
The Antioxidant and Immunomodulatory Potential of Coccoloba alnifolia Leaf Extracts

 , , ,
, , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
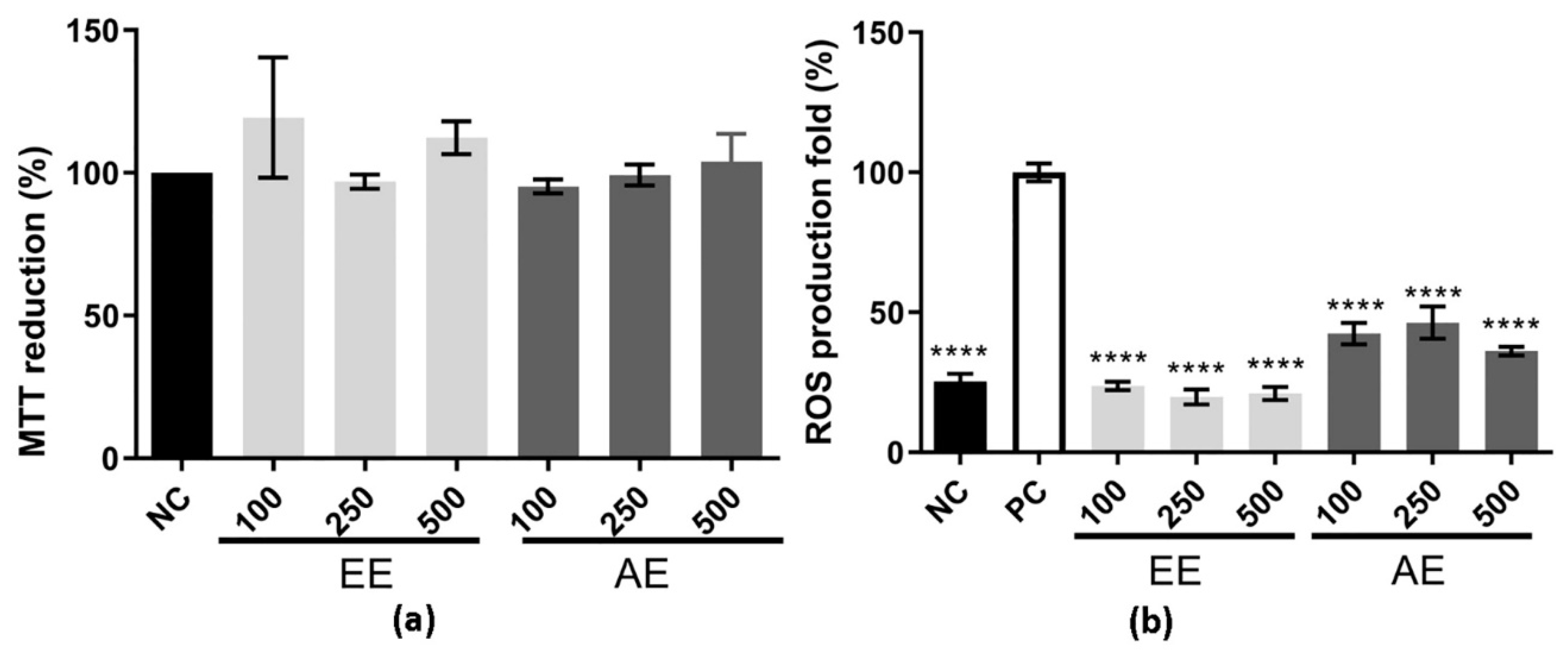
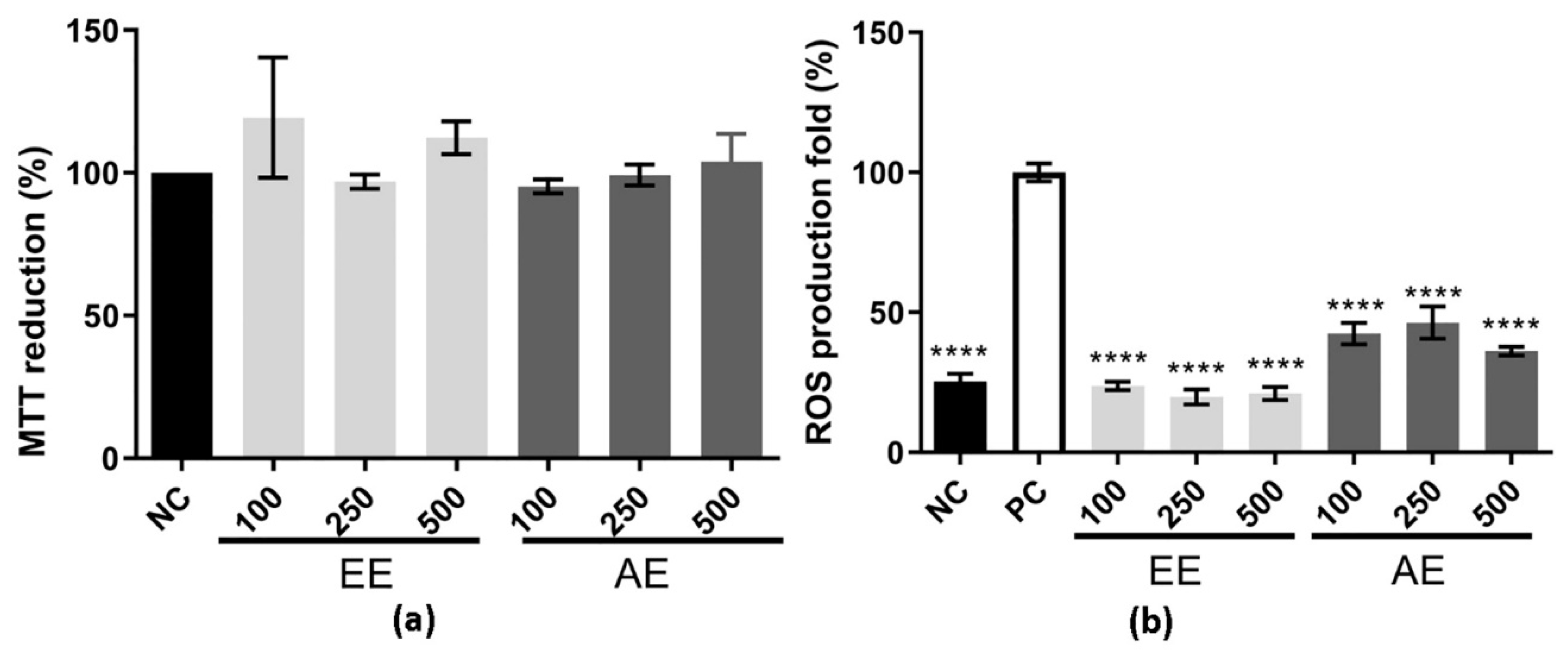
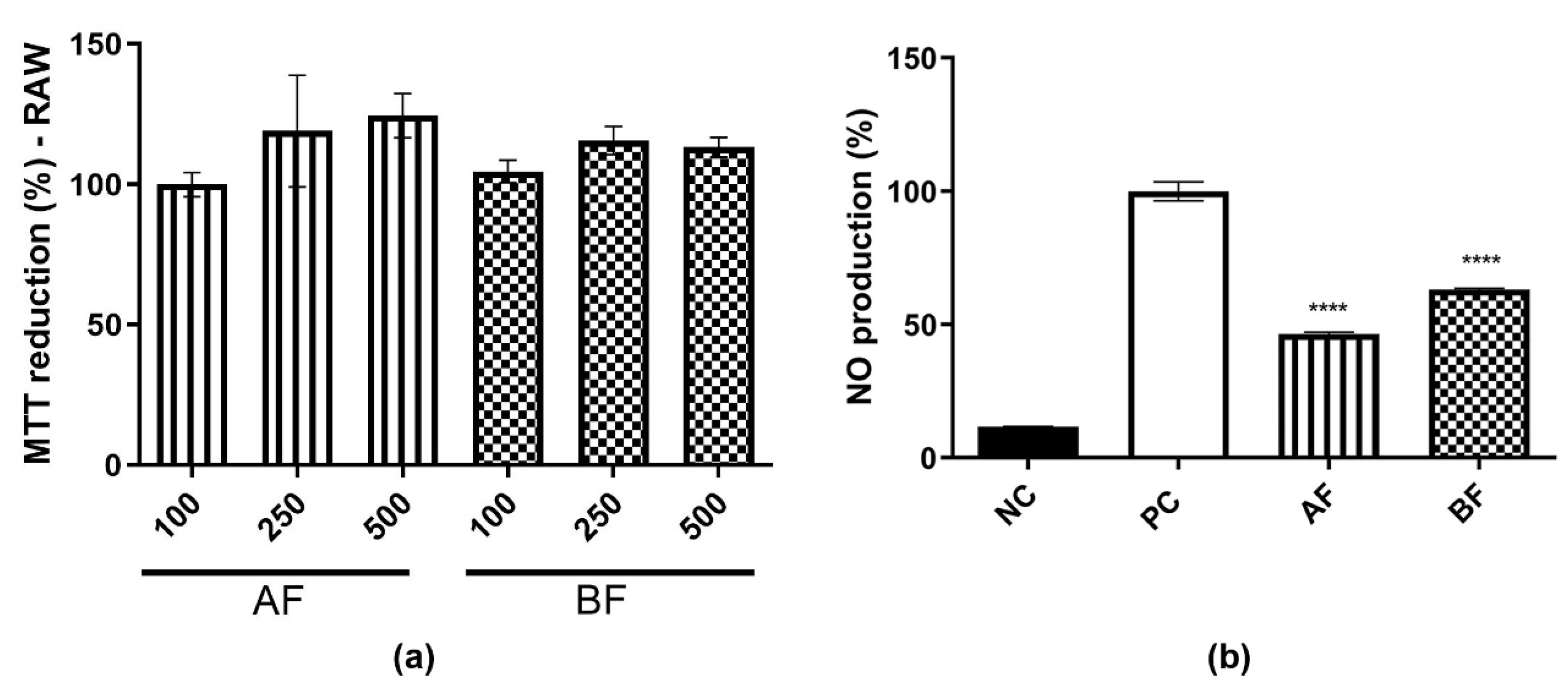
2.1. MTT Reduction and Intracellular ROS Production Assay
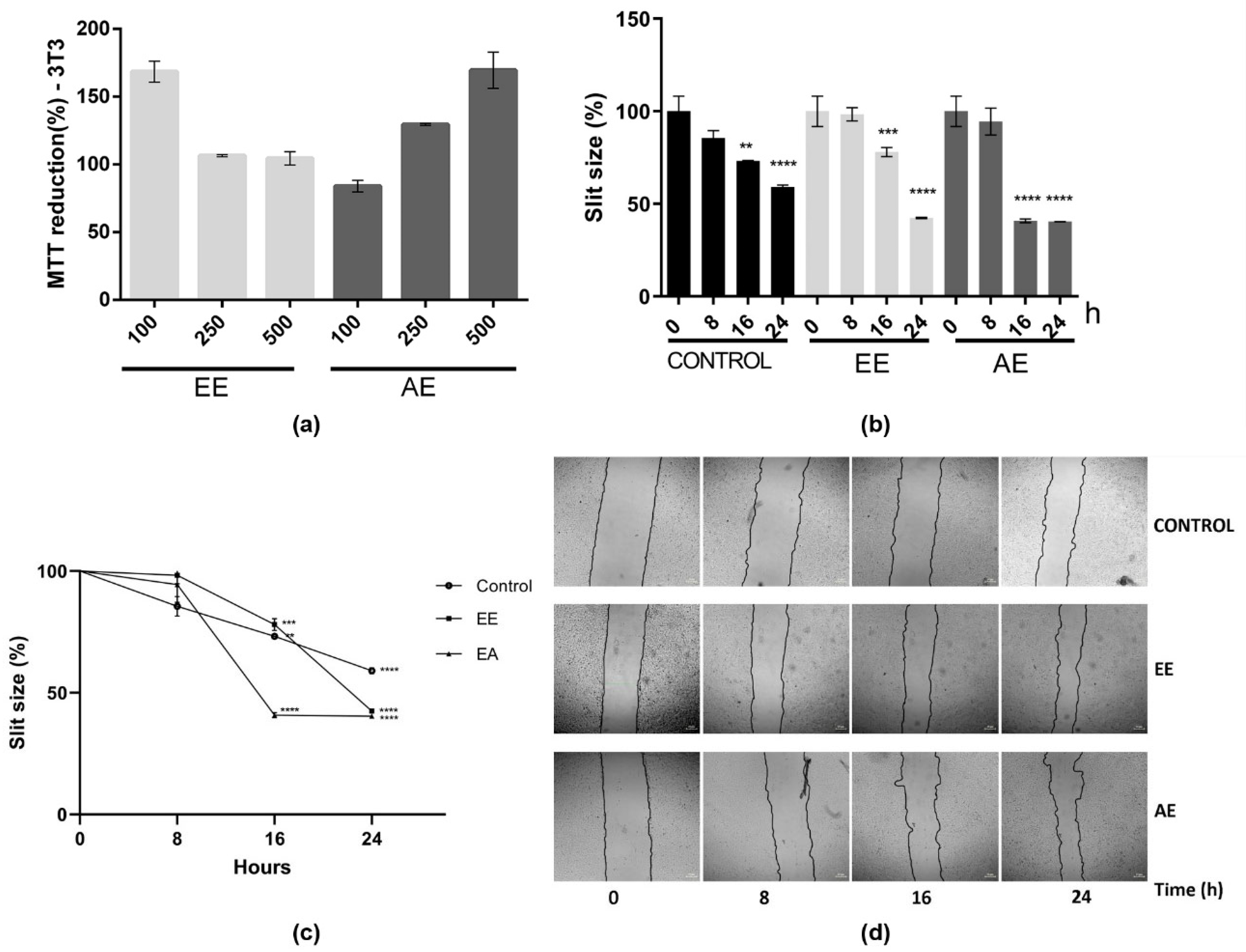
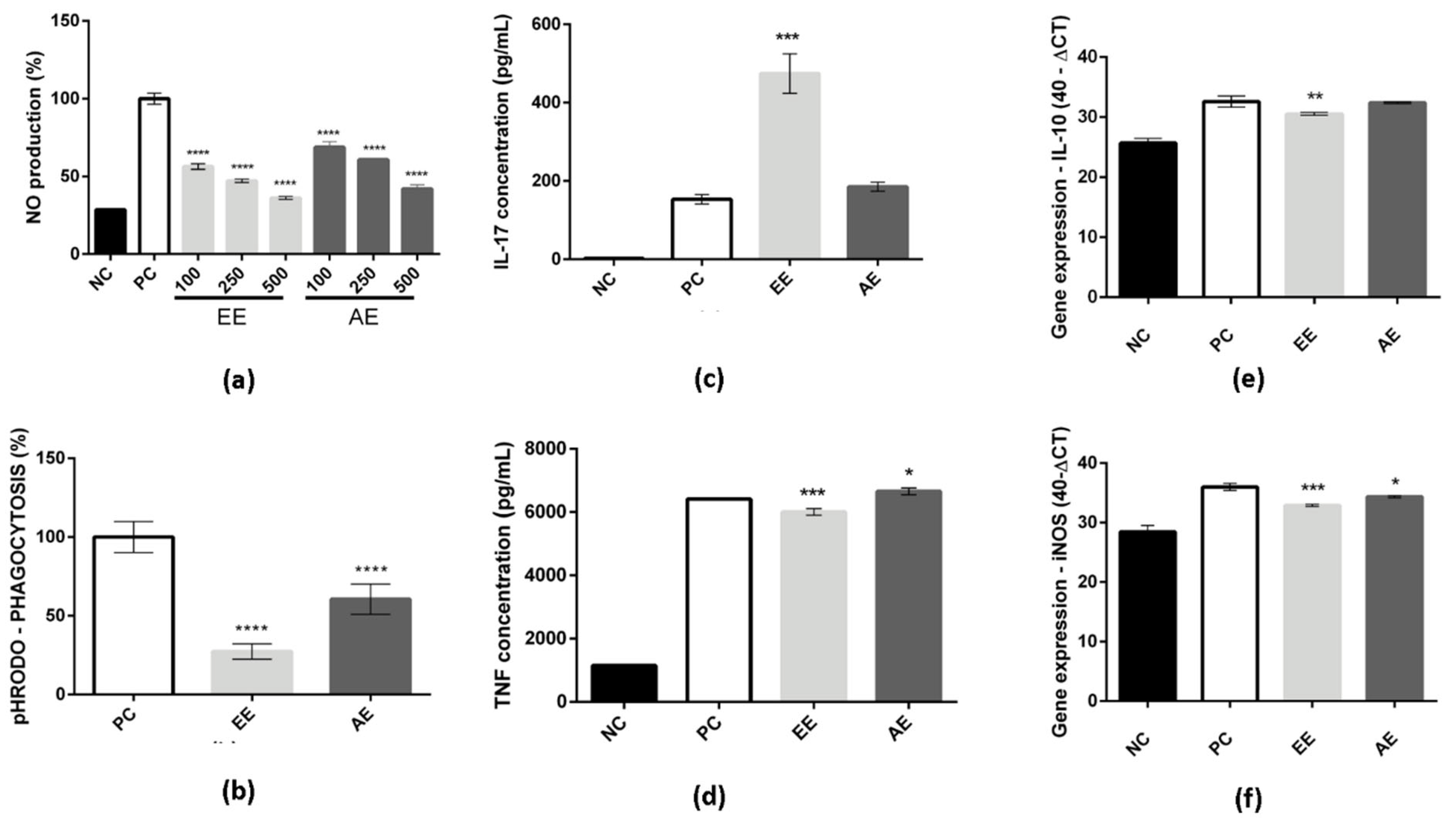
2.2. EE and AE Immunomodulatory Activity
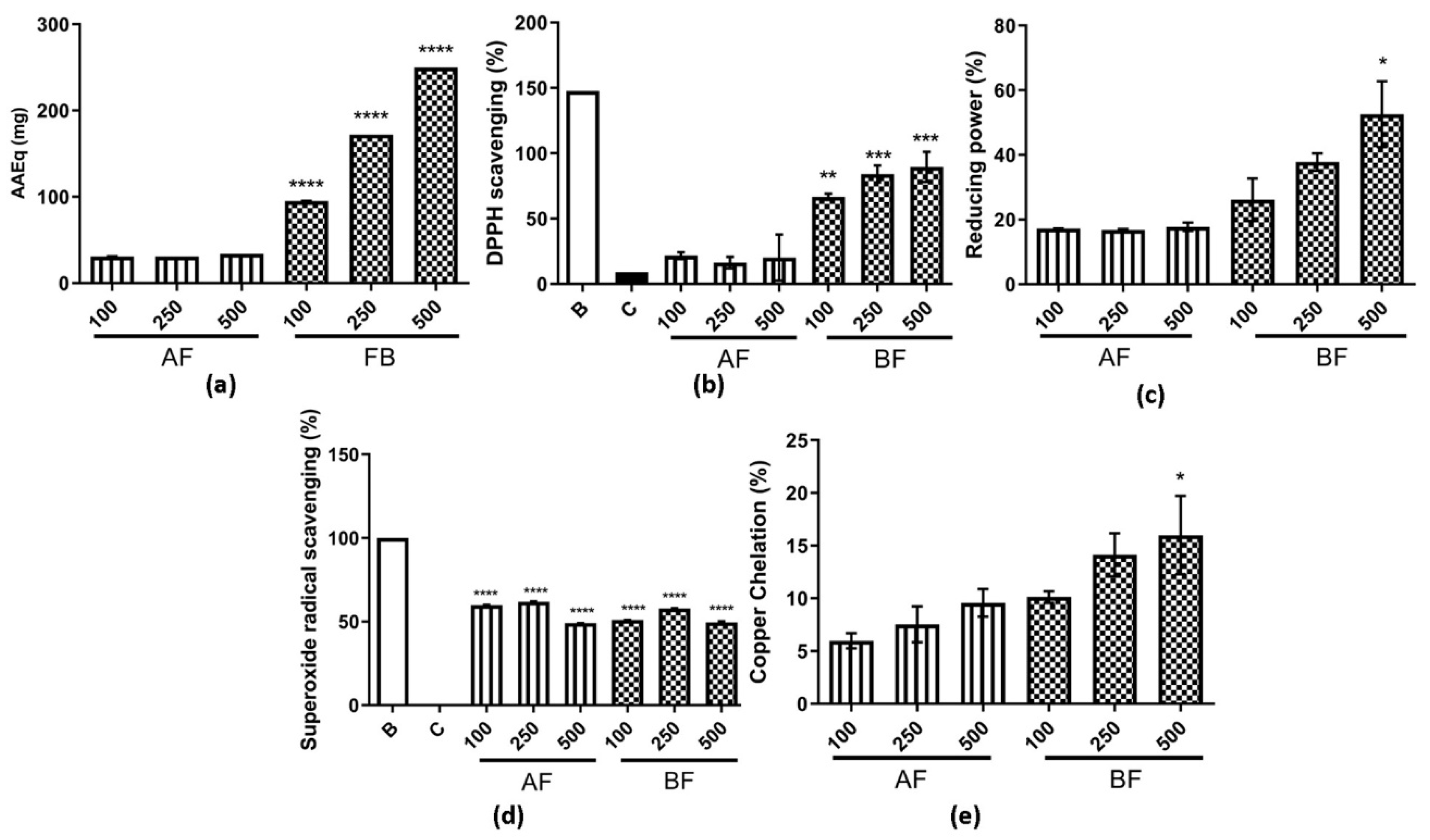
2.3. AE Partition Phases Antioxidant Activity Characterization
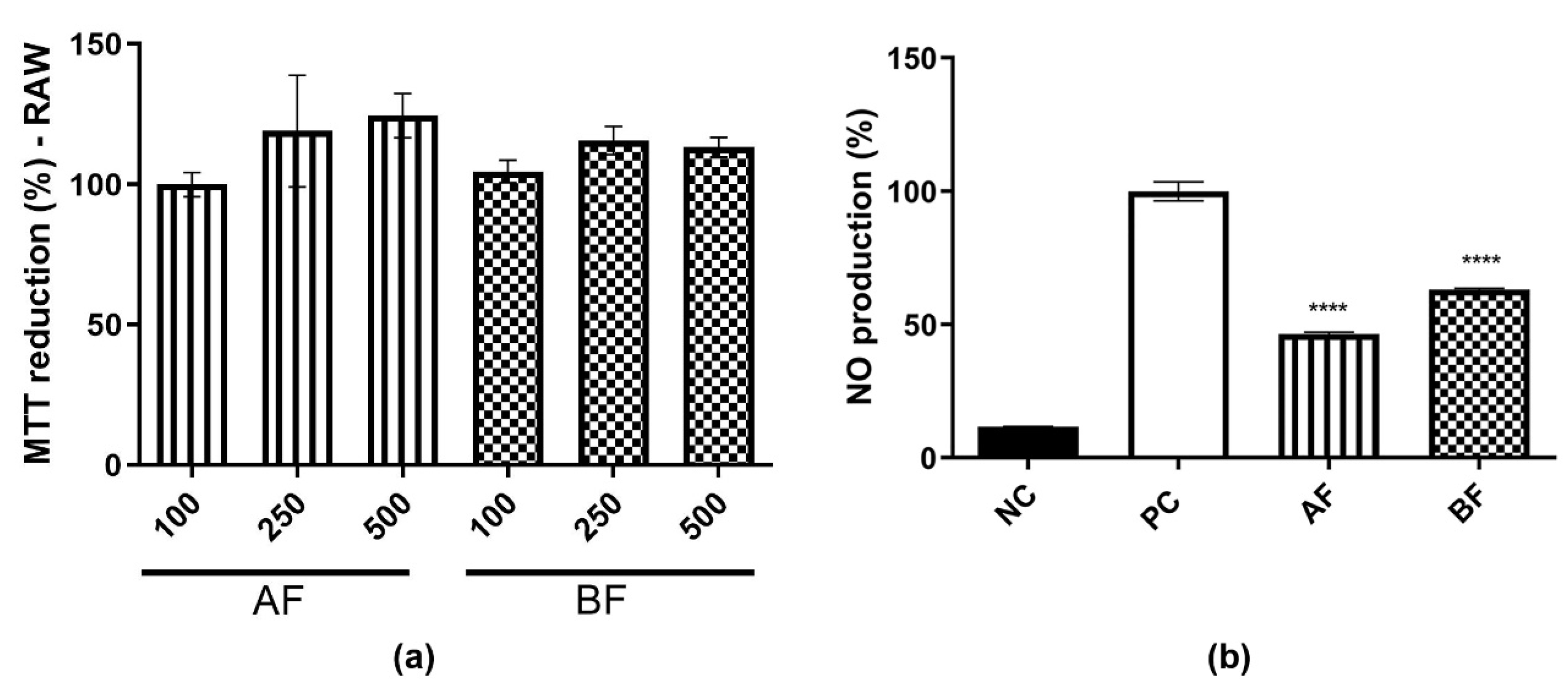
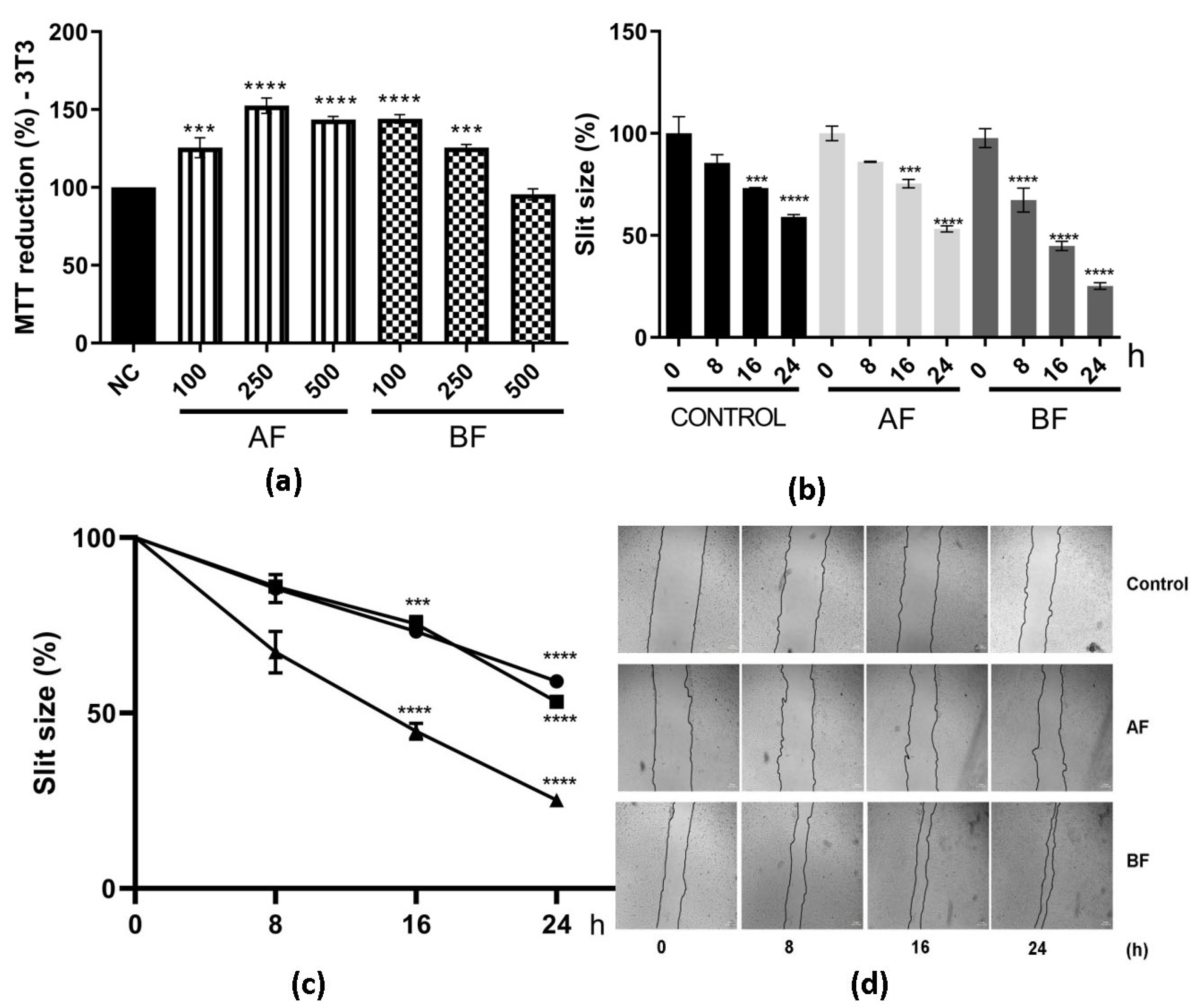
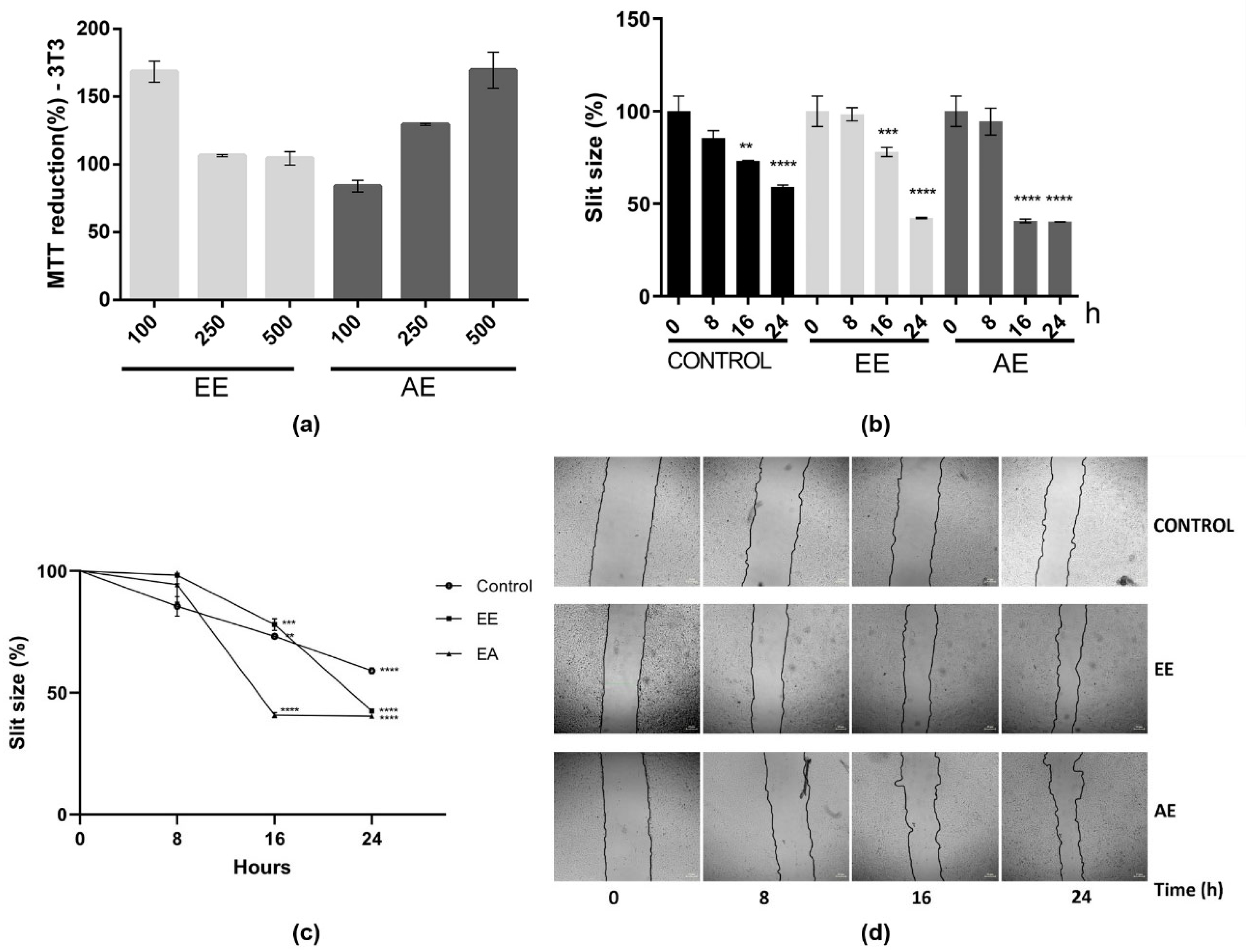
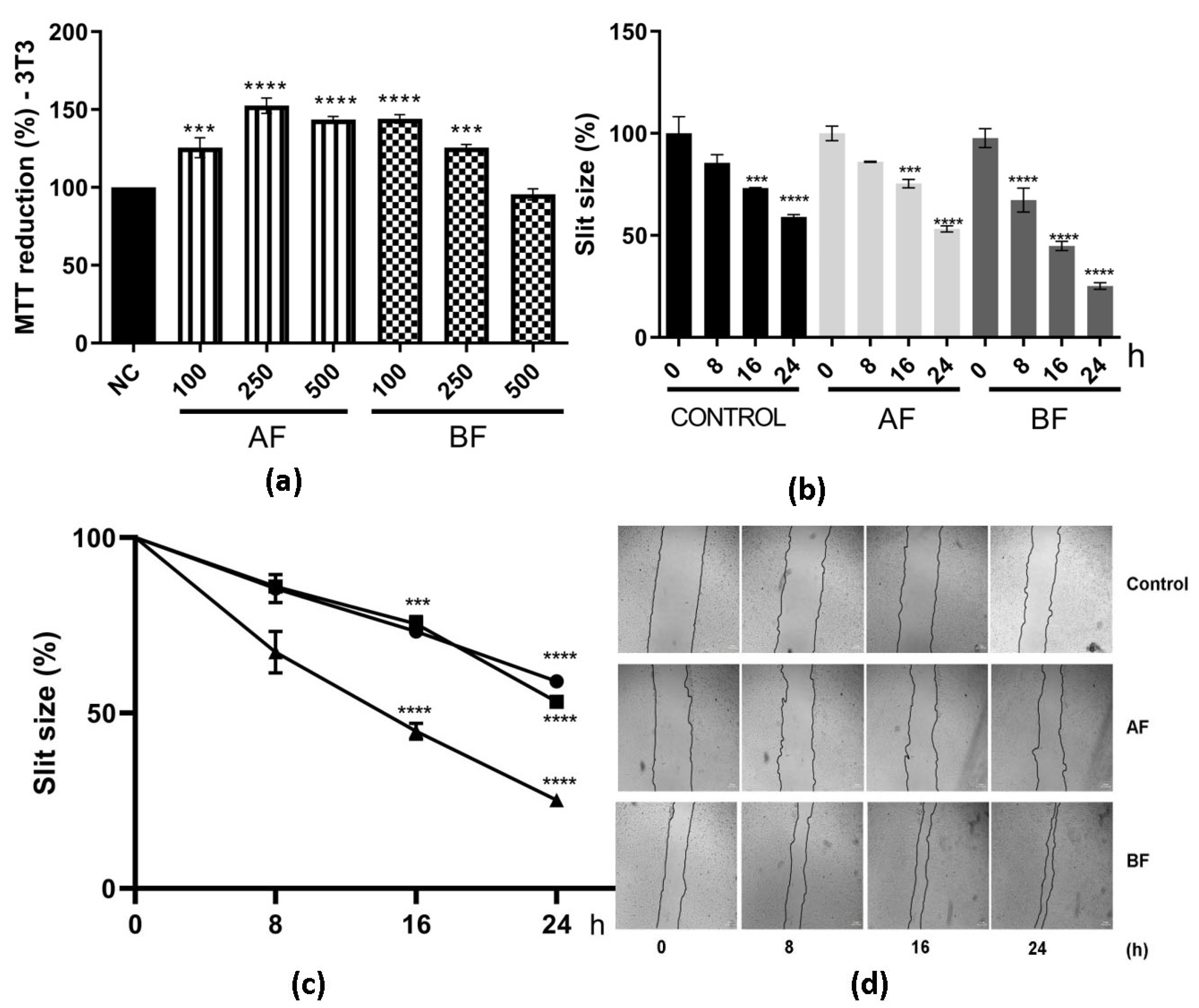
2.4. C. alnifolia Partition Phases Immunomodulatory Activity
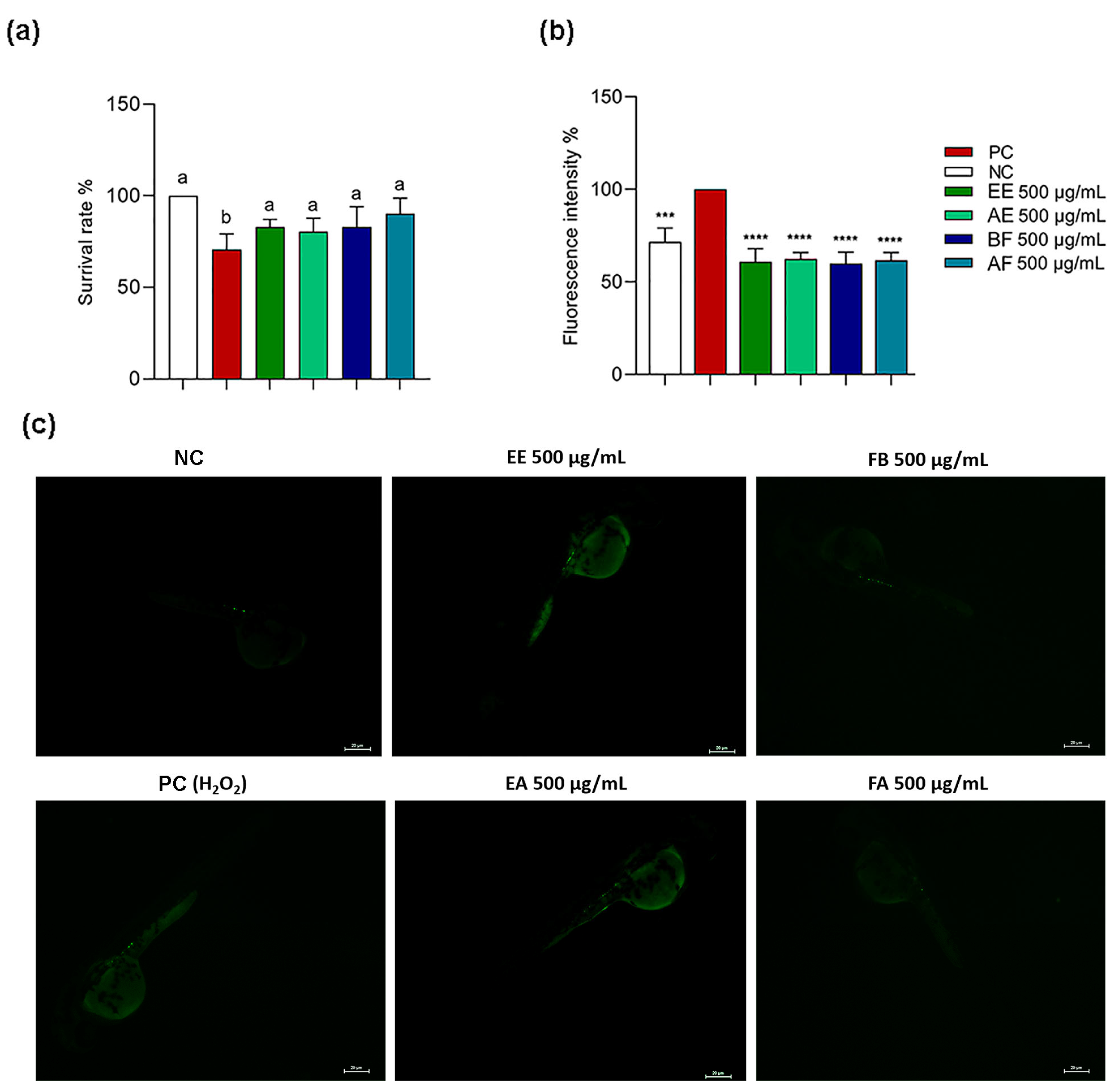
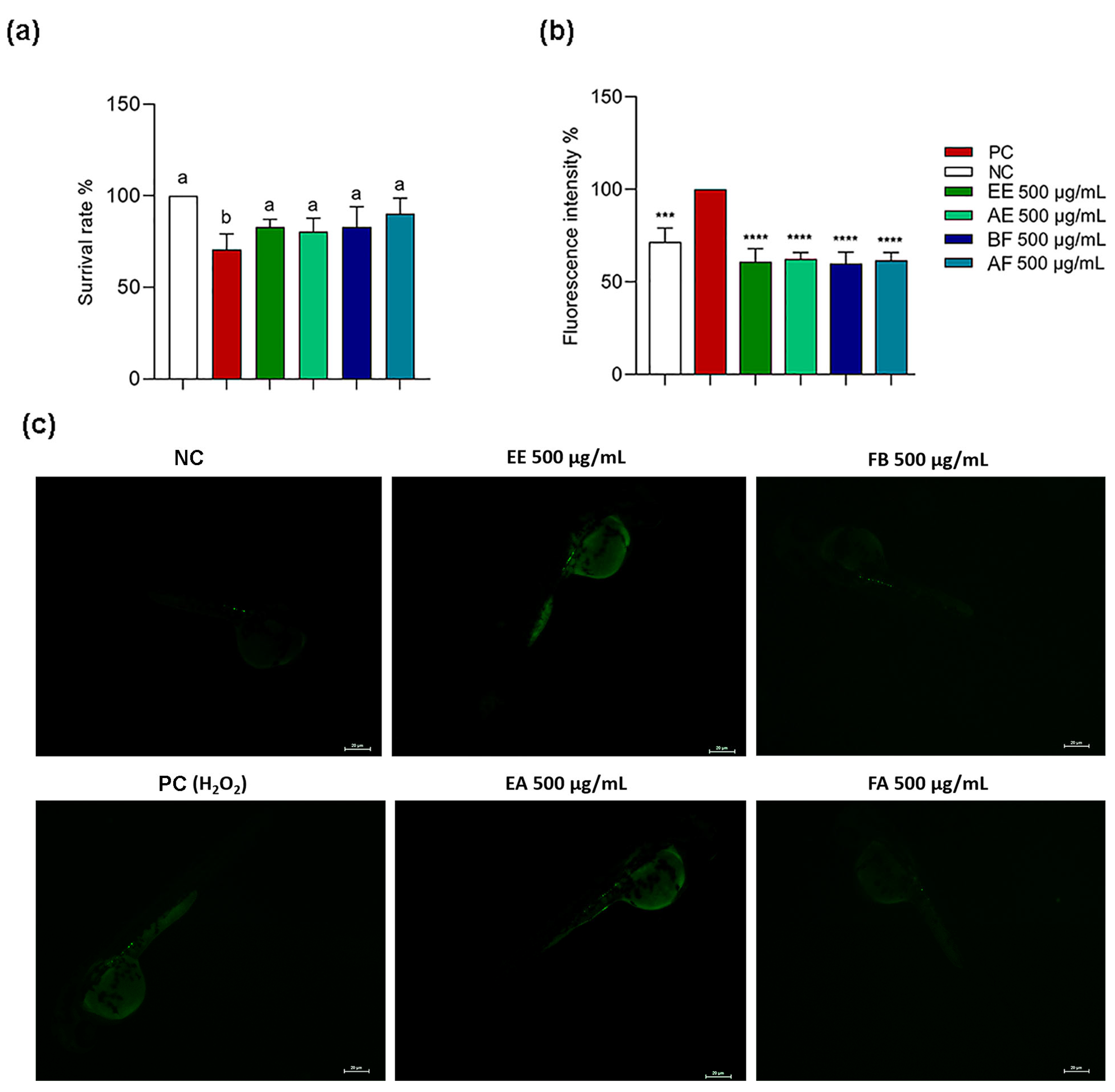
2.5. C. alnifolia Extracts and Partition Phase Characterization Using Zebrafish Model
2.6. BF Partition Phase Characterization
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Biological Material
4.3. Extraction
4.4. Lineage and Cell Culture
4.5. MTT Reduction Assay
4.6. Production of Intracellular ROS
4.7. Dosage of Nitric Oxide
4.8. Phagocytic Activity
4.9. RNA Extraction and Quantitative PCR (qRT-PCR)
4.10. Dosage of Cytokines
4.11. In Vitro Scratch Assay
4.12. Antioxidant Assays
4.12.1. Total Antioxidant Capacity (TAC)
4.12.2. DPPH
4.12.3. Reducing Power
4.12.4. Superoxide Scavenging Activity
4.12.5. Copper Chelating
4.13. Statistical Analysis
4.14. Partition Phase Analysis in HPLC Ms/Ms
4.15. Molecular Network
4.16. Measurement of Intracellular ROS Induced by Oxidative Stress Generation and Image Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medzhitov, R. Inflammation 2010: New Adventures of an Old Flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Tasneem, S.; Liu, B.; Li, B.; Choudhary, M.I.; Wang, W. Molecular Pharmacology of Inflammation: Medicinal Plants as Anti-Inflammatory Agents. Pharmacol. Res. 2019, 139, 126–140. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as Anti-Inflammatory Agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Wei, Y.; Hu, T. Flavonoids of Polygonum hydropiper L. Attenuates Lipopolysaccharide-Induced Inflammatory Injury via Suppressing Phosphorylation in MAPKs Pathways. BMC Complement. Altern. Med. 2015, 16, 25. [Google Scholar] [CrossRef]
- Yue, Y.; Shen, P.; Xu, Y.; Park, Y. p-Coumaric Acid Improves Oxidative and Osmosis Stress Responses in Caenorhabditis elegans: Oxidative and Osmosis Stress Responses in Caenorhabditis elegans. J. Sci. Food Agric. 2019, 99, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, 2004433. [Google Scholar] [CrossRef] [PubMed]
- Kamoto, K.; Chiwaya, A.; Nambala, P.; Chammudzi, P.; Senga, E.; Chisi, J.; Matovu, E.; Musaya, J. Plasma Cytokenes Quantification among Trypanosoma brucei Rhodesiense Sleeping Sickness Cases and Controls in Rumphi, Malawi. Mal. Med. J. 2021, 33, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Apaza Ticona, L.; Slowing, K.; Serban, A.M.; Humanes Bastante, M.; Hernáiz, M.J. Wound Healing, Anti-Inflammatory and Anti-Melanogenic Activities of Ursane-Type Triterpenes from Semialarium mexicanum (Miers) Mennega. J. Ethnopharmacol. 2022, 289, 115009. [Google Scholar] [CrossRef]
- Vaid, B.; Chopra, B.S.; Raut, S.; Sagar, A.; Badmalia, M.D.; Ashish; Khatri, N. Antioxidant and Wound Healing Property of Gelsolin in 3T3-L1 Cells. Oxidative Med. Cell. Longev. 2020, 2020, 4045365. [Google Scholar] [CrossRef] [PubMed]
- Che Zain, M.S.; Lee, S.Y.; Sarian, M.N.; Fakurazi, S.; Shaari, K. In Vitro Wound Healing Potential of Flavonoid C-Glycosides from Oil Palm (Elaeis guineensis Jacq.) Leaves on 3T3 Fibroblast Cells. Antioxidants 2020, 9, 326. [Google Scholar] [CrossRef]
- De Freitas Figueiredo, F.; Cechinel Filho, V.; Damazo, A.S.; Arunachalam, K.; Colodel, E.M.; Ribeiro, M.; Venturini, C.L.; Oliveira, D.M.; Machado, M.T.M.; Pavan, E.; et al. Sorocea Guilleminiana Gaudich.: Wound Healing Activity, Action Mechanisms, and Chemical Characterization of the Leaf Infusion. J. Ethnopharmacol. 2020, 248, 112307. [Google Scholar] [CrossRef] [PubMed]
- Sarkhail, P.; Navidpour, L.; Rahimifard, M.; Hosseini, N.M.; Souri, E. Bioassay-Guided Fractionation and Identification of Wound Healing Active Compound from Pistacia vera L. Hull Extract. J. Ethnopharmacol. 2020, 248, 112335. [Google Scholar] [CrossRef]
- Kaleci, B.; Koyuturk, M. Efficacy of Resveratrol in the Wound Healing Process by Reducing Oxidative Stress and Promoting Fibroblast Cell Proliferation and Migration. Dermatol. Ther. 2020, 33, e14357. [Google Scholar] [CrossRef]
- Khan, T.; Ali, M.; Khan, A.; Nisar, P.; Jan, S.A.; Afridi, S.; Shinwari, Z.K. Anticancer Plants: A Review of the Active Phytochemicals, Applications in Animal Models, and Regulatory Aspects. Biomolecules 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Thomford, N.; Senthebane, D.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Koehn, F.E.; Carter, G.T. The Evolving Role of Natural Products in Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef]
- Oliveira, P.V.; Ferreira, J.C.; Moura, F.S.; Lima, G.S.; de Oliveira, F.M.; Oliveira, P.E.S.; Conserva, L.M.; Giulietti, A.M.; Lemos, R.P.L. Larvicidal Activity of 94 Extracts from Ten Plant Species of Northeastern of Brazil against Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res. 2010, 107, 403–407. [Google Scholar] [CrossRef]
- Kaewpiboon, C.; Lirdprapamongkol, K.; Srisomsap, C.; Winayanuwattikun, P.; Yongvanich, T.; Puwaprisirisan, P.; Svasti, J.; Assavalapsakul, W. Studies of the in Vitro Cytotoxic, Antioxidant, Lipase Inhibitory and Antimicrobial Activities of Selected Thai Medicinal Plants. BMC Complement. Altern. Med. 2012, 12, 217. [Google Scholar] [CrossRef]
- de Barros, I.B.; de Souza Daniel, J.F.; Pinto, J.P.; Rezende, M.I.; Braz Filho, R.; Ferreira, D.T. Phytochemical and Antifungal Activity of Anthraquinones and Root and Leaf Extracts of Coccoloba mollis on Phytopathogens. Braz. Arch. Biol. Technol. 2011, 54, 535–541. [Google Scholar] [CrossRef]
- Tsuboy, M.S.; Marcarini, J.C.; Luiz, R.C.; Barros, I.B.; Ferreira, D.T.; Ribeiro, L.R.; Mantovani, M.S. In Vitro Evaluation of the Genotoxic Activity and Apoptosis Induction of the Extracts of Roots and Leaves from the Medicinal Plant Coccoloba mollis (Polygonaceae). J. Med. Food 2010, 13, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.E.P.S.; Pereda, M.d.C.V.; Eberlin, S.; Dieamant, G.C.; Di Stasi, L.C. Effects of Coccoloba uvifera L. on UV-Stimulated Melanocytes. Photodermatol. Photoimmunol. Photomed. 2008, 24, 308–313. [Google Scholar] [CrossRef]
- Cota, B.B.; de Oliveira, A.B.; de Souza-Filho, J.D.; Braga, F.C. Antimicrobial Activity and Constituents of Coccoloba acrostichoides. Fitoterapia 2003, 74, 729–731. [Google Scholar] [CrossRef] [PubMed]
- de Melo, L.F.M.; Gomes, D.L.; da Silva, L.F.; Silva, L.M.P.; Machado, M.L.; Cadavid, C.O.M.; Zucolotto, S.M.; de Paula Oliveira, R.; dos Santos, D.Y.A.C.; Rocha, H.A.O.; et al. Coccoloba alnifolia Leaf Extract as a Potential Antioxidant Molecule Using In Vitro and In Vivo Assays. Oxidative Med. Cell. Longev. 2020, 2020, 3928706. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. Targeting Oxidative Stress in Disease: Promise and Limitations of Antioxidant Therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, J.; Lu, W. The Significance of Mitochondrial Dysfunction in Cancer. Int. J. Mol. Sci. 2020, 21, 5598. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, S.; Ohnishi, S.; Ma, N.; Hiraku, Y.; Murata, M. Crosstalk between DNA Damage and Inflammation in the Multiple Steps of Carcinogenesis. Int. J. Mol. Sci. 2017, 18, 1808. [Google Scholar] [CrossRef] [PubMed]
- Bermas, B.L. Non-Steroidal Anti Inflammatory Drugs, Glucocorticoids and Disease Modifying Anti-Rheumatic Drugs for the Management of Rheumatoid Arthritis before and during Pregnancy. Curr. Opin. Rheumatol. 2014, 26, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.S.; Corrêa, M.A.; Chung, M.C.; dos Santos, J.L. Synthesis, Antioxidant and Photoprotection Activities of Hybrid Derivatives Useful to Prevent Skin Cancer. Bioorganic Med. Chem. 2014, 22, 2733–2738. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A. Anti-Inflammatory Activity of Natural Products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.P.; Arruda, C.; Abd El-Salam, M.; Bastos, J.K. Brazilian Medicinal Plants with Corroborated Anti-Inflammatory Activities: A Review. Pharm. Biol. 2018, 56, 253–268. [Google Scholar] [CrossRef]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An Overview on the Anti-Inflammatory Potential and Antioxidant Profile of Eugenol. Oxidative Med. Cell. Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.S.; Spolidorio, L.C.; Manthey, J.A.; Cesar, T.B. Citrus Flavanones Prevent Systemic Inflammation and Ameliorate Oxidative Stress in C57BL/6J Mice Fed High-Fat Diet. Food Funct. 2016, 7, 2675–2681. [Google Scholar] [CrossRef] [PubMed]
- Asgharpour, F.; Moghadamnia, A.A.; Motallebnejad, M.; Nouri, H.R. Propolis Attenuates Lipopolysaccharide-induced Inflammatory Responses through Intracellular ROS and NO Levels along with Downregulation of IL-1β and IL-6 Expressions in Murine RAW 264.7 Macrophages. J. Food Biochem. 2019, 43, e12926. [Google Scholar] [CrossRef]
- Yang, R.; Yuan, B.-C.; Ma, Y.-S.; Zhou, S.; Liu, Y. The Anti-Inflammatory Activity of Licorice, a Widely Used Chinese Herb. Pharm. Biol. 2017, 55, 5–18. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Min, J.-W.; Kong, W.-L.; He, X.-H.; Li, J.-X.; Peng, B.-W. A Review on the Pharmacological Effects of Vitexin and Isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Rosa, F.; Arruda, A.; Siqueira, E.; Arruda, S. Phytochemical Compounds and Antioxidant Capacity of Tucum-Do-Cerrado (Bactris setosa Mart), Brazil’s Native Fruit. Nutrients 2016, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Rasoanaivo, P.; Wright, C.W.; Willcox, M.L.; Gilbert, B. Whole Plant Extracts versus Single Compounds for the Treatment of Malaria: Synergy and Positive Interactions. Malar. J. 2011, 10 (Suppl. S1), S4. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Cech, N.B. Synergy and Antagonism in Natural Product Extracts: When 1 + 1 Does Not Equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Sun, F.; Ruan, J.; Yan, J.; Huang, P.; Wang, J.; Han, L.; Zhang, Y.; Wang, T. Anti-Inflammatory Constituents from Cortex dictamni. Fitoterapia 2019, 134, 465–473. [Google Scholar] [CrossRef]
- Kheiry, M.; Dianat, M.; Badavi, M.; Mard, S.A.; Bayati, V. P-Coumaric Acid Attenuates Lipopolysaccharide-Induced Lung Inflammation in Rats by Scavenging ROS Production: An In Vivo and In Vitro Study. Inflammation 2019, 42, 1939–1950. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.-B.; Jung, W.-K.; Park, S.-J.; Kim, Y.-T.; Kim, W.-S.; Je, J.-Y. Gallic Acid-g-Chitosan Modulates Inflammatory Responses in LPS-Stimulated RAW264.7 Cells Via NF-ΚB, AP-1, and MAPK Pathways. Inflammation 2016, 39, 366–374. [Google Scholar] [CrossRef]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-Inflammatory Potential of Ellagic Acid, Gallic Acid and Punicalagin A&B Isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic Acid: Pharmacological Activities and Molecular Mechanisms Involved in Inflammation-Related Diseases. Biomed. Pharmacother. 2021, 133, 110985. [Google Scholar] [CrossRef] [PubMed]
- Peluso, I.; Miglio, C.; Morabito, G.; Ioannone, F.; Serafini, M. Flavonoids and Immune Function in Human: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Chansiw, N.; Chotinantakul, K.; Srichairatanakool, S. Anti-Inflammatory and Antioxidant Activities of the Extracts from Leaves and Stems of Polygonum odoratum Lour. AIAAMC 2019, 18, 45–54. [Google Scholar] [CrossRef]
- Nam, T.G.; Lim, T.-G.; Lee, B.H.; Lim, S.; Kang, H.; Eom, S.H.; Yoo, M.; Jang, H.W.; Kim, D.-O. Comparison of Anti-Inflammatory Effects of Flavonoid-Rich Common and Tartary Buckwheat Sprout Extracts in Lipopolysaccharide-Stimulated RAW 264.7 and Peritoneal Macrophages. Oxidative Med. Cell. Longev. 2017, 2017, 9658030. [Google Scholar] [CrossRef] [PubMed]
- Hossen, M.J.; Baek, K.-S.; Kim, E.; Yang, W.S.; Jeong, D.; Kim, J.H.; Kweon, D.-H.; Yoon, D.H.; Kim, T.W.; Kim, J.-H.; et al. In Vivo and in Vitro Anti-Inflammatory Activities of Persicaria chinensis Methanolic Extract Targeting Src/Syk/NF-ΚB. J. Ethnopharmacol. 2015, 159, 9–16. [Google Scholar] [CrossRef]
- Malar, D.S.; Suryanarayanan, V.; Prasanth, M.I.; Singh, S.K.; Balamurugan, K.; Devi, K.P. Vitexin Inhibits Aβ25-35 Induced Toxicity in Neuro-2a Cells by Augmenting Nrf-2/HO-1 Dependent Antioxidant Pathway and Regulating Lipid Homeostasis by the Activation of LXR-α. Toxicol. Vitr. 2018, 50, 160–171. [Google Scholar] [CrossRef]
- Pasparakis, M. Regulation of Tissue Homeostasis by NF-ΚB Signalling: Implications for Inflammatory Diseases. Nat. Rev. Immunol. 2009, 9, 778–788. [Google Scholar] [CrossRef]
- Burian, E.A.; Sabah, L.; Karlsmark, T.; Kirketerp-Møller, K.; Moffatt, C.J.; Thyssen, J.P.; Ågren, M.S. Cytokines and Venous Leg Ulcer Healing—A Systematic Review. Int. J. Mol. Sci. 2022, 23, 6526. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; He, L.; Chen, Y.; Wu, L.; Wang, L.; Liu, Z. Anti-Inflammatory and Antioxidative Effects of Camellia oleifera Abel Components. Future Med. Chem. 2017, 9, 2069–2079. [Google Scholar] [CrossRef]
- Abdel Hakim, F.; Gad, H.; Radwan, R.; Ayoub, N.; El-Shazly, M. Biological and Phytochemical Review on the Genus Coccoloba (Polygonaceae). Arch. Pharm. Sci. Ain Shams Univ. 2019, 3, 180–194. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I. Functional Foods for Health: The Interrelated Antioxidant and Anti-Inflammatory Role of Fruits, Vegetables, Herbs, Spices and Cocoa in Humans. Curr. Pharm. Des. 2017, 22, 6701–6715. [Google Scholar] [CrossRef] [PubMed]
- Kalmarzi, R.N.; Naleini, S.N.; Ashtary-Larky, D.; Peluso, I.; Jouybari, L.; Rafi, A.; Ghorat, F.; Heidari, N.; Sharifian, F.; Mardaneh, J.; et al. Anti-Inflammatory and Immunomodulatory Effects of Barberry (Berberis vulgaris) and Its Main Compounds. Oxidative Med. Cell. Longev. 2019, 2019, 6183965. [Google Scholar] [CrossRef]
- Baron, C.S.; van Oudenaarden, A. Unravelling Cellular Relationships during Development and Regeneration Using Genetic Lineage Tracing. Nat. Rev. Mol. Cell Biol. 2019, 20, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.R.; Burger, B.; Kühl, C.M.C.; Candreva, T.; dos Anjos, M.B.P.; Rodrigues, H.G. Wound Healing and Omega-6 Fatty Acids: From Inflammation to Repair. Mediat. Inflamm. 2018, 2018, 2503950. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; McGeachy, M.J. IL-17 in the Pathogenesis of Disease: Good Intentions Gone Awry. Annu. Rev. Immunol. 2021, 39, 537–556. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, M. The Role of Th17-Related Cytokines in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 1314. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, X.; Herjan, T.; Li, X. The Role of Interleukin-17 in Tumor Development and Progression. J. Exp. Med. 2020, 217, e20190297. [Google Scholar] [CrossRef] [PubMed]
- Modarresi Chahardehi, A.; Arsad, H.; Lim, V. Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants. Plants 2020, 9, 1345. [Google Scholar] [CrossRef] [PubMed]
- García, N.; Zazueta, C.; Aguilera-Aguirre, L. Oxidative Stress and Inflammation in Cardiovascular Disease. Oxidative Med. Cell. Longev. 2017, 2017, 5853238. [Google Scholar] [CrossRef] [PubMed]
- Ambade, A.; Mandrekar, P. Oxidative Stress and Inflammation: Essential Partners in Alcoholic Liver Disease. Int. J. Hepatol. 2012, 2012, 853175. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free. Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Barbosa, J.D.S.; Costa, M.S.S.P.; Melo, L.F.M.D.; Medeiros, M.J.C.D.; Pontes, D.D.L.; Scortecci, K.C.; Rocha, H.A.O. Caulerpa cupressoides Var. Flabellata. Mar. Drugs 2019, 17, 105. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of Nitrate, Nitrite, and [15N]Nitrate in Biological Fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.-R. Eleven Golden Rules of Quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef] [PubMed]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-Free Analysis of Quantitative Real-Time Polymerase Chain Reaction (PCR) Data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Pant, B.D.; Buhtz, A.; Kehr, J.; Scheible, W.-R. MicroRNA399 Is a Long-Distance Signal for the Regulation of Plant Phosphate Homeostasis. Plant J. 2008, 53, 731–738. [Google Scholar] [CrossRef]
- Balekar, N.; Katkam, N.G.; Nakpheng, T.; Jehtae, K.; Srichana, T. Evaluation of the Wound Healing Potential of Wedelia trilobata (L.) Leaves. J. Ethnopharmacol. 2012, 141, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Tonin, T.D.; Thiesen, L.C.; de Oliveira Nunes, M.L.; Broering, M.F.; Donato, M.P.; Goss, M.J.; Petreanu, M.; Niero, R.; Machado, I.D.; Santin, J.R. Rubus imperialis (Rosaceae) Extract and Pure Compound Niga-Ichigoside F1: Wound Healing and Anti-Inflammatory Effects. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2016, 389, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative Properties of Xanthan on the Autoxidation of Soybean Oil in Cyclodextrin Emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant Activity of Sulfated Polysaccharide Fractions Extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, N.; De, B. Antioxidant Activity of Piper betle L. Leaf Extract in vitro. Food Chem. 2004, 88, 219–224. [Google Scholar] [CrossRef]
- Melo-Silveira, R.F.; Fidelis, G.P.; Costa, M.S.S.P.; Telles, C.B.S.; Dantas-Santos, N.; de Oliveira Elias, S.; Ribeiro, V.B.; Barth, A.L.; Macedo, A.J.; Leite, E.L.; et al. In Vitro Antioxidant, Anticoagulant and Antimicrobial Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted from Corn Cobs. Int. J. Mol. Sci. 2011, 13, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-Y.; Kim, E.-A.; Lee, J.-H.; Kang, M.-C.; Lee, J.-S.; Kim, J.-S.; Jung, W.-K.; Jeon, Y.-J. Protective Effect of Aquacultured Flounder Fish-Derived Peptide against Oxidative Stress in Zebrafish. Fish Shellfish. Immunol. 2014, 36, 320–323. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
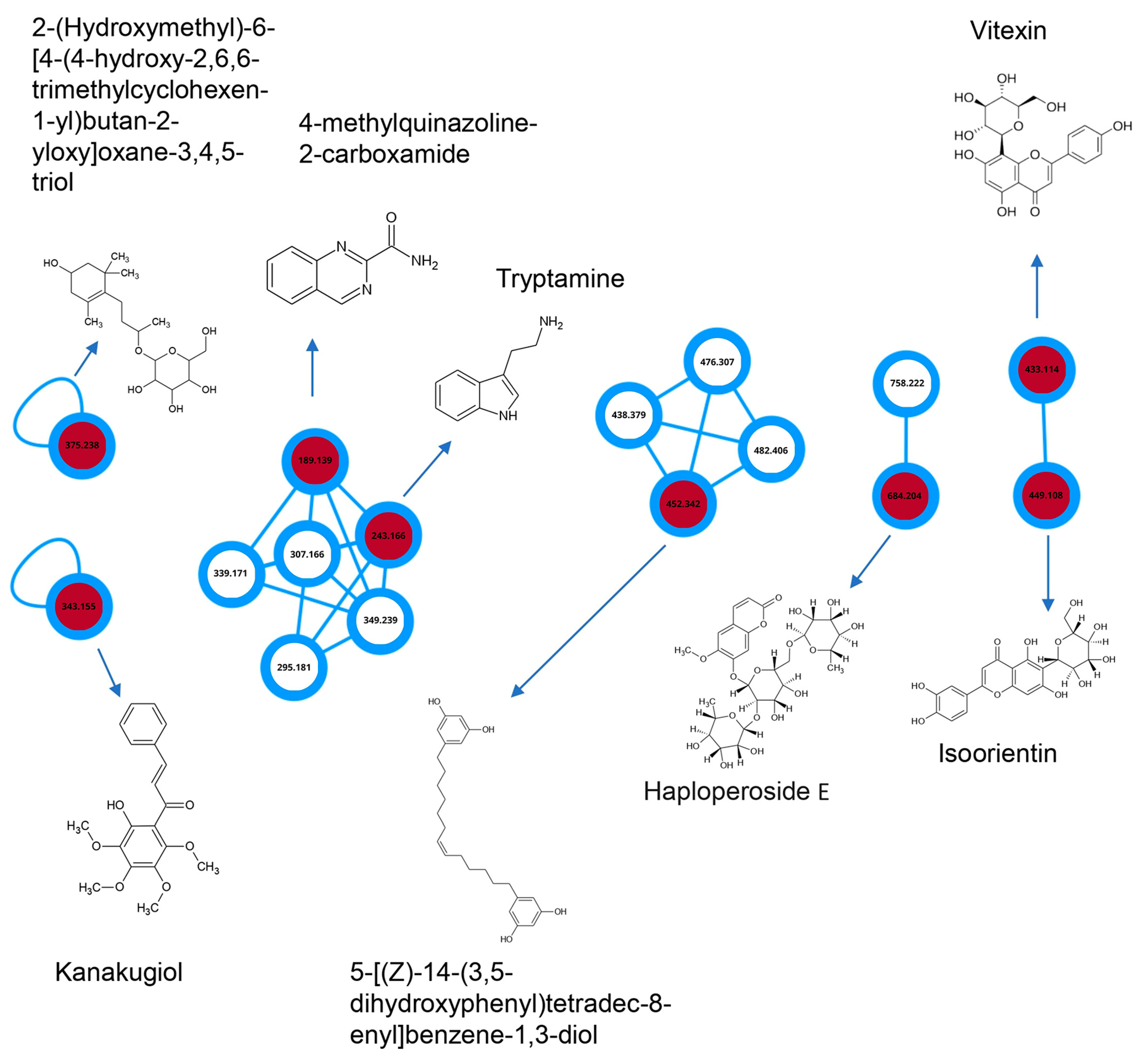{kind=link}
| Retention Time (min.) | Λ | Phenolic Compound by Library Comparison | AF | BF |
|---|---|---|---|---|
| 2.000 | 280 | galic acid | X | ND |
| 10.257 | 280 | p-cumaric acid | X | ND |
| 13.576 | 280/352 | vitexin | X | X |
| t.r. (s) | Molecules | Mass [M+H]+ | Mass Difference | Class |
|---|---|---|---|---|
| 1218.59 | Isoorientin | 449.11 | 0 | Flavonoid/Flavone |
| 1440.46 | 2-(hydroxymethyl)-6-[4-(4-hydroxy-2,6,6-trimethylcyclohexen-1-yl)butan-2-yloxy]oxane-3,4,5-triol | 375.24 | 0 | |
| 1334.56 | Vitexin | 433.11 | 0 | Flavonoid/Flavone |
| 2655.09 | Haploperodide E | 684.20 | 0.97 | cumarin |
| 2199.66 | Kanakugiol | 343.15 | 1.98 | Chalcone |
| 826.32 | 4-methylquinazoline-2-carboxamide | 189.14 | 1.06 | Alkaloid |
| 2185.62 | 5-[(Z)-14-(3,5-dihydroxyphenyl)tetradec-8-enyl]benzene-1,3-diol | 452.34 | 1.12 | Polikedy |
| 1497.61 | tryptamine | 243.19 | 1.98 | Alkaloid |
| Primer | Sequence |
|---|---|
| Elongation Factor (EF) | Reverse–5′ GGG TCA GAT TTC TTG AT′G GG 3′ |
| Forward–5′ CTG THE AG CGG CTG GCC′ AAG 3′ | |
| β-actin | Reverse–5′ CCT AGA AGC ATT TGC GGT GCA GAG 3′ |
| Forward–5′ TCA TGA AGT GTG CGT TGA CAT ′CG T 3′ | |
| Nitric Oxide Synthase (iNOS) | Reverse–5′ GTG CTT GTC ACC ACC AGC AGT AGT 3′ |
| Forward–5′ ACC TTG TTC AGC TCA GCC TTC AAC 3′ | |
| Interleukin 10 (IL-10) | Reverse–5′ TGG CAT GCT CAT TCA GCT CTT ATC 3′ |
| Forward–5′TAT GCA AGG ATT TGG TGT TGT TGG 3′; |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Melo, L.F.M.; Barbosa, J.d.S.; Cordeiro, M.L.d.S.; Aquino-Martins, V.G.d.Q.; Silva, A.P.d.; Paiva, W.d.S.; Silveira, E.R.; dos Santos, D.Y.A.C.; Rocha, H.A.O.; Scortecci, K.C. The Antioxidant and Immunomodulatory Potential of Coccoloba alnifolia Leaf Extracts. Int. J. Mol. Sci. 2023, 24, 15885. https://doi.org/10.3390/ijms242115885
de Melo LFM, Barbosa JdS, Cordeiro MLdS, Aquino-Martins VGdQ, Silva APd, Paiva WdS, Silveira ER, dos Santos DYAC, Rocha HAO, Scortecci KC. The Antioxidant and Immunomodulatory Potential of Coccoloba alnifolia Leaf Extracts. International Journal of Molecular Sciences. 2023; 24(21):15885. https://doi.org/10.3390/ijms242115885
Chicago/Turabian Stylede Melo, Luciana Fentanes Moura, Jefferson da Silva Barbosa, Maria Lúcia da Silva Cordeiro, Verônica Giuliani de Queiroz Aquino-Martins, Ariana Pereira da Silva, Weslley de Souza Paiva, Elielson Rodrigo Silveira, Déborah Yara A. Cursino dos Santos, Hugo Alexandre Oliveira Rocha, and Kátia Castanho Scortecci. 2023. "The Antioxidant and Immunomodulatory Potential of Coccoloba alnifolia Leaf Extracts" International Journal of Molecular Sciences 24, no. 21: 15885. https://doi.org/10.3390/ijms242115885
APA Stylede Melo, L. F. M., Barbosa, J. d. S., Cordeiro, M. L. d. S., Aquino-Martins, V. G. d. Q., Silva, A. P. d., Paiva, W. d. S., Silveira, E. R., dos Santos, D. Y. A. C., Rocha, H. A. O., & Scortecci, K. C. (2023). The Antioxidant and Immunomodulatory Potential of Coccoloba alnifolia Leaf Extracts. International Journal of Molecular Sciences, 24(21), 15885. https://doi.org/10.3390/ijms242115885