Extracellular Vesicles and Their Role in Lung Infections



Abstract
:1. Introduction
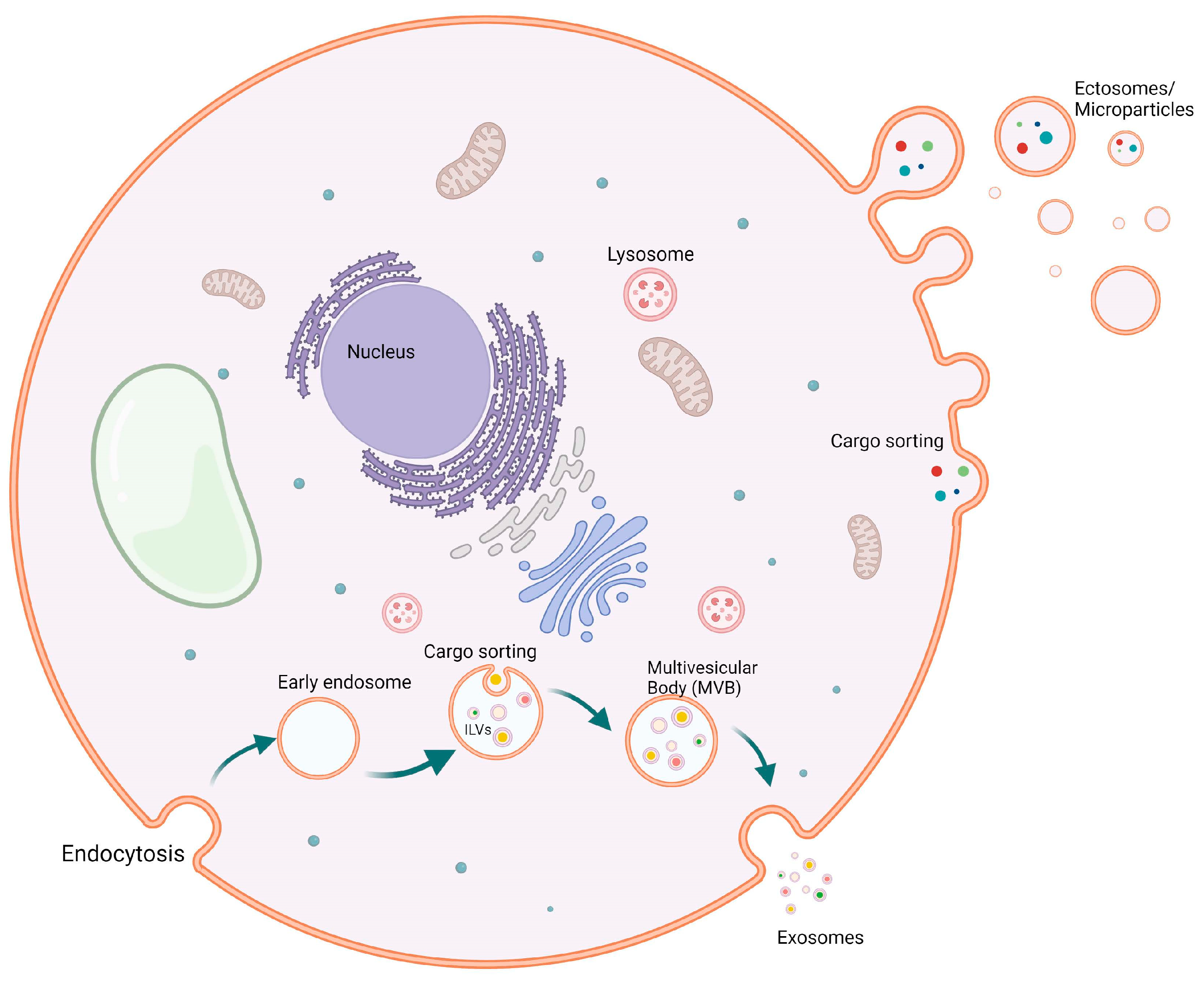
2. Function of Extracellular Vesicles
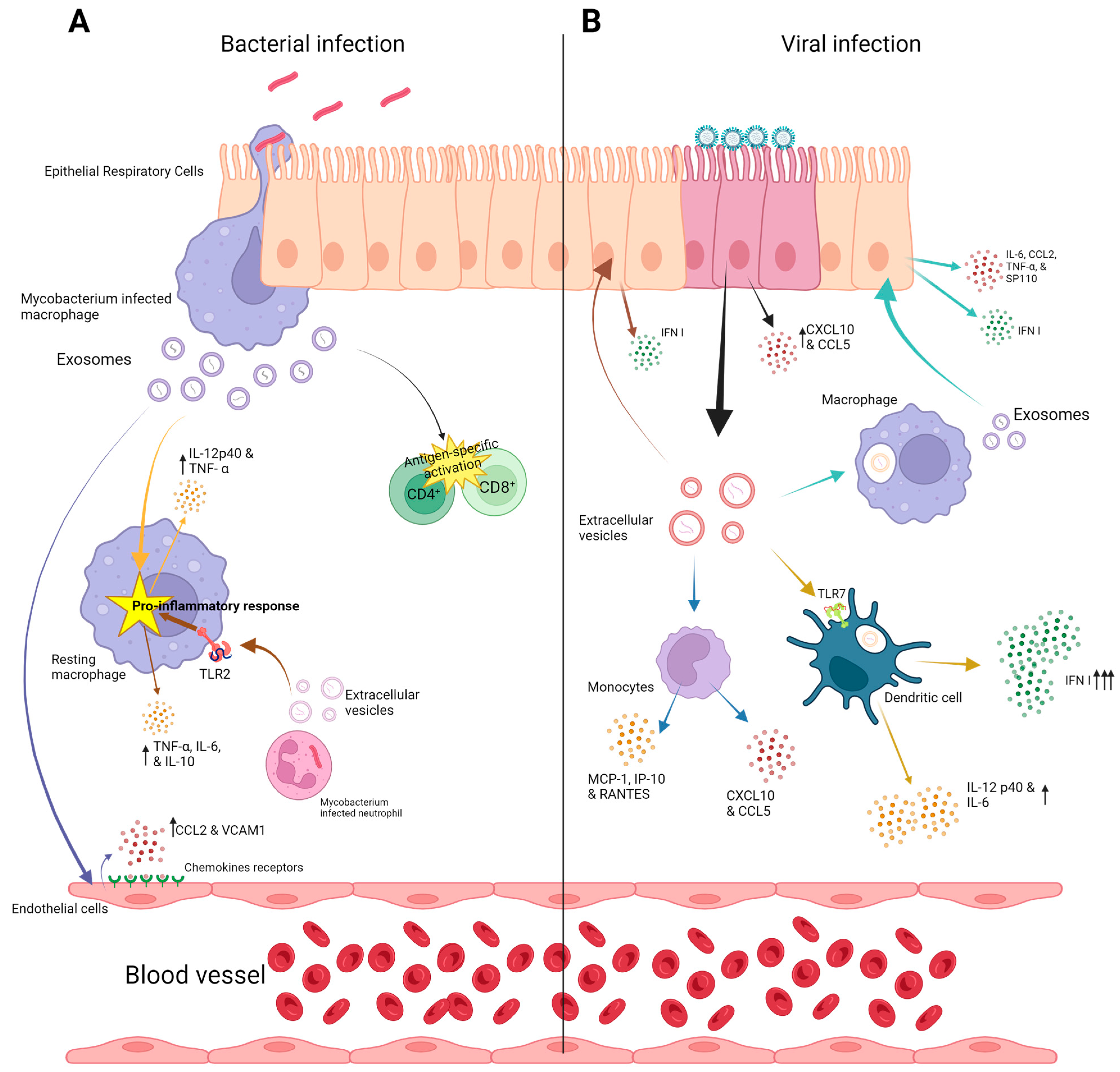
2.1. Extracellular Vesicles in Bacterial Lung Infections
2.2. Extracellular Vesicles in Viral Lung Infections
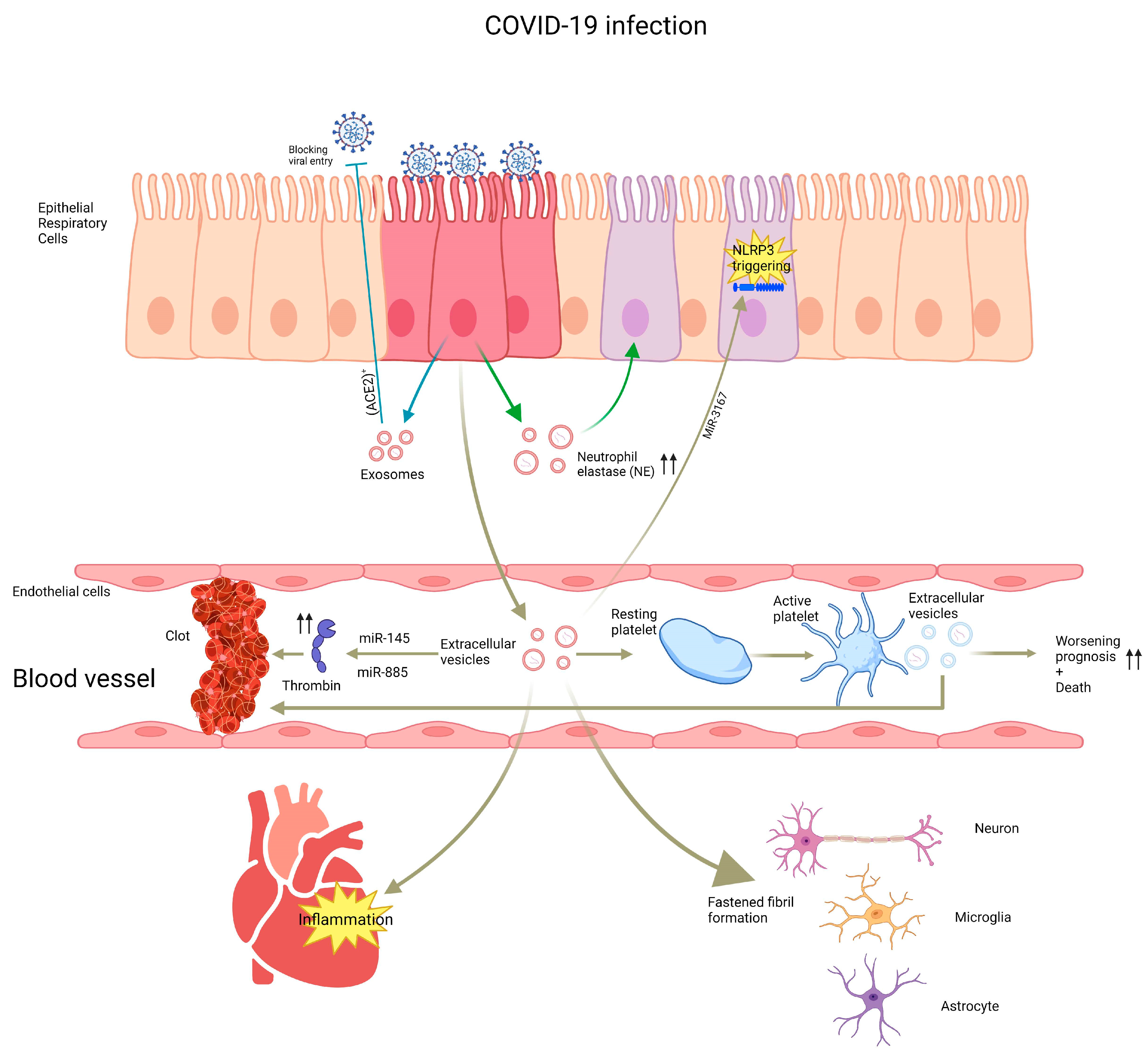
2.3. Extracellular Vesicles as a Biomarker in Lung infections
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACE2 | Angiotensin-converting enzyme 2 |
| AKR1B10 | Aldo-keto reductase 1 B10 |
| ARDS | Acute respiratory distress syndrome |
| ARF6 | ADP-ribosylation factor 6 |
| BALF | Bronchoalveolar lavage fluid |
| BCG | Mycobacterium bovis |
| CCL2 | Chemokine (C-C motif) ligand 2 |
| COPB2 | coat complex subunit beta 2 |
| ESCRT | Endosomal complexes required for transport |
| EVs | Extracellular vesicles |
| Hsp | Heat shock protein |
| IAV | Influenza A virus |
| IFN | Interferon |
| ILVs | Intraluminal vesicles |
| IP-10 | interferon-gamma-induced protein 10 |
| KRAS | KRAS proto-oncogene |
| LAM | Lipoarabinomannan |
| LC3 | Microtubule-associated protein 1A/1B-light chain 3 |
| MAP | Mycobacterium avium subsp. Paratuberculosis |
| MCP-1 | Monocyte Chemoattractant Protein-1MVE |
| MOIs | Multiplicity of infections |
| Mtb | Mycobacterium tuberculosis |
| MV | Membrane vesicle |
| MyD88 | Myeloid differentiation primary response 88 |
| NE | Neutrophil elastase |
| OMV | Outer membrane vesicles |
| PAMPs | Pathogen-associated molecular patterns |
| PBECs | Primary human bronchial epithelial cells |
| PBMCs | Human peripheral blood mononuclear cells |
| PRKCB | Protein kinase C beta |
| Rabs | Ras-associated binding proteins |
| RANTES | regulated on activation, normal T cell expressed and secreted |
| RHOC | Ras homolog family member C |
| RIG-I | Retinoic-acid-inducible gene I |
| RSV | Respiratory syncytial virus |
| S-EV | Spontaneously released EVs |
| TIM-4 | T-cell immunoglobulin mucin protein 4 |
| TLR2 | Toll-like receptor 2 |
| TN-C | Tenascin-C |
| TNF | Tumor necrosis factor |
| TOM20 | Mitochondrial import receptor subunit 20 |
References
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Agarwal, S.; Clauss, M.; Britt, N.S.; Dhillon, N.K. Extracellular vesicles: Novel communicators in lung diseases. Respir. Res. 2020, 21, 175. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Teng, K.; Wu, C.; Adam, M.; Johnstone, R.M. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol. 1985, 101, 942–948. [Google Scholar] [CrossRef]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Colombo, M.; Moita, C.; van Niel, G.; Kowal, J.; Vigneron, J.; Benaroch, P.; Manel, N.; Moita, L.F.; Thery, C.; Raposo, G. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 2013, 126, 5553–5565. [Google Scholar] [CrossRef]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef]
- Muralidharan-Chari, V.; Clancy, J.; Plou, C.; Romao, M.; Chavrier, P.; Raposo, G.; D’Souza-Schorey, C. ARF6-regulated shedding of tumor cell-derived plasma membrane microvesicles. Curr. Biol. 2009, 19, 1875–1885. [Google Scholar] [CrossRef]
- Thery, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Bose, S.; Aggarwal, S.; Singh, D.V.; Acharya, N. Extracellular vesicles: An emerging platform in gram-positive bacteria. Microb. Cell 2020, 7, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small GTPases 2017, 8, 220–232. [Google Scholar] [CrossRef]
- Hugel, B.; Martinez, M.C.; Kunzelmann, C.; Freyssinet, J.M. Membrane microparticles: Two sides of the coin. Physiology 2005, 20, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.; Pendlebury, S.A.; Green, C. Lipid organization in erythrocyte membrane microvesicles. Biochem. J. 1984, 224, 285–290. [Google Scholar] [CrossRef]
- Kastelowitz, N.; Yin, H. Exosomes and microvesicles: Identification and targeting by particle size and lipid chemical probes. Chembiochem 2014, 15, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Ignatchenko, V.; Ignatchenko, A.; Mejia-Guerrero, S.; Kislinger, T. In-depth proteomic analyses of ovarian cancer cell line exosomes reveals differential enrichment of functional categories compared to the NCI 60 proteome. Biochem. Biophys. Res. Commun. 2014, 445, 694–701. [Google Scholar] [CrossRef]
- Christianson, H.C.; Svensson, K.J.; van Kuppevelt, T.H.; Li, J.P.; Belting, M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef]
- Todkar, K.; Chikhi, L.; Desjardins, V.; El-Mortada, F.; Pepin, G.; Germain, M. Selective packaging of mitochondrial proteins into extracellular vesicles prevents the release of mitochondrial DAMPs. Nat. Commun. 2021, 12, 1971. [Google Scholar] [CrossRef]
- van Niel, G.; Carter, D.R.F.; Clayton, A.; Lambert, D.W.; Raposo, G.; Vader, P. Challenges and directions in studying cell-cell communication by extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2022, 23, 369–382. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Bobrie, A.; Colombo, M.; Raposo, G.; Thery, C. Exosome secretion: Molecular mechanisms and roles in immune responses. Traffic 2011, 12, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta 2012, 1820, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Mears, R.; Craven, R.A.; Hanrahan, S.; Totty, N.; Upton, C.; Young, S.L.; Patel, P.; Selby, P.J.; Banks, R.E. Proteomic analysis of melanoma-derived exosomes by two-dimensional polyacrylamide gel electrophoresis and mass spectrometry. Proteomics 2004, 4, 4019–4031. [Google Scholar] [CrossRef]
- Schorey, J.S.; Bhatnagar, S. Exosome function: From tumor immunology to pathogen biology. Traffic 2008, 9, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Rani, S.; O’Brien, K.; Kelleher, F.C.; Corcoran, C.; Germano, S.; Radomski, M.W.; Crown, J.; O’Driscoll, L. Isolation of exosomes for subsequent mRNA, MicroRNA, and protein profiling. Methods Mol. Biol. 2011, 784, 181–195. [Google Scholar] [CrossRef]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef]
- Qu, X.; Li, Q.; Yang, J.; Zhao, H.; Wang, F.; Zhang, F.; Zhang, S.; Zhang, H.; Wang, R.; Wang, Q.; et al. Double-Stranded DNA in Exosomes of Malignant Pleural Effusions as a Novel DNA Source for EGFR Mutation Detection in Lung Adenocarcinoma. Front. Oncol. 2019, 9, 931. [Google Scholar] [CrossRef]
- Fernando, M.R.; Jiang, C.; Krzyzanowski, G.D.; Ryan, W.L. New evidence that a large proportion of human blood plasma cell-free DNA is localized in exosomes. PLoS ONE 2017, 12, e0183915. [Google Scholar] [CrossRef]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef]
- Vagner, T.; Spinelli, C.; Minciacchi, V.R.; Balaj, L.; Zandian, M.; Conley, A.; Zijlstra, A.; Freeman, M.R.; Demichelis, F.; De, S.; et al. Large extracellular vesicles carry most of the tumour DNA circulating in prostate cancer patient plasma. J. Extracell. Vesicles 2018, 7, 1505403. [Google Scholar] [CrossRef]
- Cai, J.; Han, Y.; Ren, H.; Chen, C.; He, D.; Zhou, L.; Eisner, G.M.; Asico, L.D.; Jose, P.A.; Zeng, C. Extracellular vesicle-mediated transfer of donor genomic DNA to recipient cells is a novel mechanism for genetic influence between cells. J. Mol. Cell Biol. 2013, 5, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Torralba, D.; Baixauli, F.; Villarroya-Beltri, C.; Fernandez-Delgado, I.; Latorre-Pellicer, A.; Acin-Perez, R.; Martin-Cofreces, N.B.; Jaso-Tamame, A.L.; Iborra, S.; Jorge, I.; et al. Priming of dendritic cells by DNA-containing extracellular vesicles from activated T cells through antigen-driven contacts. Nat. Commun. 2018, 9, 2658. [Google Scholar] [CrossRef]
- Santavanond, J.P.; Rutter, S.F.; Atkin-Smith, G.K.; Poon, I.K.H. Apoptotic Bodies: Mechanism of Formation, Isolation and Functional Relevance. Subcell. Biochem. 2021, 97, 61–88. [Google Scholar] [CrossRef]
- Ma, L.; Li, Y.; Peng, J.; Wu, D.; Zhao, X.; Cui, Y.; Chen, L.; Yan, X.; Du, Y.; Yu, L. Discovery of the migrasome, an organelle mediating release of cytoplasmic contents during cell migration. Cell Res. 2015, 25, 24–38. [Google Scholar] [CrossRef]
- Nicolas-Avila, J.A.; Lechuga-Vieco, A.V.; Esteban-Martinez, L.; Sanchez-Diaz, M.; Diaz-Garcia, E.; Santiago, D.J.; Rubio-Ponce, A.; Li, J.L.; Balachander, A.; Quintana, J.A.; et al. A Network of Macrophages Supports Mitochondrial Homeostasis in the Heart. Cell 2020, 183, 94–109 e123. [Google Scholar] [CrossRef]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef]
- Wang, G.; Li, J.; Bojmar, L.; Chen, H.; Li, Z.; Tobias, G.C.; Hu, M.; Homan, E.A.; Lucotti, S.; Zhao, F.; et al. Tumour extracellular vesicles and particles induce liver metabolic dysfunction. Nature 2023, 618, 374–382. [Google Scholar] [CrossRef]
- Benet, S.; Galvez, C.; Drobniewski, F.; Kontsevaya, I.; Arias, L.; Monguio-Tortajada, M.; Erkizia, I.; Urrea, V.; Ong, R.Y.; Luquin, M.; et al. Dissemination of Mycobacterium tuberculosis is associated to a SIGLEC1 null variant that limits antigen exchange via trafficking extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12046. [Google Scholar] [CrossRef]
- Li, L.; Cheng, Y.; Emrich, S.; Schorey, J. Activation of endothelial cells by extracellular vesicles derived from Mycobacterium tuberculosis infected macrophages or mice. PLoS ONE 2018, 13, e0198337. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Shinagawa, K.; Castellino, F.J.; Schorey, J.S. Exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and in vivo. Blood 2007, 110, 3234–3244. [Google Scholar] [CrossRef]
- Javadi, A.; Shamaei, M.; Tabarsi, P.; Nomani, M.; Varahram, M.; Kazemi, B. Extracellular vesicles from serum samples of mycobacteria patients induced cell death of THP-1 monocyte and PBMC. BMC Pulm. Med. 2022, 22, 57. [Google Scholar] [CrossRef]
- Alvarez-Jimenez, V.D.; Leyva-Paredes, K.; Garcia-Martinez, M.; Vazquez-Flores, L.; Garcia-Paredes, V.G.; Campillo-Navarro, M.; Romo-Cruz, I.; Rosales-Garcia, V.H.; Castaneda-Casimiro, J.; Gonzalez-Pozos, S.; et al. Extracellular Vesicles Released from Mycobacterium tuberculosis-Infected Neutrophils Promote Macrophage Autophagy and Decrease Intracellular Mycobacterial Survival. Front. Immunol. 2018, 9, 272. [Google Scholar] [CrossRef]
- Cheng, Y.; Schorey, J.S. Extracellular vesicles deliver Mycobacterium RNA to promote host immunity and bacterial killing. EMBO Rep. 2019, 20, e46613. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, M.; Vazquez-Flores, L.; Alvarez-Jimenez, V.D.; Castaneda-Casimiro, J.; Ibanez-Hernandez, M.; Sanchez-Torres, L.E.; Barrios-Payan, J.; Mata-Espinosa, D.; Estrada-Parra, S.; Chacon-Salinas, R.; et al. Extracellular vesicles released by J774A.1 macrophages reduce the bacterial load in macrophages and in an experimental mouse model of tuberculosis. Int. J. Nanomed. 2019, 14, 6707–6719. [Google Scholar] [CrossRef]
- Ramachandra, L.; Qu, Y.; Wang, Y.; Lewis, C.J.; Cobb, B.A.; Takatsu, K.; Boom, W.H.; Dubyak, G.R.; Harding, C.V. Mycobacterium tuberculosis synergizes with ATP to induce release of microvesicles and exosomes containing major histocompatibility complex class II molecules capable of antigen presentation. Infect. Immun. 2010, 78, 5116–5125. [Google Scholar] [CrossRef] [PubMed]
- Giri, P.K.; Schorey, J.S. Exosomes derived from M. Bovis BCG infected macrophages activate antigen-specific CD4+ and CD8+ T cells in vitro and in vivo. PLoS ONE 2008, 3, e2461. [Google Scholar] [CrossRef]
- Singh, P.P.; LeMaire, C.; Tan, J.C.; Zeng, E.; Schorey, J.S. Exosomes released from M. tuberculosis infected cells can suppress IFN-gamma mediated activation of naive macrophages. PLoS ONE 2011, 6, e18564. [Google Scholar] [CrossRef] [PubMed]
- Athman, J.J.; Sande, O.J.; Groft, S.G.; Reba, S.M.; Nagy, N.; Wearsch, P.A.; Richardson, E.T.; Rojas, R.; Boom, W.H.; Shukla, S.; et al. Mycobacterium tuberculosis Membrane Vesicles Inhibit T Cell Activation. J. Immunol. 2017, 198, 2028–2037. [Google Scholar] [CrossRef]
- Letsiou, E.; Teixeira Alves, L.G.; Felten, M.; Mitchell, T.J.; Muller-Redetzky, H.C.; Dudek, S.M.; Witzenrath, M. Neutrophil-Derived Extracellular Vesicles Activate Platelets after Pneumolysin Exposure. Cells 2021, 10, 3581. [Google Scholar] [CrossRef]
- Letsiou, E.; Teixeira Alves, L.G.; Fatykhova, D.; Felten, M.; Mitchell, T.J.; Muller-Redetzky, H.C.; Hocke, A.C.; Witzenrath, M. Microvesicles released from pneumolysin-stimulated lung epithelial cells carry mitochondrial cargo and suppress neutrophil oxidative burst. Sci. Rep. 2021, 11, 9529. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Bertazzo, S.; O’Callaghan, D.J.; Schlegel, A.A.; Kallepitis, C.; Antcliffe, D.B.; Gordon, A.C.; Stevens, M.M. Differentiating sepsis from non-infectious systemic inflammation based on microvesicle-bacteria aggregation. Nanoscale 2015, 7, 13511–13520. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.R.; Lempke, S.L.; Miller, M.M.; Bush, D.M.; Braswell, B.G.; Estes, C.L.; Benedict, E.L.; Mahon, A.R.; Sabo, S.L.; Greenlee-Wacker, M.C. Effect of extracellular vesicles from S. aureus-challenged human neutrophils on macrophages. J. Leukoc. Biol. 2020, 108, 1841–1850. [Google Scholar] [CrossRef]
- Koeppen, K.; Nymon, A.; Barnaby, R.; Bashor, L.; Li, Z.; Hampton, T.H.; Liefeld, A.E.; Kolling, F.W.; LaCroix, I.S.; Gerber, S.A.; et al. Let-7b-5p in vesicles secreted by human airway cells reduces biofilm formation and increases antibiotic sensitivity of P. aeruginosa. Proc. Natl. Acad. Sci. USA 2021, 118, e2105370118. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.L.; Herkt, C.E.; Schulz, C.; Bolte, K.; Seidel, K.; Scheller, N.; Sittka-Stark, A.; Bertrams, W.; Schmeck, B. Legionella pneumophila infection activates bystander cells differentially by bacterial and host cell vesicles. Sci. Rep. 2017, 7, 6301. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Belser, J.A. Resurrected pandemic influenza viruses. Annu. Rev. Microbiol. 2009, 63, 79–98. [Google Scholar] [CrossRef]
- Scheller, N.; Herold, S.; Kellner, R.; Bertrams, W.; Jung, A.L.; Janga, H.; Greulich, T.; Schulte, L.N.; Vogelmeier, C.F.; Lohmeyer, J.; et al. Proviral MicroRNAs Detected in Extracellular Vesicles from Bronchoalveolar Lavage Fluid of Patients With Influenza Virus-Induced Acute Respiratory Distress Syndrome. J. Infect. Dis. 2019, 219, 540–543. [Google Scholar] [CrossRef]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef]
- Liu, Y.M.; Tseng, C.H.; Chen, Y.C.; Yu, W.Y.; Ho, M.Y.; Ho, C.Y.; Lai, M.M.C.; Su, W.C. Exosome-delivered and Y RNA-derived small RNA suppresses influenza virus replication. J. Biomed. Sci. 2019, 26, 58. [Google Scholar] [CrossRef]
- Katze, M.G.; He, Y.; Gale, M., Jr. Viruses and interferon: A fight for supremacy. Nat. Rev. Immunol. 2002, 2, 675–687. [Google Scholar] [CrossRef]
- Ge, Y.; Liu, K.; Chi, Y.; Zhu, X.; Wu, T.; Zhao, K.; Qiao, Q.; Wu, B.; Zhu, F.; Cui, L. Exosomal microRNA expression profiles derived from A549 human lung cells in response to influenza A/H1N1pdm09 infection. Virology 2022, 574, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kesimer, M.; Scull, M.; Brighton, B.; DeMaria, G.; Burns, K.; O’Neal, W.; Pickles, R.J.; Sheehan, J.K. Characterization of exosome-like vesicles released from human tracheobronchial ciliated epithelium: A possible role in innate defense. FASEB J. 2009, 23, 1858–1868. [Google Scholar] [CrossRef] [PubMed]
- Maemura, T.; Fukuyama, S.; Sugita, Y.; Lopes, T.J.S.; Nakao, T.; Noda, T.; Kawaoka, Y. Lung-Derived Exosomal miR-483-3p Regulates the Innate Immune Response to Influenza Virus Infection. J. Infect. Dis. 2018, 217, 1372–1382. [Google Scholar] [CrossRef]
- Maemura, T.; Fukuyama, S.; Kawaoka, Y. High Levels of miR-483-3p Are Present in Serum Exosomes Upon Infection of Mice With Highly Pathogenic Avian Influenza Virus. Front. Microbiol. 2020, 11, 144. [Google Scholar] [CrossRef]
- Xia, C.; Xu, W.; Ai, X.; Zhu, Y.; Geng, P.; Niu, Y.; Zhu, H.; Zhou, W.; Huang, H.; Shi, X. Autophagy and Exosome Coordinately Enhance Macrophage M1 Polarization and Recruitment in Influenza A Virus Infection. Front. Immunol. 2022, 13, 722053. [Google Scholar] [CrossRef]
- Schneider, D.J.; Smith, K.A.; Latuszek, C.E.; Wilke, C.A.; Lyons, D.M.; Penke, L.R.; Speth, J.M.; Marthi, M.; Swanson, J.A.; Moore, B.B.; et al. Alveolar macrophage-derived extracellular vesicles inhibit endosomal fusion of influenza virus. EMBO J. 2020, 39, e105057. [Google Scholar] [CrossRef]
- Ludlow, M. Respiratory syncytial virus infection in the modern era. Curr. Opin. Infect. Dis. 2023, 36, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Li, X.; Dacosta-Urbieta, A.; Billard, M.N.; Wildenbeest, J.; Korsten, K.; Martinon-Torres, F.; Heikkinen, T.; Cunningham, S.; Snape, M.D.; et al. Economic burden and health-related quality-of-life among infants with respiratory syncytial virus infection: A multi-country prospective cohort study in Europe. Vaccine 2023, 41, 2707–2715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jamaluddin, M.; Zhang, Y.; Sun, H.; Ivanciuc, T.; Garofalo, R.P.; Brasier, A.R. Systematic Analysis of Cell-Type Differences in the Epithelial Secretome Reveals Insights into the Pathogenesis of Respiratory Syncytial Virus-Induced Lower Respiratory Tract Infections. J. Immunol. 2017, 198, 3345–3364. [Google Scholar] [CrossRef]
- Liu, M.; Guo, S.; Stiles, J.K. The emerging role of CXCL10 in cancer (Review). Oncol. Lett. 2011, 2, 583–589. [Google Scholar] [CrossRef]
- Culley, F.J.; Pennycook, A.M.; Tregoning, J.S.; Dodd, J.S.; Walzl, G.; Wells, T.N.; Hussell, T.; Openshaw, P.J. Role of CCL5 (RANTES) in viral lung disease. J. Virol. 2006, 80, 8151–8157. [Google Scholar] [CrossRef]
- Chahar, H.S.; Corsello, T.; Kudlicki, A.S.; Komaravelli, N.; Casola, A. Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells. Sci. Rep. 2018, 8, 387. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sinha, A.; Lutter, R.; Yang, J.; Ascoli, C.; Sterk, P.J.; Nemsick, N.K.; Perkins, D.L.; Finn, P.W. Analysis of Exosomal MicroRNA Dynamics in Response to Rhinovirus Challenge in a Longitudinal Case-Control Study of Asthma. Viruses 2022, 14, 2444. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.T.; Schwenzer, A.; Marsh, E.K.; Edwards, M.R.; Sabroe, I.; Midwood, K.S.; Parker, L.C. Airway Epithelial Cells Generate Pro-inflammatory Tenascin-C and Small Extracellular Vesicles in Response to TLR3 Stimuli and Rhinovirus Infection. Front. Immunol. 2019, 10, 1987. [Google Scholar] [CrossRef] [PubMed]
- Sims, B.; Gu, L.; Krendelchtchikov, A.; Matthews, Q.L. Neural stem cell-derived exosomes mediate viral entry. Int. J. Nanomed. 2014, 9, 4893–4897. [Google Scholar] [CrossRef]
- Ipinmoroti, A.O.; Crenshaw, B.J.; Pandit, R.; Kumar, S.; Sims, B.; Matthews, Q.L. Human Adenovirus Serotype 3 Infection Modulates the Biogenesis and Composition of Lung Cell-Derived Extracellular Vesicles. J. Immunol. Res. 2021, 2021, 2958394. [Google Scholar] [CrossRef]
- Girard, T.J.; Antunes, L.; Zhang, N.; Amrute, J.M.; Subramanian, R.; Eldem, I.; Remy, K.E.; Mazer, M.; Erlich, E.C.; Cruchaga, C.; et al. Peripheral blood mononuclear cell tissue factor (F3 gene) transcript levels and circulating extracellular vesicles are elevated in severe coronavirus 2019 (COVID-19) disease. J. Thromb. Haemost. 2023, 21, 629–638. [Google Scholar] [CrossRef]
- Chen, L.; Chen, R.; Yao, M.; Feng, Z.; Yuan, G.; Ye, F.; Nguyen, K.; Karn, J.; McComsey, G.A.; McIntyre, T.M.; et al. COVID-19 plasma exosomes promote proinflammatory immune responses in peripheral blood mononuclear cells. Sci. Rep. 2022, 12, 21779. [Google Scholar] [CrossRef] [PubMed]
- Barnwal, A.; Basu, B.; Tripathi, A.; Soni, N.; Mishra, D.; Banerjee, A.; Kumar, R.; Vrati, S.; Bhattacharyya, J. SARS-CoV-2 Spike Protein-Activated Dendritic Cell-Derived Extracellular Vesicles Induce Antiviral Immunity in Mice. ACS Biomater. Sci. Eng. 2022, 8, 5338–5348. [Google Scholar] [CrossRef]
- Lascano, J.; Oshins, R.; Eagan, C.; Wadood, Z.; Qiang, X.; Flagg, T.; Scindia, Y.; Mehrad, B.; Brantly, M.; Khodayari, N. Correlation of alpha-1 antitrypsin levels and exosome associated neutrophil elastase endothelial injury in subjects with SARS-CoV2 infection. PLoS ONE 2022, 17, e0274427. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Steele, R.; Isbell, T.S.; Ray, R.; Ray, R.B. Circulatory Exosomes from COVID-19 Patients Trigger NLRP3 Inflammasome in Endothelial Cells. mBio 2022, 13, e0095122. [Google Scholar] [CrossRef]
- Chabert, C.; Vitte, A.L.; Iuso, D.; Chuffart, F.; Trocme, C.; Buisson, M.; Poignard, P.; Lardinois, B.; Debois, R.; Rousseaux, S.; et al. AKR1B10, One of the Triggers of Cytokine Storm in SARS-CoV2 Severe Acute Respiratory Syndrome. Int. J. Mol. Sci. 2022, 23, 1911. [Google Scholar] [CrossRef] [PubMed]
- Meidert, A.S.; Hermann, S.; Brandes, F.; Kirchner, B.; Buschmann, D.; Billaud, J.N.; Klein, M.; Lindemann, A.; Aue, E.; Schelling, G.; et al. Extracellular Vesicle Associated miRNAs Regulate Signaling Pathways Involved in COVID-19 Pneumonia and the Progression to Severe Acute Respiratory Corona Virus-2 Syndrome. Front. Immunol. 2021, 12, 784028. [Google Scholar] [CrossRef] [PubMed]
- Ching, K.L.; de Vries, M.; Gago, J.; Dancel-Manning, K.; Sall, J.; Rice, W.J.; Barnett, C.; Khodadadi-Jamayran, A.; Tsirigos, A.; Liang, F.X.; et al. ACE2-containing defensosomes serve as decoys to inhibit SARS-CoV-2 infection. PLoS Biol. 2022, 20, e3001754. [Google Scholar] [CrossRef] [PubMed]
- Tey, S.K.; Lam, H.; Wong, S.W.K.; Zhao, H.; To, K.K.; Yam, J.W.P. ACE2-enriched extracellular vesicles enhance infectivity of live SARS-CoV-2 virus. J. Extracell. Vesicles 2022, 11, e12231. [Google Scholar] [CrossRef] [PubMed]
- El-Shennawy, L.; Hoffmann, A.D.; Dashzeveg, N.K.; McAndrews, K.M.; Mehl, P.J.; Cornish, D.; Yu, Z.; Tokars, V.L.; Nicolaescu, V.; Tomatsidou, A.; et al. Circulating ACE2-expressing extracellular vesicles block broad strains of SARS-CoV-2. Nat. Commun. 2022, 13, 405. [Google Scholar] [CrossRef]
- Mezine, F.; Guerin, C.L.; Philippe, A.; Gendron, N.; Soret, L.; Sanchez, O.; Mirault, T.; Diehl, J.L.; Chocron, R.; Boulanger, C.M.; et al. Increased Circulating CD62E(+) Endothelial Extracellular Vesicles Predict Severity and in- Hospital Mortality of COVID-19 Patients. Stem Cell Rev. Rep. 2023, 19, 114–119. [Google Scholar] [CrossRef]
- Raineri, D.; Venegoni, C.; Calella, M.G.; Vaschetto, R.; Scotti, L.; Canciani, E.; Manfredi, M.; Gavelli, F.; Castello, L.; Chiocchetti, A.; et al. Worse Disease Prognosis Is Associated to an Increase of Platelet-Derived Extracellular Vesicles in Hospitalized SARS-CoV-2 Patients. Dis. Markers 2022, 2022, 8074655. [Google Scholar] [CrossRef]
- Darwish, S.; Liu, L.P.; Robinson, T.O.; Tarugu, S.; Owings, A.H.; Glover, S.C.; Alli, A.A. COVID-19 Plasma Extracellular Vesicles Increase the Density of Lipid Rafts in Human Small Airway Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 1654. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Pan, X.; Luo, R.H.; Shen, X.; Li, S.; Wang, Y.; Zuo, X.; Wu, Y.; Guo, Y.; Xiao, G.; et al. Extracellular vesicles mediate antibody-resistant transmission of SARS-CoV-2. Cell Discov. 2023, 9, 2. [Google Scholar] [CrossRef]
- Berry, F.; Morin-Dewaele, M.; Majidipur, A.; Jamet, T.; Bartier, S.; Ignjatovic, E.; Toniutti, D.; Gaspar Lopes, J.; Soyeux-Porte, P.; Maille, P.; et al. Proviral role of human respiratory epithelial cell-derived small extracellular vesicles in SARS-CoV-2 infection. J. Extracell. Vesicles 2022, 11, e12269. [Google Scholar] [CrossRef]
- Rudiansyah, M.; Terefe, E.M.; Opulencia, M.J.C.; Abdelbasset, W.K.; Bokov, D.O.; El-Sehrawy, A.A.; Baymakov, S.; Hammid, A.T.; Shirvaliloo, M.; Akhavan-Sigari, R. Type 2 alveolar epithelial cell-derived circulating extracellular vesicle-encapsulated surfactant protein C as a mediator of cardiac inflammation in COVID-19. Inflamm. Res. 2022, 71, 1003–1009. [Google Scholar] [CrossRef]
- Barberis, E.; Vanella, V.V.; Falasca, M.; Caneapero, V.; Cappellano, G.; Raineri, D.; Ghirimoldi, M.; De Giorgis, V.; Puricelli, C.; Vaschetto, R.; et al. Circulating Exosomes Are Strongly Involved in SARS-CoV-2 Infection. Front. Mol. Biosci. 2021, 8, 632290. [Google Scholar] [CrossRef]
- Setua, S.; Thangaraju, K.; Dzieciatkowska, M.; Wilkerson, R.B.; Nemkov, T.; Lamb, D.R.; Tagaya, Y.; Boyer, T.; Rowden, T.; Doctor, A.; et al. Coagulation potential and the integrated omics of extracellular vesicles from COVID-19 positive patient plasma. Sci. Rep. 2022, 12, 22191. [Google Scholar] [CrossRef] [PubMed]
- Zebardast, A.; Latifi, T.; Shabani, M.; Hasanzadeh, A.; Danesh, M.; Babazadeh, S.; Sadeghi, F. Thrombotic storm in coronavirus disease 2019: From underlying mechanisms to its management. J. Med. Microbiol. 2022, 71, 1591. [Google Scholar] [CrossRef] [PubMed]
- Moraes, E.; Martins-Goncalves, R.; da Silva, L.R.; Mandacaru, S.C.; Melo, R.M.; Azevedo-Quintanilha, I.; Perales, J.; Bozza, F.A.; Souza, T.M.L.; Castro-Faria-Neto, H.C.; et al. Proteomic Profile of Procoagulant Extracellular Vesicles Reflects Complement System Activation and Platelet Hyperreactivity of Patients with Severe COVID-19. Front. Cell Infect. Microbiol. 2022, 12, 926352. [Google Scholar] [CrossRef] [PubMed]
- Ebeyer-Masotta, M.; Eichhorn, T.; Weiss, R.; Laukova, L.; Weber, V. Activated Platelets and Platelet-Derived Extracellular Vesicles Mediate COVID-19-Associated Immunothrombosis. Front. Cell Dev. Biol. 2022, 10, 914891. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, J.; Kansakar, U.; Sardu, C.; Messina, V.; Jankauskas, S.S.; Marfella, R.; Maggi, P.; Wang, X.; Mone, P.; Paolisso, G.; et al. Exosomal miR-145 and miR-885 Regulate Thrombosis in COVID-19. J. Pharmacol. Exp. Ther. 2023, 384, 109–115. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, A.; Kaur, J.; Verma, N.; Pandey, A.K.; Das, S.; Bhattacharyya, S.; Guchhait, P. Upregulation of cytokine signalling in platelets increases risk of thrombophilia in severe COVID-19 patients. Blood Cells Mol. Dis. 2022, 94, 102653. [Google Scholar] [CrossRef]
- Strong, M.J. SARS-CoV-2, aging, and post-COVID-19 neurodegeneration. J. Neurochem. 2022, 165, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.M.; Huang, X.; Shui, G. Neurological aspects of SARS-CoV-2 infection: Lipoproteins and exosomes as Trojan horses. Trends Endocrinol. Metab. 2022, 33, 554–568. [Google Scholar] [CrossRef]
- Ahmed, S.; Paramasivam, P.; Kamath, M.; Sharma, A.; Rome, S.; Murugesan, R. Genetic Exchange of Lung-Derived Exosome to Brain Causing Neuronal Changes on COVID-19 Infection. Mol. Neurobiol. 2021, 58, 5356–5368. [Google Scholar] [CrossRef]
- Gunasekaran, M.; Bansal, S.; Ravichandran, R.; Sharma, M.; Perincheri, S.; Rodriguez, F.; Hachem, R.; Fisher, C.E.; Limaye, A.P.; Omar, A.; et al. Respiratory viral infection in lung transplantation induces exosomes that trigger chronic rejection. J. Heart Lung Transpl. 2020, 39, 379–388. [Google Scholar] [CrossRef]
- Kaushik, A.C.; Wu, Q.; Lin, L.; Li, H.; Zhao, L.; Wen, Z.; Song, Y.; Wu, Q.; Wang, J.; Guo, X.; et al. Exosomal ncRNAs profiling of mycobacterial infection identified miRNA-185-5p as a novel biomarker for tuberculosis. Brief. Bioinform. 2021, 22, bbab210. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Jiang, X.; Wu, A.; Zhou, J.; Liu, H.; He, F.; Zhang, Q.; Zen, K.; Gu, S.; Wang, J. Two Small Extracellular Vesicle sRNAs Derived from Mycobacterium tuberculosis Serve as Diagnostic Biomarkers for Active Pulmonary Tuberculosis. Front. Microbiol. 2021, 12, 642559. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Pan, L.; Shen, X.; Tian, H.; Guo, L.; Zhang, Z.; Liu, X. Hsp16.3 of mycobacterium tuberculosis in exosomes as a biomarker of tuberculosis. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2427–2430. [Google Scholar] [CrossRef]
- Rondina, M.T.; Tatsumi, K.; Bastarache, J.A.; Mackman, N. Microvesicle Tissue Factor Activity and Interleukin-8 Levels are Associated with Mortality in Patients with Influenza A/H1N1 Infection. Crit. Care Med. 2016, 44, e574–e578. [Google Scholar] [CrossRef] [PubMed]
- Shaver, C.M.; Woods, J.; Clune, J.K.; Grove, B.S.; Wickersham, N.E.; McNeil, J.B.; Shemancik, G.; Ware, L.B.; Bastarache, J.A. Circulating microparticle levels are reduced in patients with ARDS. Crit. Care 2017, 21, 120. [Google Scholar] [CrossRef]
- Huang, F.; Bai, J.; Zhang, J.; Yang, D.; Fan, H.; Huang, L.; Shi, T.; Lu, G. Identification of potential diagnostic biomarkers for pneumonia caused by adenovirus infection in children by screening serum exosomal microRNAs. Mol. Med. Rep. 2019, 19, 4306–4314. [Google Scholar] [CrossRef]
- Tertel, T.; Tomic, S.; Dokic, J.; Radojevic, D.; Stevanovic, D.; Ilic, N.; Giebel, B.; Kosanovic, M. Serum-derived extracellular vesicles: Novel biomarkers reflecting the disease severity of COVID-19 patients. J. Extracell. Vesicles 2022, 11, e12257. [Google Scholar] [CrossRef]
- Fujita, Y.; Hoshina, T.; Matsuzaki, J.; Yoshioka, Y.; Kadota, T.; Hosaka, Y.; Fujimoto, S.; Kawamoto, H.; Watanabe, N.; Sawaki, K.; et al. Early prediction of COVID-19 severity using extracellular vesicle COPB2. J. Extracell. Vesicles 2021, 10, e12092. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Pathogens | Effect | References |
|---|---|---|
| Mycobacterium tuberculosis | Infected Mφ EVs: + (CCL2, VCAM1) in endothelial cells ++ Peptide-MHC-II complexes in T-cells Serum EVs: ++ Death rate in THP-1 & PBMCs Infected neutrophils EVs: ++ Mφ autophagy ++ Mφ superoxide anion ++ Mφ bacterial killing ++ (TNF-α, IL-6, IL-10) in Mφ Mtb RNA in EVs: ++ IFN in Mφ Spontaneously released EVs: -- MCP-1, -- TNF-α, -- bacterial load in mtb-infected Mφ Mtb-EVs & S-EVs: --- Bacterial load in Mtb-infected mice. | [39,41,42,43,45] |
| Mycobacterium bovis | Infected THP-1 EVs: +++ (IL-12p40, TNF) Infected Mφ EVs ++ Peptide-MHC-II complexes in T-cells + Antigen-specific activation of CD4+ & CD8+ T cells in mice | [40,45,46] |
| Streptococcus pneumoniae | Pneumolysin+ Neutrophils EVs: ++ CD62P in platelets Pneumolysin+ Lung epithelial cells microvesicles: -- ROS in neutrophils | [49,50] |
| Staphylococcus aureus | Infected neutrophils EVs: ++ (IL-6, IL-1β) in Mφ | [52] |
| Pseudomonas aeruginosa | Let-7b-gb miRNA in EVs: - Biofilm formation, + sensitivity to beta-lactam antibiotic (Aztreonam) | [53] |
| Legionella penuemophila | Supernatant EVs: ++ (CXCL8, TNF-α, IL-1β, MCP-1) in THP-1 | [54] |
| Influenza A | Infected lung cells EVs: +++ (Type I IFN, IL-12p40, IL-6) in dendritic cells Blocking viral replication in A549 infected cells Infected A549 cells EVs: Neutralizing virus particles Infected lung Mφ EVs: ++ (Type I IFN, IL-6, CCL2, TNF-α) in epithelial cells Alveolar macrophages EVs: Acceleration of endosomal acidification in epithelial cells | [57,58,59,61,62,65] |
| Respiratory syncytial virus | Infected Epithelial cells exosomes showed +(CXCL10 and CCL5). ++ (MCP-1, IP-10 & RANTES) in monocytes & A549 cells ++ TNF-α in A549 cells | [68,71] |
| Rhinovirus | + (IFN-γ & IP-10) in Th1 + IL-13 in Th2 | [72] |
| Adenovirus | Adenovirus 3 EVs: +++ Rab5, 7 & 35 | [75] |
| SARS-CoV-2 | + (IL-6, IL-8 and TNF-α) ++ neurtrophil elastase-related endothelial damage Aldo-Keto Reductase 1B10-related cytokine storm - IL-8 (ACE2)+-related viral entry block CD62E+-related hospital mortality Platelets EVs-related worsened prognosis and mortality Lipid raft formation in human small airways Active viral particles-related disease transmission Epithelial EVS-related cardiac inflammation Coagulation Platelet-derived EVs cytokine signaling upregulation-related thrombophilia Fastened fibril formation and progression in neurons, astrocytes and microglia Transcription factors transmission-related neurodegenerative processes | [77,79,81,83,84,85,86,87,88,89,90,91,92,93,94,95,96,98,99,100,101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hambo, S.; Harb, H. Extracellular Vesicles and Their Role in Lung Infections. Int. J. Mol. Sci. 2023, 24, 16139. https://doi.org/10.3390/ijms242216139
Hambo S, Harb H. Extracellular Vesicles and Their Role in Lung Infections. International Journal of Molecular Sciences. 2023; 24(22):16139. https://doi.org/10.3390/ijms242216139
Chicago/Turabian StyleHambo, Shadi, and Hani Harb. 2023. "Extracellular Vesicles and Their Role in Lung Infections" International Journal of Molecular Sciences 24, no. 22: 16139. https://doi.org/10.3390/ijms242216139
APA StyleHambo, S., & Harb, H. (2023). Extracellular Vesicles and Their Role in Lung Infections. International Journal of Molecular Sciences, 24(22), 16139. https://doi.org/10.3390/ijms242216139