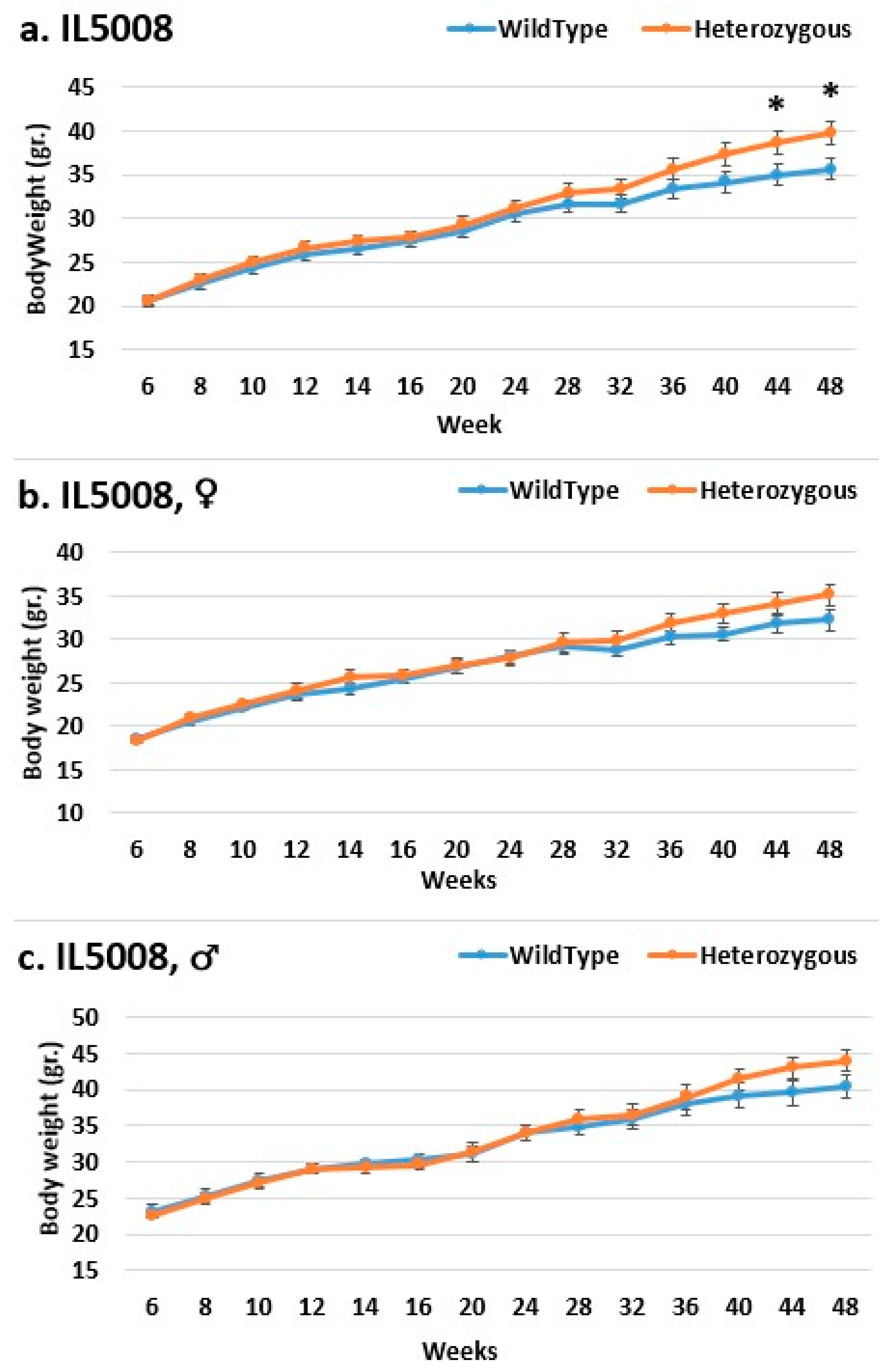
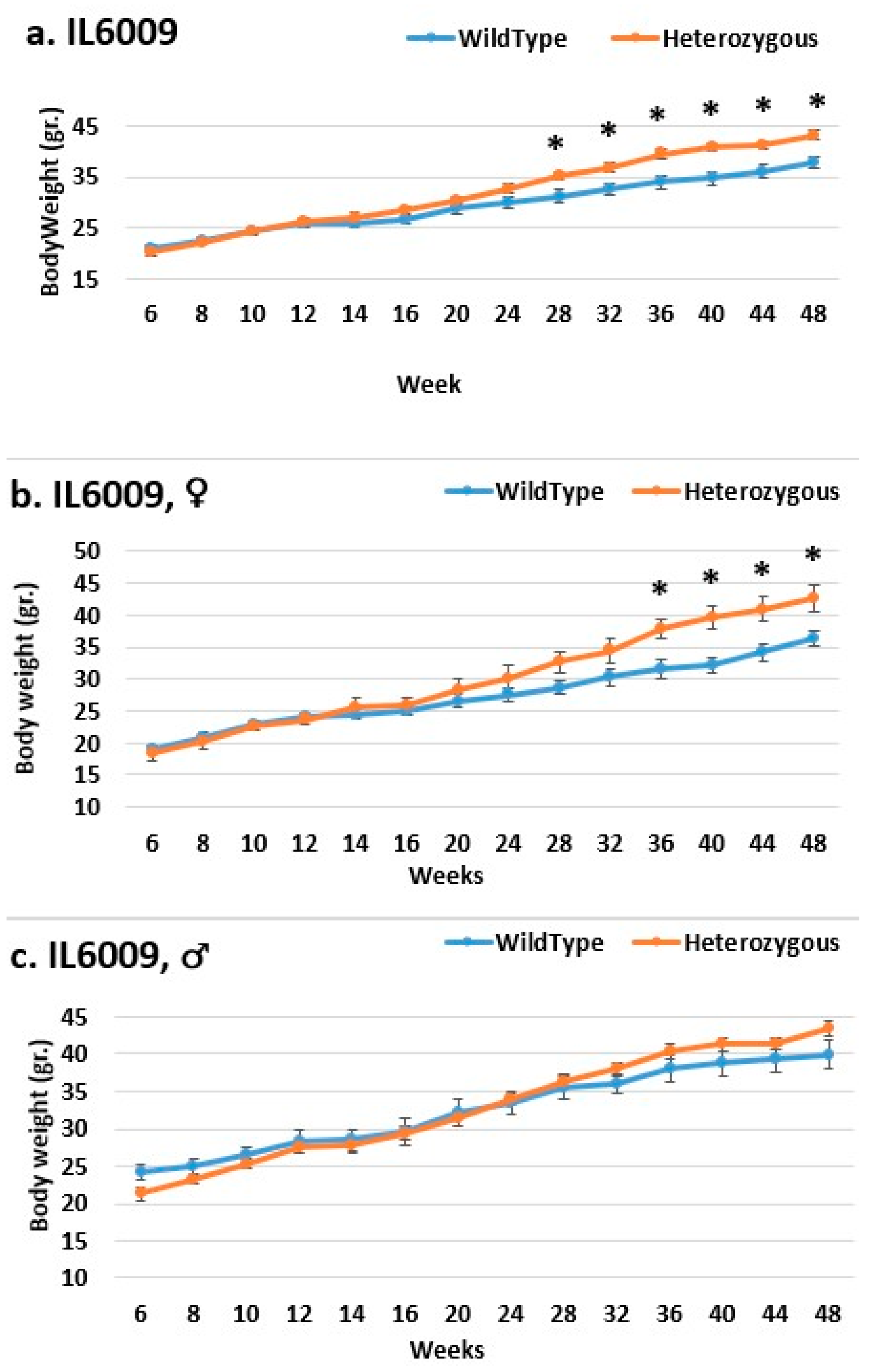
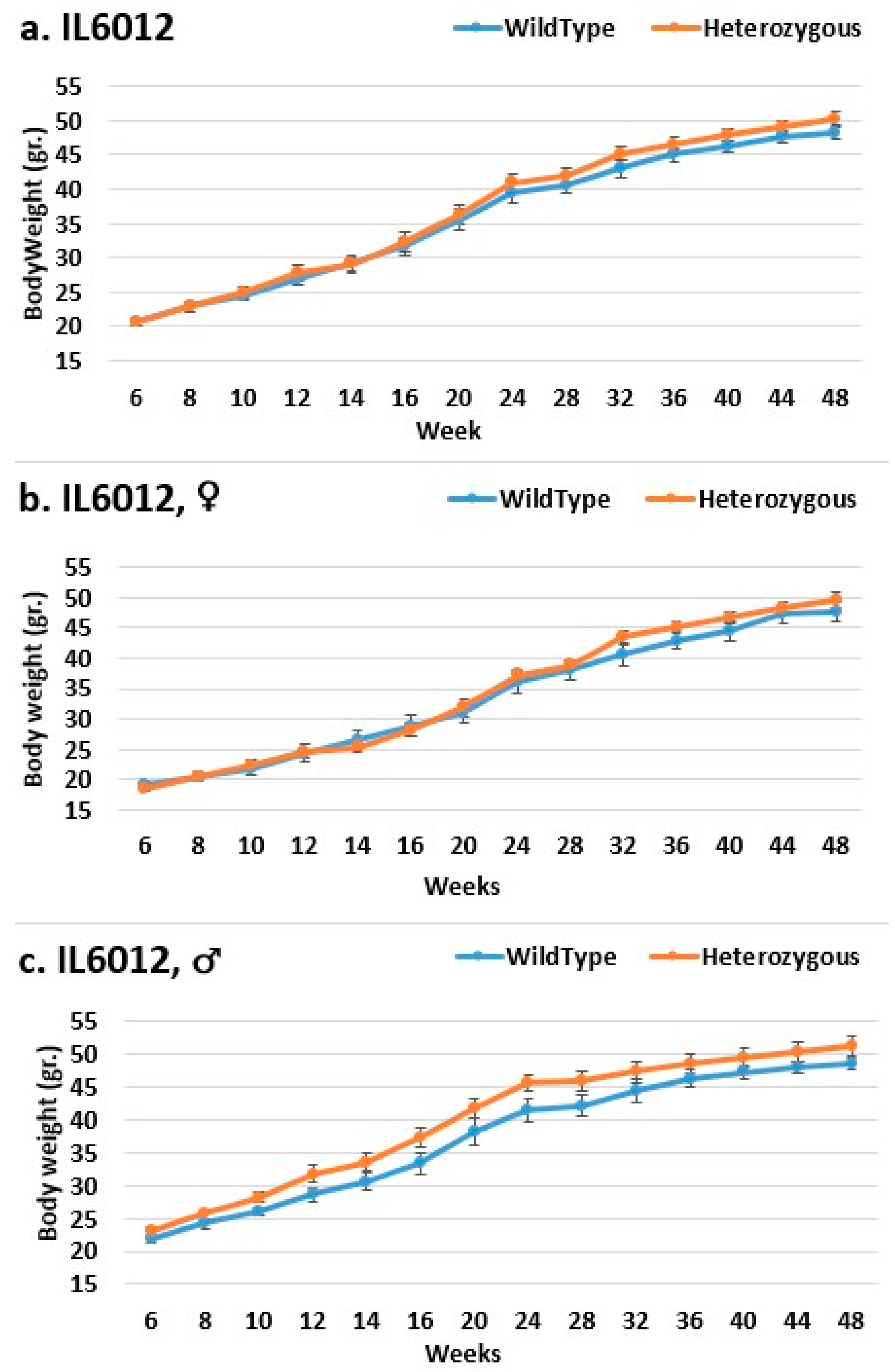
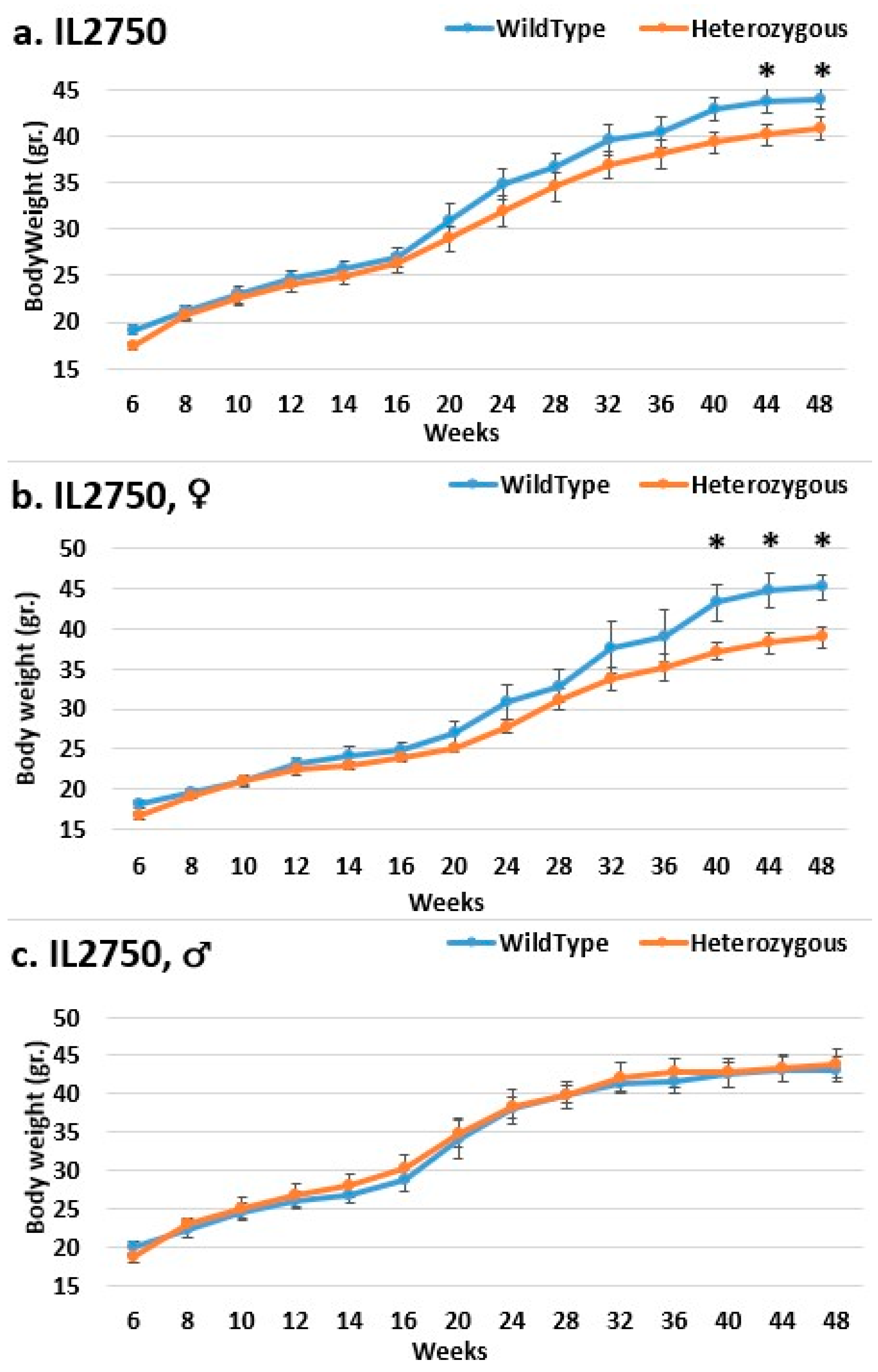
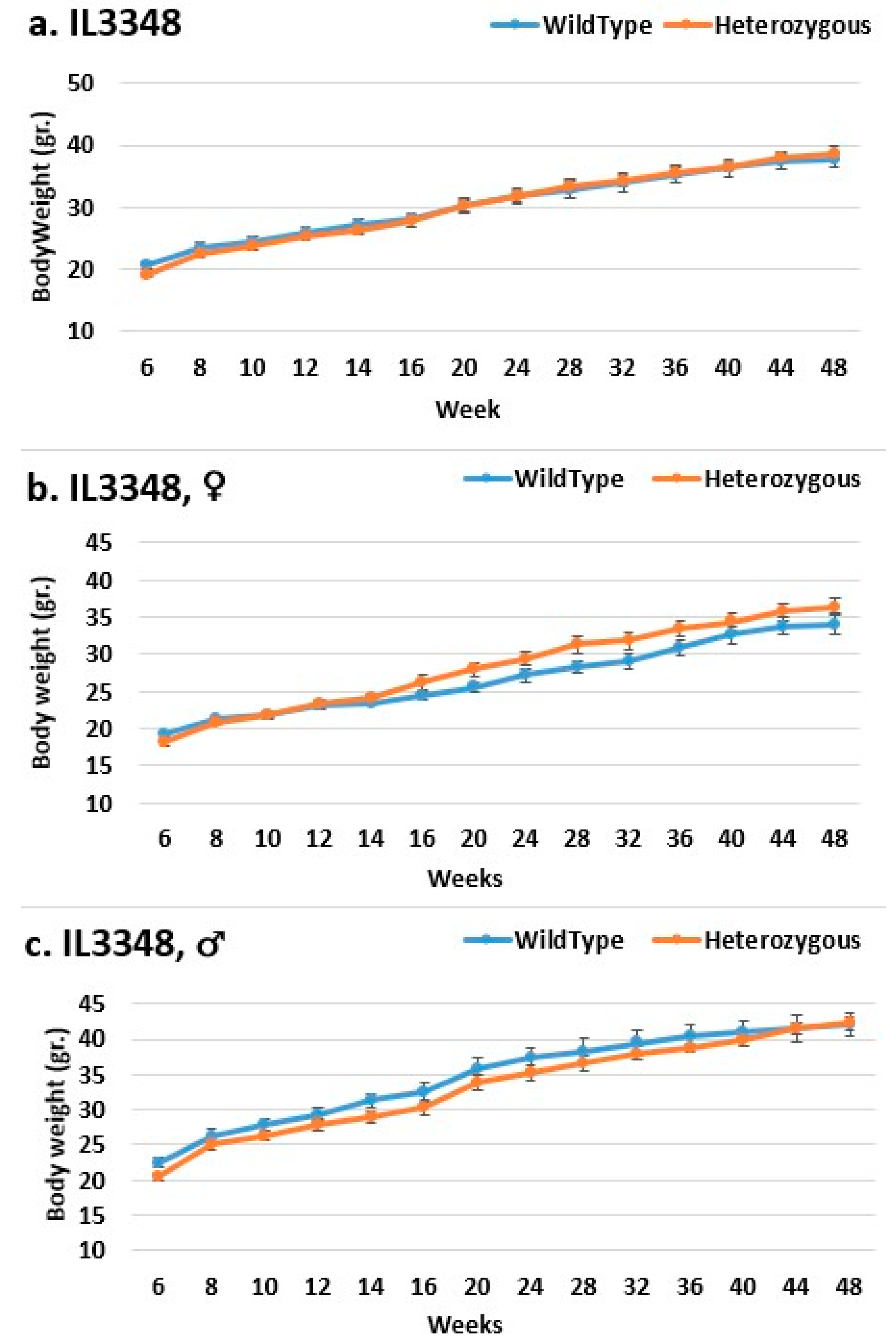
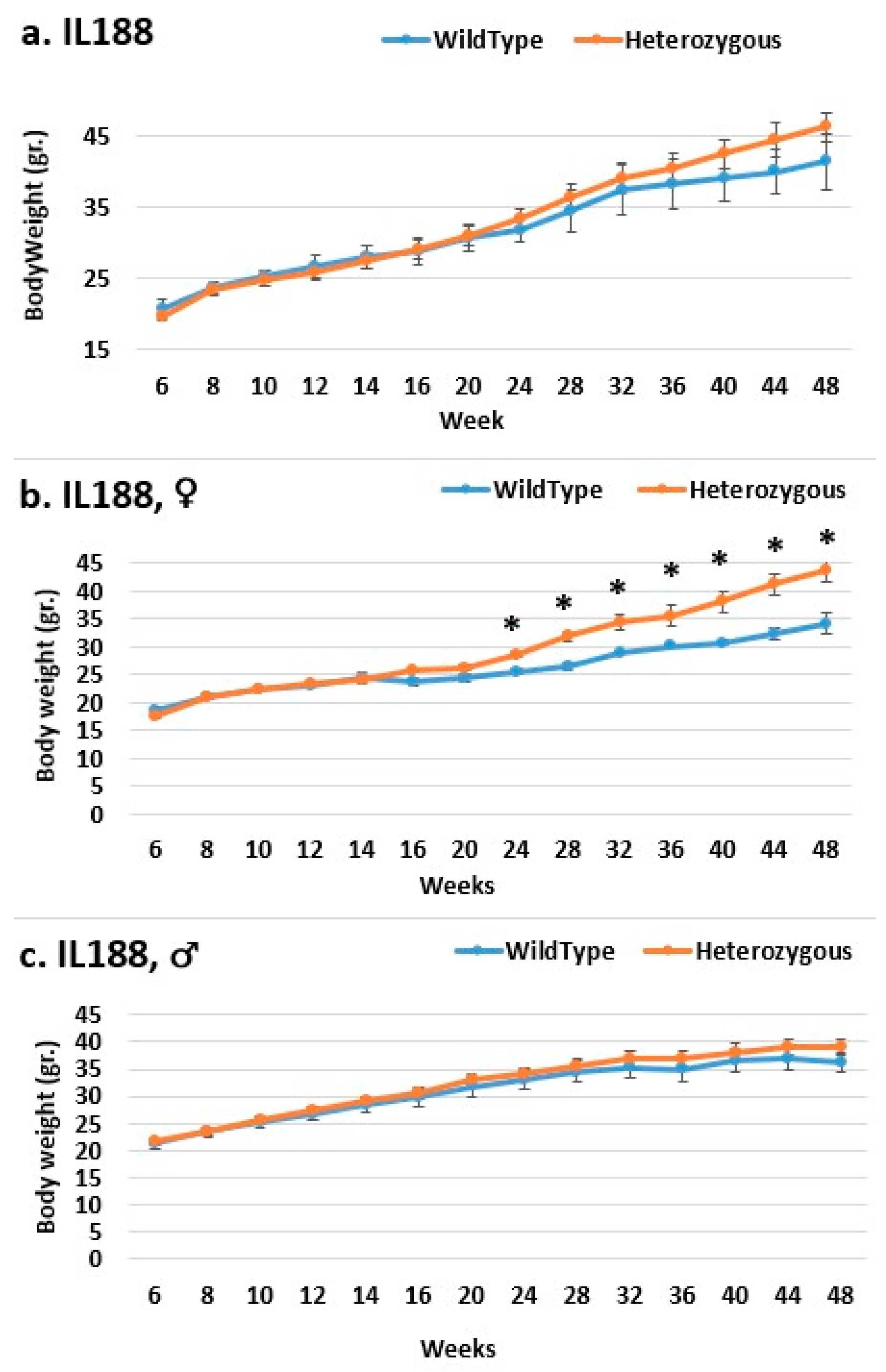
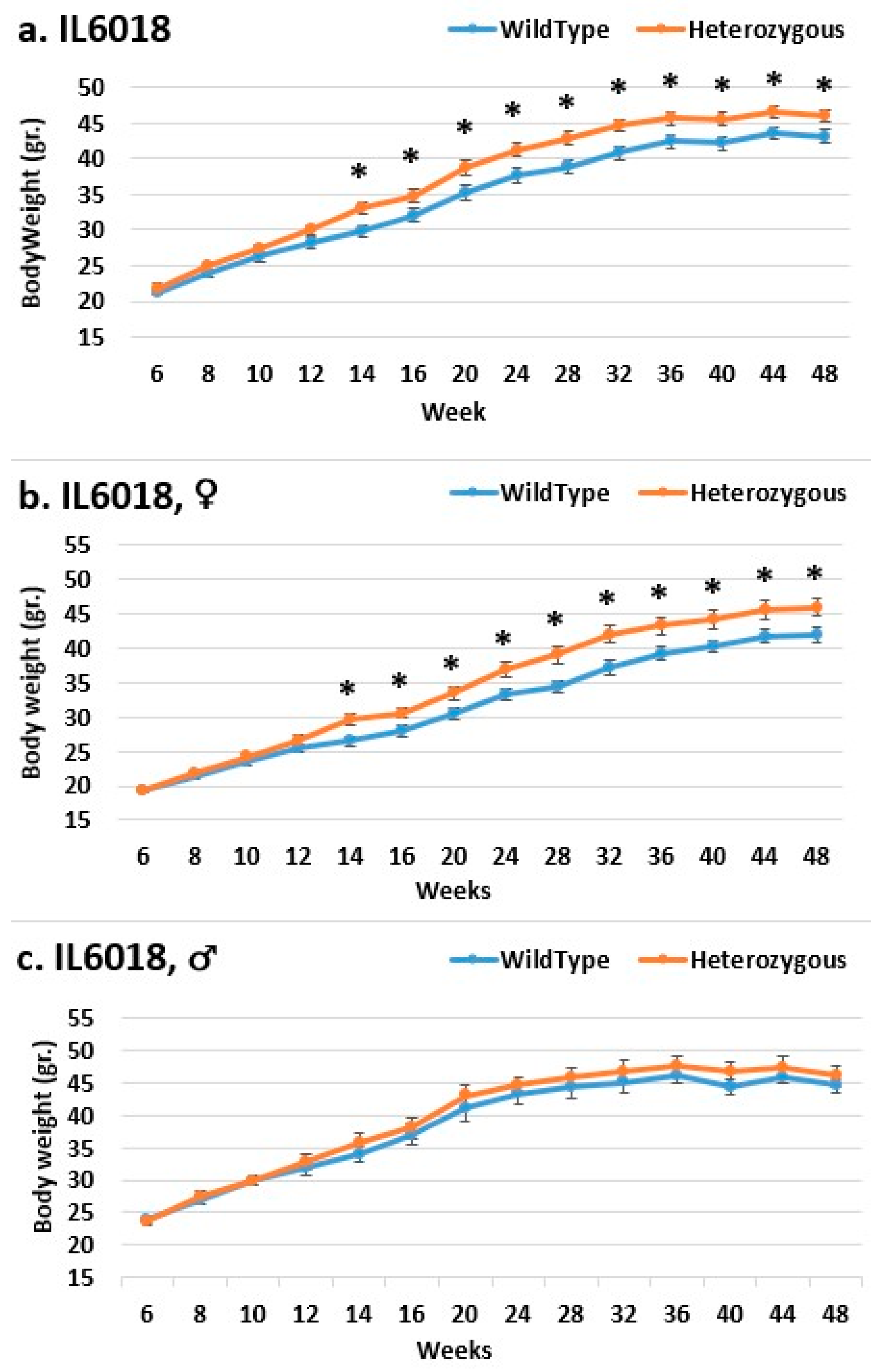
2.3. Dynamics of BW Changes in Grams over the 48-Week Experiment for Each CC Line
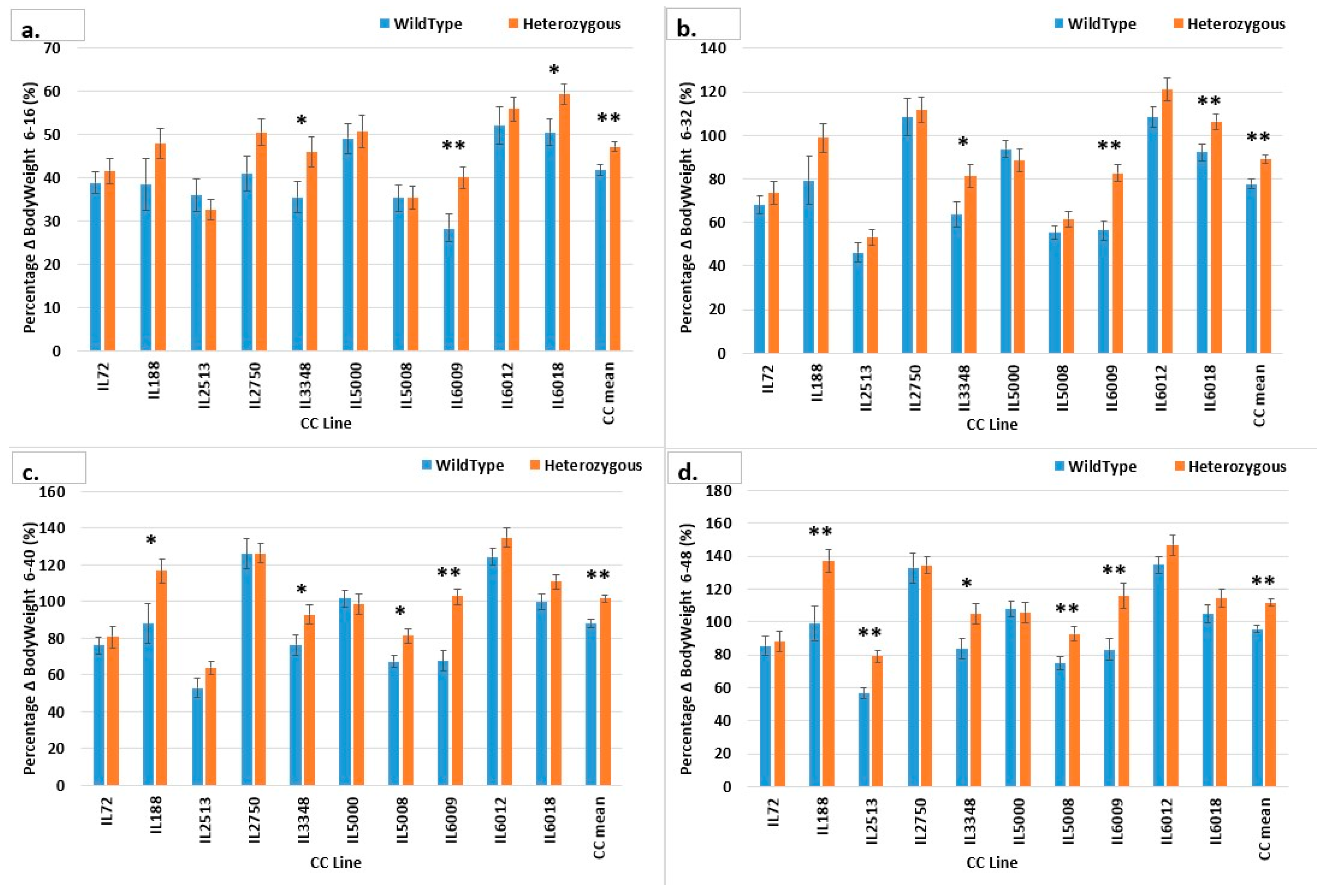
2.4. Percentage of BW Gain (g) of CC Lines after 48 Weeks
2.5. Computational Methods
2.5.1. Heritability
2.5.2. Regression Models
2.5.3. Model Details
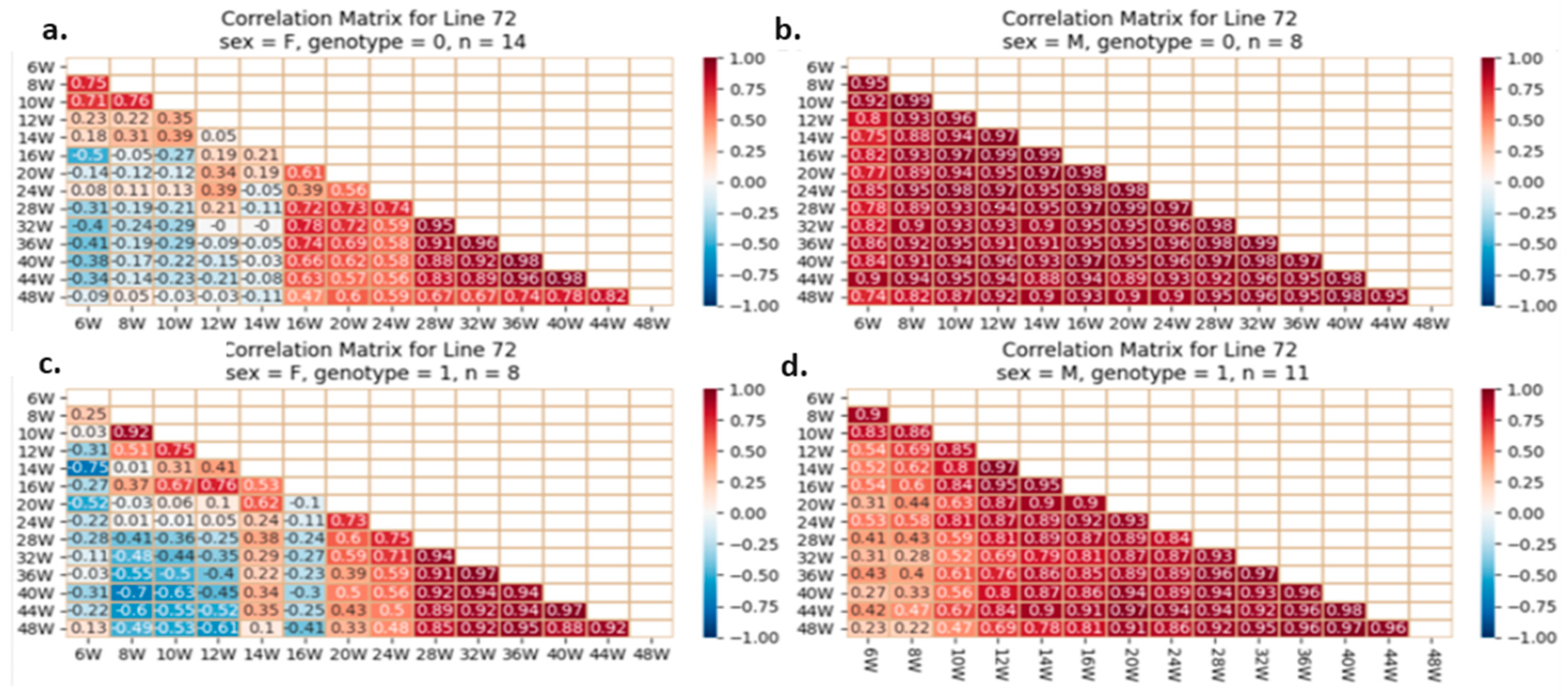
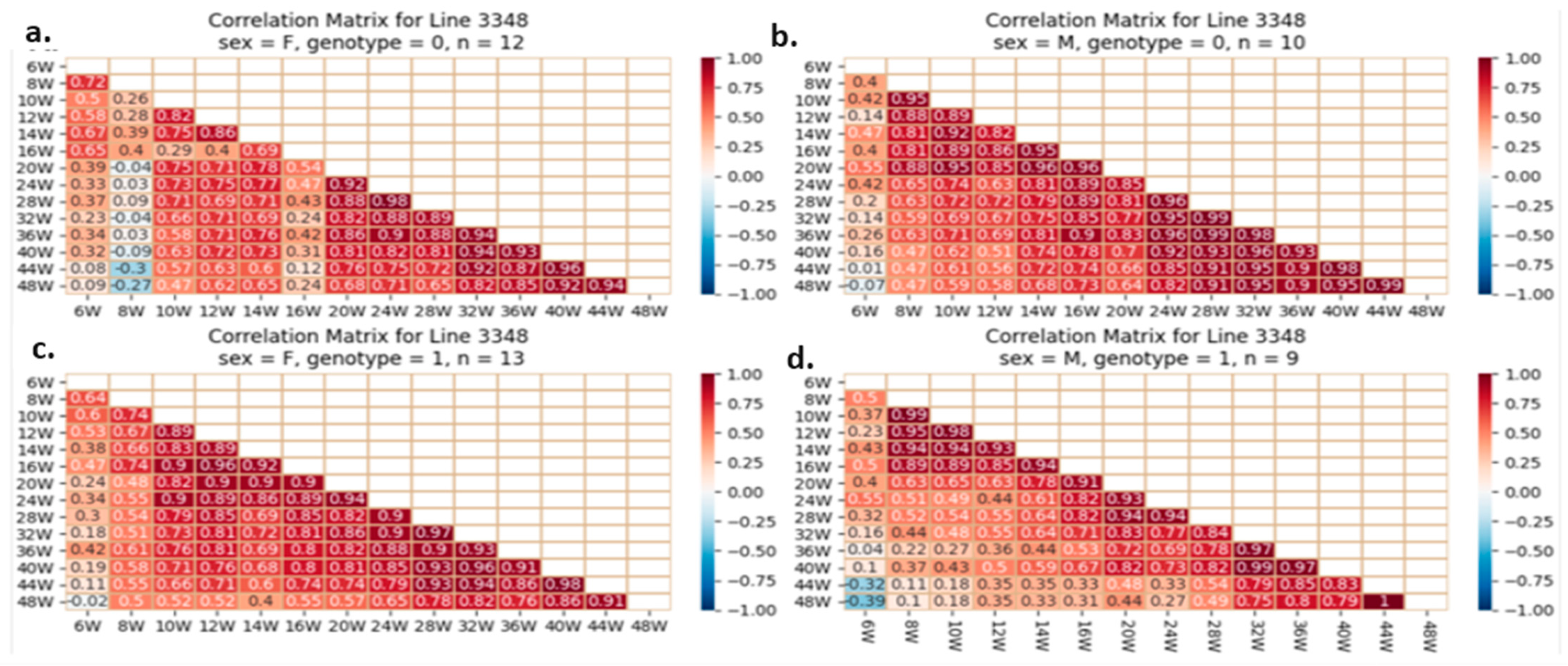
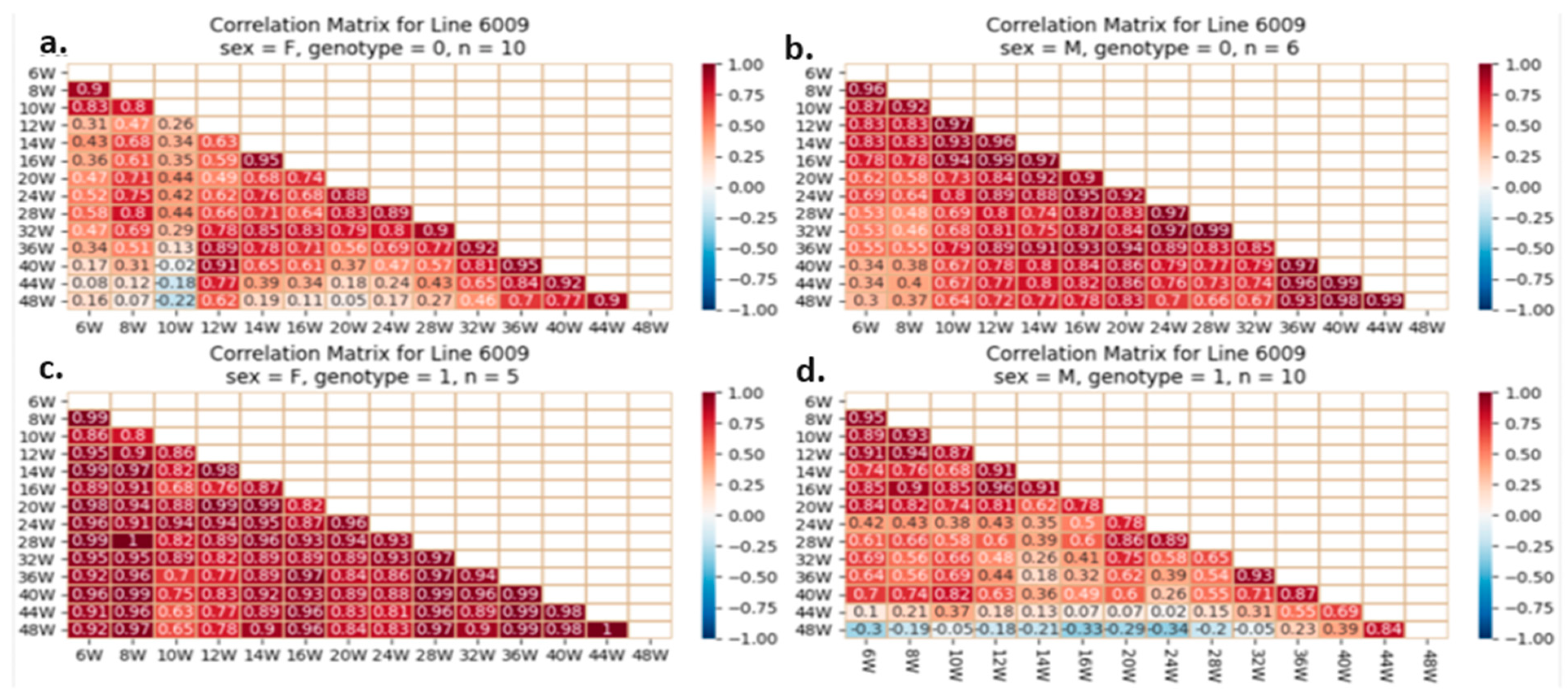
2.5.4. Correlation Analysis between the Studied Traits
3. Discussion
4. Materials and Methods
4.1. Ethical Aspects of the Project
4.2. Study Cohort
4.3. Study Design
4.4. Genotype
4.5. Heritability and Genetic Coefficient Variation
4.6. Computational Methods
4.6.1. Classification Models
4.6.2. Decision Tree
4.6.3. K-Neighbors
4.6.4. Random Forest
4.6.5. Naïve Bayes
4.6.6. Support Vector Machine Classifier
4.6.7. Logistic Regression (LR)
4.6.8. Model Details
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Safaei, M.; Sundararajan, E.A.; Driss, M.; Boulila, W.; Shapi’I, A. A systematic literature review on obesity: Understanding the causes & consequences of obesity and reviewing various machine learning approaches used to predict obesity. Comput. Biol. Med. 2021, 136, 104754. [Google Scholar] [CrossRef]
- El-Sayed Moustafa, J.S.; Froguel, P. From Obesity Genetics to the Future of Personalized Obesity Therapy. Nat. Rev. Endocrinol. 2013, 9, 402–413. [Google Scholar] [CrossRef]
- Lone, I.M.; Midlej, K.; Ben Nun, N.; Iraqi, F.A. Intestinal cancer development in response to oral infection with high-fat diet-induced Type 2 diabetes (T2D) in collaborative cross mice under different host genetic background effects. Mamm. Genome 2023, 34, 56–75. [Google Scholar] [CrossRef]
- Da Luz, F.Q.; Hay, P.; Touyz, S.; Sainsbury, A. Obesity with Comorbid Eating Disorders: Associated Health Risks and Treatment Approaches. Nutrients 2018, 10, 829. [Google Scholar] [CrossRef]
- Ghnaim, A.; Lone, I.M.; Ben Nun, N.; Iraqi, F.A. Unraveling the Host Genetic Background Effect on Internal Organ Weight Influenced by Obesity and Diabetes Using Collaborative Cross Mice. Int. J. Mol. Sci. 2023, 24, 8201. [Google Scholar] [CrossRef]
- Ligibel, J.A.; Alfano, C.M.; Courneya, K.S.; Demark-Wahnefried, W.; Burger, R.A.; Chlebowski, R.T.; Fabian, C.J.; Gucalp, A.; Hershman, D.L.; Hudson, M.M.; et al. American Society of Clinical Oncology Position Statement on Obesity and Cancer. J. Clin. Oncol. 2014, 32, 3568–3574. [Google Scholar] [CrossRef]
- Bhaskaran, K.; Douglas, I.; Forbes, H.; dos-Santos-Silva, I.; Leon, D.A.; Smeeth, L. Body-mass index and risk of 22 specific cancers: A population-based cohort study of 5.24 million UK adults. Lancet 2014, 384, 755–765. [Google Scholar] [CrossRef]
- Yehia, R.; Lone, I.M.; Yehia, I.; Iraqi, F.A. Studying the Pharmagenomic effect of Portulaca oleracea extract on anti-diabetic therapy using the Collaborative Cross mice. Phytomed. Plus 2023, 3, 100394. [Google Scholar] [CrossRef]
- Lone, I.M.; Iraqi, F.A. Genetics of murine type 2 diabetes and comorbidities. Mamm. Genome 2022, 33, 421–436. [Google Scholar] [CrossRef]
- Kamboh, M.I.; Demirci, F.Y.; Wang, X.; Minster, R.L.; Carrasquillo, M.M.; Pankratz, V.S.; Younkin, S.G.; Saykin, A.J.; Jun, G.; Baldwin, C.; et al. Genome-wide association study of Alzheimer’s disease. Transl. Psychiatry 2012, 2, e117. [Google Scholar] [CrossRef]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Kelishadi, R.; Poursafa, P. A Review on the Genetic, Environmental, and Lifestyle Aspects of the Early-Life Origins of Cardiovascular Disease. Curr. Probl. Pediatr. Adolesc. Health Care 2014, 44, 54–72. [Google Scholar] [CrossRef] [PubMed]
- Burza, M.A.; Spagnuolo, R.; Montalcini, T.; Doldo, P.; Pujia, A.; Romeo, S. Effect of Excess Body Weight on the Genetic Susceptibility to Cancer. J. Clin. Gastroenterol. 2014, 48, S78–S79. [Google Scholar] [CrossRef] [PubMed]
- Lone, I.M.; Zohud, O.; Nashef, A.; Kirschneck, C.; Proff, P.; Watted, N.; Iraqi, F.A. Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue. Int. J. Mol. Sci. 2023, 24, 2570. [Google Scholar] [CrossRef]
- Hahn, S.A.; Schutte, M.; Hoque, A.T.M.S.; Moskaluk, C.A.; da Costa, L.T.; Rozenblum, E.; Weinstein, C.L.; Fischer, A.; Yeo, C.J.; Hruban, R.H.; et al. DPC4, A Candidate Tumor Suppressor Gene at Human Chromosome 18q21.1. Science 1996, 271, 350–353. [Google Scholar] [CrossRef]
- Zohud, O.; Lone, I.M.; Midlej, K.; Obaida, A.; Masarwa, S.; Schröder, A.; Küchler, E.C.; Nashef, A.; Kassem, F.; Reiser, V.; et al. Towards Genetic Dissection of Skeletal Class III Malocclusion: A Review of Genetic Variations Underlying the Phenotype in Humans and Future Directions. J. Clin. Med. 2023, 12, 3212. [Google Scholar] [CrossRef]
- Takaku, K.; Oshima, M.; Miyoshi, H.; Matsui, M.; Seldin, M.F.; Taketo, M.M. Intestinal Tumorigenesis in Compound Mutant Mice of both Dpc4(Smad4) and Apc Genes. Cell 1998, 92, 645–656. [Google Scholar] [CrossRef]
- Kitamura, T.; Kometani, K.; Hashida, H.; Matsunaga, A.; Miyoshi, H.; Hosogi, H.; Aoki, M.; Oshima, M.; Hattori, M.; Takabayashi, A.; et al. SMAD4-deficient intestinal tumors recruit CCR1+ myeloid cells that promote invasion. Nat. Genet. 2007, 39, 467–475. [Google Scholar] [CrossRef]
- Izeradjene, K.; Combs, C.; Best, M.; Gopinathan, A.; Wagner, A.; Grady, W.M.; Deng, C.-X.; Hruban, R.H.; Adsay, N.V.; Tuveson, D.A.; et al. KrasG12D and Smad4/Dpc4 Haploinsufficiency Cooperate to Induce Mucinous Cystic Neoplasms and Invasive Adenocarcinoma of the Pancreas. Cancer Cell 2007, 11, 229–243. [Google Scholar] [CrossRef]
- Bardeesy, N.; Cheng, K.-H.; Berger, J.H.; Chu, G.C.; Pahler, J.; Olson, P.; Hezel, A.F.; Horner, J.; Lauwers, G.Y.; Hanahan, D.; et al. Smad4 is dispensable for normal pancreas development yet critical in progression and tumor biology of pancreas cancer. Genes Dev. 2006, 20, 3130–3146. [Google Scholar] [CrossRef]
- Teng, Y.; Sun, A.-N.; Pan, X.-C.; Yang, G.; Yang, L.-L.; Wang, M.-R.; Yang, X. Synergistic Function of Smad4 and PTEN in Suppressing Forestomach Squamous Cell Carcinoma in the Mouse. Cancer Res. 2006, 66, 6972–6981. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Kobayashi, S.; Qiao, W.; Li, C.; Xiao, C.; Radaeva, S.; Stiles, B.; Wang, R.-H.; Ohara, N.; Yoshino, T.; et al. Induction of intrahepatic cholangiocellular carcinoma by liver-specific disruption ofSmad4 andPten in mice. J. Clin. Investig. 2006, 116, 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Pühringer-Oppermann, F.; Sarbia, M.; Ott, N.; Brücher, B.L.D.M. The predictive value of genes of the TGF-β1 pathway in multimodally treated squamous cell carcinoma of the esophagus. Int. J. Color. Dis. 2010, 25, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Richardson, J.A.; Parada, L.F.; Graff, J.M. Smad3 Mutant Mice Develop Metastatic Colorectal Cancer. Cell 1998, 94, 703–714. [Google Scholar] [CrossRef]
- Tan, C.K.; Leuenberger, N.; Tan, M.J.; Yan, Y.W.; Chen, Y.; Kambadur, R.; Wahli, W.; Tan, N.S. Smad3 Deficiency in Mice Protects against Insulin Resistance and Obesity Induced by a High-Fat Diet. Diabetes 2011, 60, 464–476. [Google Scholar] [CrossRef]
- Yadav, H.; Quijano, C.; Kamaraju, A.K.; Gavrilova, O.; Malek, R.; Chen, W.; Zerfas, P.; Zhigang, D.; Wright, E.C.; Stuelten, C.; et al. Protection from Obesity and Diabetes by Blockade of TGF-β/Smad3 Signaling. Cell Metab. 2011, 14, 67–79. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.B.Y.; Chen, W.; Fan, J.; Li, S.; Qu, X.; Chen, Q.; Chen, R.; Zhu, D.; Zhang, J.; et al. Smad4 promotes diabetic nephropathy by modulating glycolysis and OXPHOS. EMBO Rep. 2020, 21, e48781. [Google Scholar] [CrossRef]
- Wang, C.; Wu, Y.; Li, Y.; Zhang, Y.; Fung, Y.L.; Tong, K.K.; Lau, C.W.; Xiang, L.; Kwan, K.M.; You, L.-R.; et al. Smad4-mediated angiogenesis facilitates the beiging of white adipose tissue in mice. iScience 2023, 26, 106272. [Google Scholar] [CrossRef]
- Churchill, G.A.; Airey, D.C.; Allayee, H.; Angel, J.M.; Attie, A.D.; Beatty, J.; Beavis, W.D.; Belknap, J.K.; Bennett, B.; Berrettini, W.; et al. The Collaborative Cross, a Community Resource for the Genetic Analysis of Complex Traits. Nat. Genet. 2004, 36, 1133–1137. [Google Scholar] [CrossRef]
- Aylor, D.L.; Valdar, W.; Foulds-Mathes, W.; Buus, R.J.; Verdugo, R.A.; Baric, R.S.; Ferris, M.T.; Frelinger, J.A.; Heise, M.; Frieman, M.B.; et al. Genetic analysis of complex traits in the emerging Collaborative Cross. Genome Res. 2011, 21, 1213–1222. [Google Scholar] [CrossRef]
- Chesler, E.J.; Miller, D.R.; Branstetter, L.R.; Galloway, L.D.; Jackson, B.L.; Philip, V.M.; Voy, B.H.; Culiat, C.T.; Threadgill, D.W.; Williams, R.W.; et al. The Collaborative Cross at Oak Ridge National Laboratory: Developing a powerful resource for systems genetics. Mamm. Genome 2008, 19, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Atamni, H.A.T.; Nashef, A.; Iraqi, F.A. The Collaborative Cross mouse model for dissecting genetic susceptibility to infectious diseases. Mamm. Genome 2018, 29, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Durrant, C.; Tayem, H.; Yalcin, B.; Cleak, J.; Goodstadt, L.; de Villena, F.P.-M.; Mott, R.; Iraqi, F.A. Collaborative Cross mice and their power to map host susceptibility to Aspergillus fumigatus infection. Genome Res. 2011, 21, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Lone, I.M.; Zohud, O.; Midlej, K.; Awadi, O.; Masarwa, S.; Krohn, S.; Kirschneck, C.; Proff, P.; Watted, N.; Iraqi, F.A. Narrating the Genetic Landscape of Human Class I Occlusion: A Perspective-Infused Review. J. Pers. Med. 2023, 13, 1465. [Google Scholar] [CrossRef] [PubMed]
- Lone, I.M.; Zohud, O.; Midlej, K.; Proff, P.; Watted, N.; Iraqi, F.A. Skeletal Class II Malocclusion: From Clinical Treatment Strategies to the Roadmap in Identifying the Genetic Bases of Development in Humans with the Support of the Collaborative Cross Mouse Population. J. Clin. Med. 2023, 12, 5148. [Google Scholar] [CrossRef] [PubMed]
- Keane, T.M.; Goodstadt, L.; Danecek, P.; White, M.A.; Wong, K.; Yalcin, B.; Heger, A.; Agam, A.; Slater, G.; Goodson, M.; et al. Mouse genomic variation and its effect on phenotypes and gene regulation. Nature 2011, 477, 289–294. [Google Scholar] [CrossRef]
- Edgar, T.W.; Manz, D.O. Research Methods for Cyber Security; Syngress: Oxford, UK, 2017. [Google Scholar]
- Morton, D.; Griffiths, P. Guidelines on the recognition of pain, distress and discomfort in experimental animals and an hypothesis for assessment. Veter. Rec. 1985, 116, 431–436. [Google Scholar] [CrossRef]
- Morton, D.B. A systematic approach for establishing humane endpoints. ILAR J. 2000, 41, 80–86. [Google Scholar] [CrossRef]
- Paster, E.V.; Villines, K.A.; Hickman, D.L. Endpoints for mouse abdominal tumor models: Refinement of current criteria. Comp. Med. 2009, 59, 234–241. [Google Scholar]
- Argilés, J.M.; Busquets, S.; Stemmler, B.; López-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef]
- Snell, G.D. Inheritance in the house mouse, the linkage relations of short-ear, hairless, and naked. Genetics 1931, 16, 42–74. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.R.; Lawler, M.P.; Tordoff, M.G. Reduced body weight is a common effect of gene knockout in mice. BMC Genet. 2008, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Woods, S.C.; Seeley, R.J.; Barsh, G.S.; Baskin, D.G.; Leibel, R.L. Is the Energy Homeostasis System Inherently Biased toward Weight Gain? Diabetes 2003, 52, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Lone, I.M.; Ben Nun, N.; Ghnaim, A.; Schaefer, A.S.; Houri-Haddad, Y.; Iraqi, F.A. High-fat diet and oral infection induced type 2 diabetes and obesity development under different genetic backgrounds. Anim. Model. Exp. Med. 2023, 6, 131–145. [Google Scholar] [CrossRef]
- Nashef, A.; Qahaz, N.; Abu El-Naaj, I.; Iraqi, F.A. Systems genetics analysis of oral squamous cell carcinoma susceptibility using the mouse model: Current position and new perspective. Mamm. Genome 2021, 32, 323–331. [Google Scholar] [CrossRef]
- Truett, G.; Heeger, P.; Mynatt, R.; Truett, A.; Walker, J.; Warman, M.; Meeker, N.D.; Hutchinson, S.A.; Ho, L.; Trede, N.S.; et al. Preparation of PCR-Quality Mouse Genomic DNA with Hot Sodium Hydroxide and Tris (HotSHOT). Biotechniques 2000, 29, 52–54. [Google Scholar] [CrossRef]
- Soofi, A.A.; Awan, A. Classification Techniques in Machine Learning: Applications and Issues. J. Basic Appl. Sci. 2017, 13, 459–465. [Google Scholar] [CrossRef]
- Zou, Q.; Qu, K.; Luo, Y.; Yin, D.; Ju, Y.; Tang, H. Predicting Diabetes Mellitus With Machine Learning Techniques. Front. Genet. 2018, 9, 515. [Google Scholar] [CrossRef]
- van Kruijsdijk, R.C.M.; van der Wall, E.; Visseren, F.L.J. Obesity and Cancer: The Role of Dysfunctional Adipose Tissue. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2569–2578. [Google Scholar] [CrossRef]
- da Silva, M.; Weiderpass, E.; Licaj, I.; Lissner, L.; Rylander, C. Excess body weight, weight gain and obesity-related cancer risk in women in Norway: The Norwegian Women and Cancer study. Br. J. Cancer 2018, 119, 646–656. [Google Scholar] [CrossRef]
- Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; Dandona, L.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived with Disability, and Disability-Adjusted Life-Years for 32 Cancer Groups, 1990 to 2015: A Systematic Analysis for the Global Burden of Disease Study Global Burden of Disease Cancer Collaboration. JAMA Oncol. 2017, 3, 524–548. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
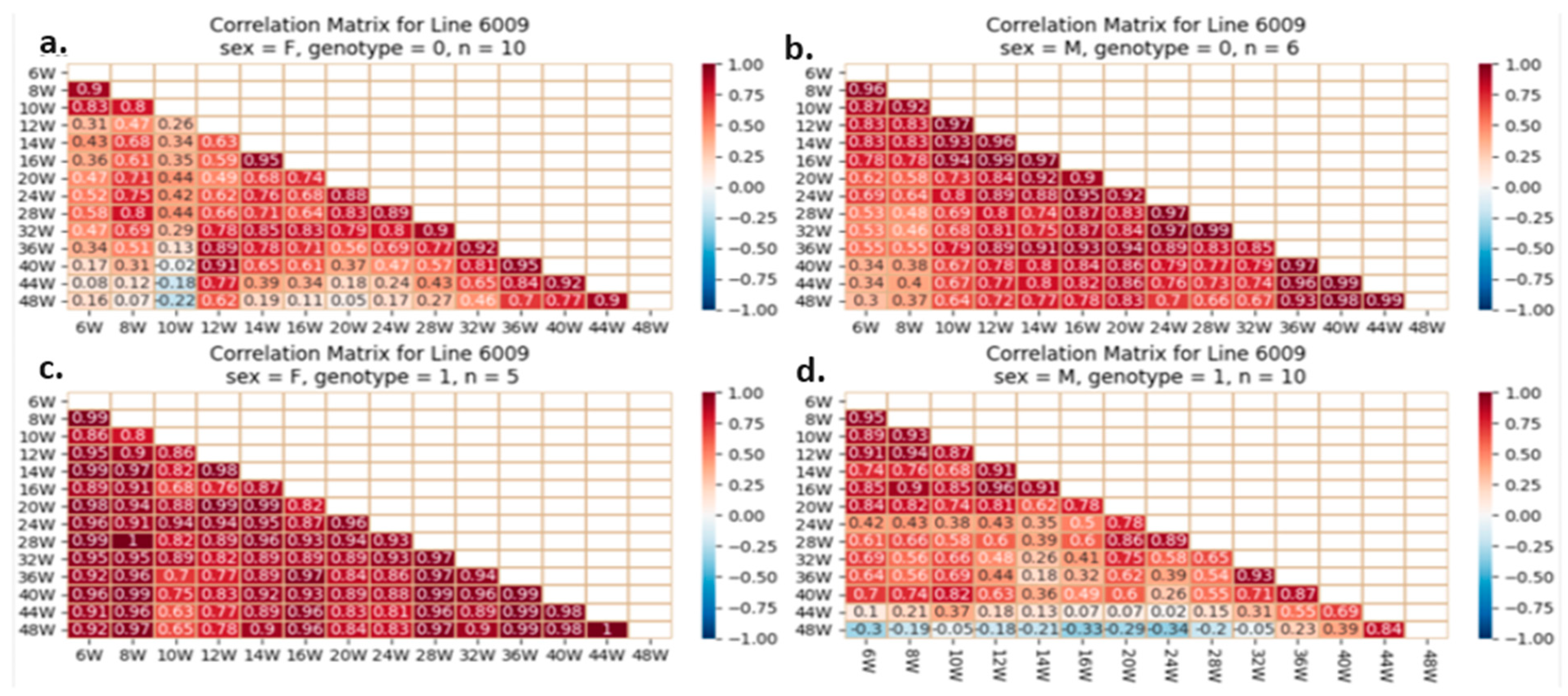{kind=link}
| Male | Female | ||||
|---|---|---|---|---|---|
| Wild Type | Mutant | Wild Type | Mutant | ||
| IL72 | 8 | 11 | 14 | 8 | 41 |
| IL188 | 5 | 8 | 3 | 8 | 24 |
| IL2513 | 8 | 3 | 4 | 12 | 27 |
| IL2750 | 6 | 7 | 6 | 11 | 30 |
| IL3348 | 10 | 9 | 12 | 13 | 44 |
| IL5000 | 14 | 10 | 9 | 12 | 45 |
| IL5008 | 9 | 12 | 13 | 11 | 45 |
| IL6009 | 6 | 10 | 10 | 5 | 31 |
| IL6012 | 10 | 9 | 6 | 11 | 36 |
| IL6018 | 16 | 18 | 19 | 15 | 68 |
| Total | 391 | ||||
| Frequency | Percent (%) | Total | |
|---|---|---|---|
| Wild Type | 188 | 48.1 | |
| Mutant | 203 | 51.9 | |
| Female | 202 | 51.7 | |
| Male | 189 | 48.3 | |
| 391 (100%) |
| Trait | df between | df within | n | MS between | MS within | VG | H2 | ||
|---|---|---|---|---|---|---|---|---|---|
| Female | Wild | %∆BW 16-6 | 10 | 87 | 8 | 582.107 | 170.343 | 51.47049 | 0.232044 |
| %∆BW 32-6 | 10 | 87 | 8 | 5380.415 | 464.75 | 614.4581 | 0.56936 | ||
| %∆BW 40-6 | 10 | 87 | 8 | 7035.673 | 457.407 | 822.2833 | 0.642564 | ||
| %∆BW 48-6 | 10 | 86 | 7.909 | 6986.655 | 612.58 | 792.0095 | 0.659886 | ||
| Mutant | %∆BW 16-6 | 10 | 97 | 8.909 | 682.415 | 154.546 | 59.2506 | 0.277135 | |
| %∆BW 32-6 | 10 | 97 | 8.909 | 7073.207 | 497.555 | 738.0834 | 0.59733 | ||
| %∆BW 40-6 | 10 | 97 | 8.909 | 7974.515 | 570.821 | 831.0269 | 0.592808 | ||
| %∆BW 48-6 | 10 | 97 | 8.909 | 8705.475 | 614.002 | 908.2266 | 0.596643 | ||
| Male | Wild | %∆BW 16-6 | 10 | 83 | 7.636 | 923.507 | 289.006 | 83.08942 | 0.223301 |
| %∆BW 32-6 | 10 | 83 | 7.636 | 3753.41 | 400.828 | 439.0286 | 0.522742 | ||
| %∆BW 40-6 | 10 | 83 | 7.636 | 3558.534 | 476.411 | 403.6113 | 0.458638 | ||
| %∆BW 48-6 | 10 | 83 | 7.636 | 4110.591 | 541.017 | 467.4442 | 0.463522 | ||
| Mutant | %∆BW 16-6 | 10 | 88 | 8.091 | 1230.387 | 201.046 | 127.2219 | 0.387555 | |
| %∆BW 32-6 | 10 | 88 | 8.091 | 3524.053 | 361.562 | 390.8697 | 0.519475 | ||
| %∆BW 40-6 | 10 | 88 | 8.091 | 2554.555 | 403.156 | 265.9032 | 0.397429 | ||
| %∆BW 48-6 | 10 | 88 | 8.091 | 2597.145 | 536.887 | 254.6386 | 0.321706 |
| Line | IL72 | IL188 | IL2513 | IL2750 | IL3348 | IL5000 | IL5008 | IL6009 | IL6012 | IL6018 |
|---|---|---|---|---|---|---|---|---|---|---|
| N | 41 | 24 | 27 | 29 | 44 | 45 | 45 | 31 | 36 | 68 |
| DT | 0.504 | 0.5 | 0.652 | 0.513 | 0.41 | 0.497 | 0.543 | 0.58 | 0.494 | 0.433 |
| NaBa | 0.603 | 0.459 | 0.67 | 0.58 | 0.553 | 0.491 | 0.584 | 0.798 | 0.528 | 0.644 |
| KNN | 0.506 | 0.5 | 0.718 | 0.637 | 0.555 | 0.42 | 0.556 | 0.787 | 0.483 | 0.539 |
| RF | 0.543 | 0.619 | 0.747 | 0.546 | 0.571 | 0.512 | 0.457 | 0.718 | 0.518 | 0.561 |
| SVC | 0.418 | 0.438 | 0.682 | 0.386 | 0.407 | 0.472 | 0.325 | 0.836 | 0.459 | 0.576 |
| LR | 0.332 | 0.747 | 0.875 | 0.752 | 0.531 | 0.392 | 0.557 | 0.82 | 0.405 | 0.586 |
| Line | IL72 | IL188 | IL2513 | IL2750 | IL3348 | IL5000 | IL5008 | IL6009 | IL6012 | IL6018 |
|---|---|---|---|---|---|---|---|---|---|---|
| N | 41 | 24 | 27 | 29 | 44 | 45 | 45 | 31 | 36 | 68 |
| 6W | 0.29 | −0.30 | 0.64 | 0.08 | 0.30 | 0.35 | 0.38 | 0.23 | 0.33 | 0.44 |
| 8W | 0.24 | −0.21 | 0.63 | 0.13 | 0.37 | 0.37 | 0.56 | 0.15 | 0.27 | 0.53 |
| 10W | 0.26 | −0.03 | 0.60 | 0.28 | 0.57 | 0.39 | 0.53 | 0.08 | 0.37 | 0.51 |
| 12W | 0.35 | −0.20 | 0.64 | 0.44 | 0.52 | 0.32 | 0.52 | 0.13 | 0.27 | 0.48 |
| 14W | 0.43 | −0.32 | 0.59 | 0.41 | 0.54 | 0.25 | 0.54 | 0.03 | 0.32 | 0.61 |
| 16W | 0.32 | −0.40 | 0.56 | 0.33 | 0.53 | 0.21 | 0.58 | −0.15 | 0.31 | 0.64 |
| 20W | 0.47 | −1.23 | 0.42 | 0.13 | 0.51 | 0.20 | 0.71 | −0.12 | 0.25 | 0.61 |
| 24W | 0.46 | −1.58 | 0.55 | 0.15 | 0.66 | 0.06 | 0.74 | −0.28 | 0.40 | 0.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qahaz, N.; Lone, I.M.; Khadija, A.; Ghnaim, A.; Zohud, O.; Nun, N.B.; Nashef, A.; Abu El-Naaj, I.; Iraqi, F.A. Host Genetic Background Effect on Body Weight Changes Influenced by Heterozygous Smad4 Knockout Using Collaborative Cross Mouse Population. Int. J. Mol. Sci. 2023, 24, 16136. https://doi.org/10.3390/ijms242216136
Qahaz N, Lone IM, Khadija A, Ghnaim A, Zohud O, Nun NB, Nashef A, Abu El-Naaj I, Iraqi FA. Host Genetic Background Effect on Body Weight Changes Influenced by Heterozygous Smad4 Knockout Using Collaborative Cross Mouse Population. International Journal of Molecular Sciences. 2023; 24(22):16136. https://doi.org/10.3390/ijms242216136
Chicago/Turabian StyleQahaz, Nayrouz, Iqbal M. Lone, Aya Khadija, Aya Ghnaim, Osayd Zohud, Nadav Ben Nun, Aysar Nashef, Imad Abu El-Naaj, and Fuad A. Iraqi. 2023. "Host Genetic Background Effect on Body Weight Changes Influenced by Heterozygous Smad4 Knockout Using Collaborative Cross Mouse Population" International Journal of Molecular Sciences 24, no. 22: 16136. https://doi.org/10.3390/ijms242216136
APA StyleQahaz, N., Lone, I. M., Khadija, A., Ghnaim, A., Zohud, O., Nun, N. B., Nashef, A., Abu El-Naaj, I., & Iraqi, F. A. (2023). Host Genetic Background Effect on Body Weight Changes Influenced by Heterozygous Smad4 Knockout Using Collaborative Cross Mouse Population. International Journal of Molecular Sciences, 24(22), 16136. https://doi.org/10.3390/ijms242216136