
Ultrasensitive Quantification of Crustacean Tropomyosin by Immuno-PCR

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
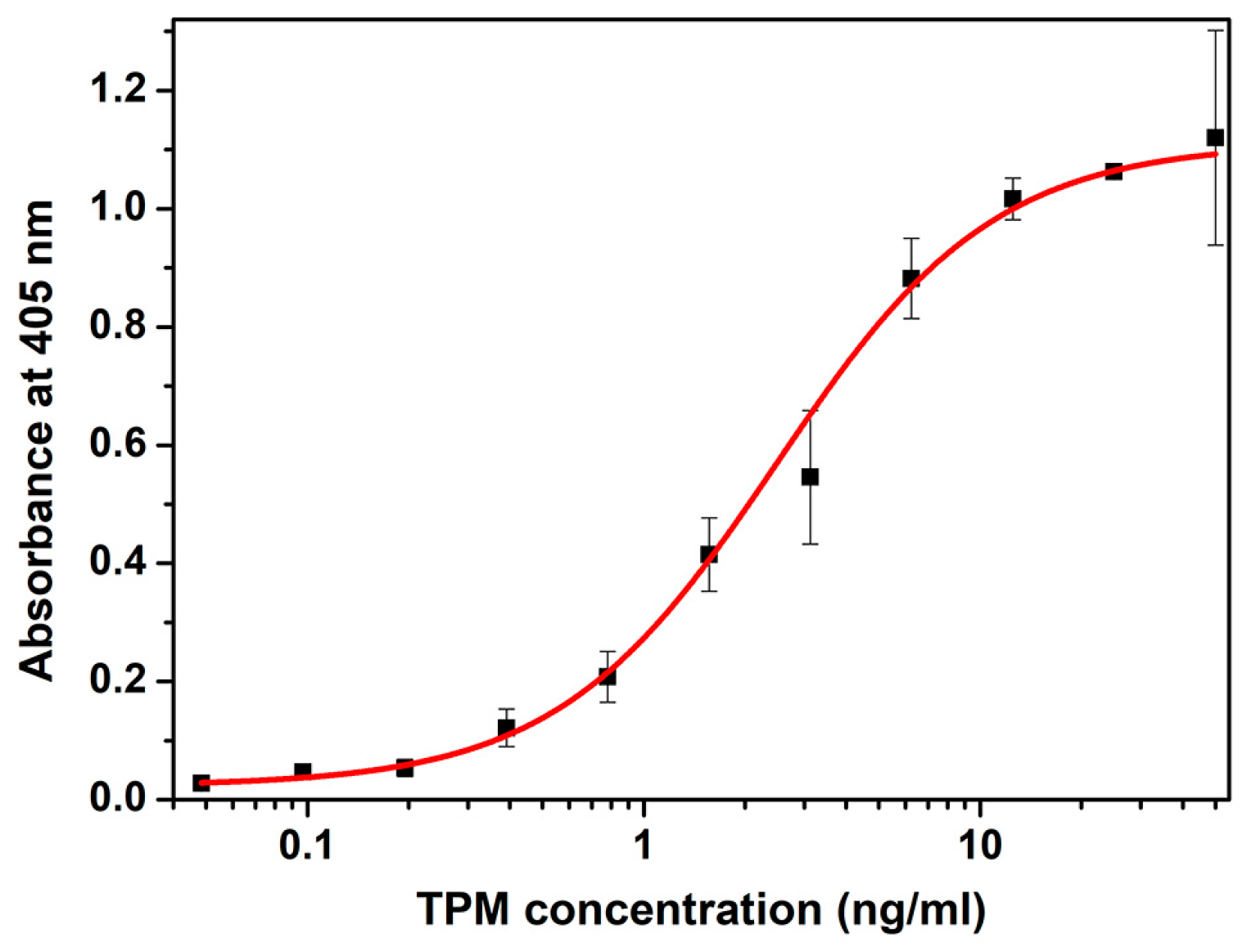
2.1. Development of Sandwich ELISA for Crustacean Tropomyosin Quantification
2.2. Antibody–DNA Conjugate Preparation
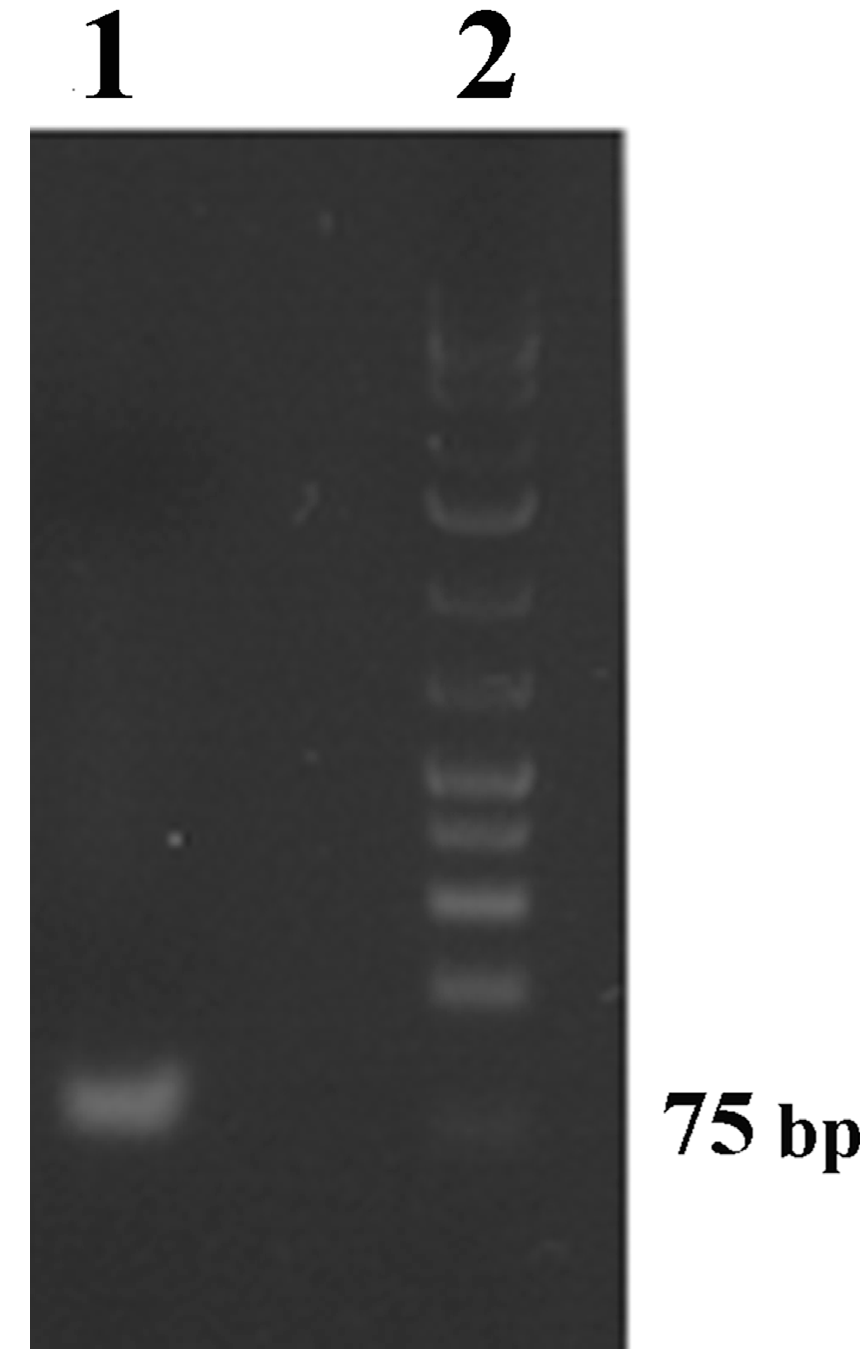
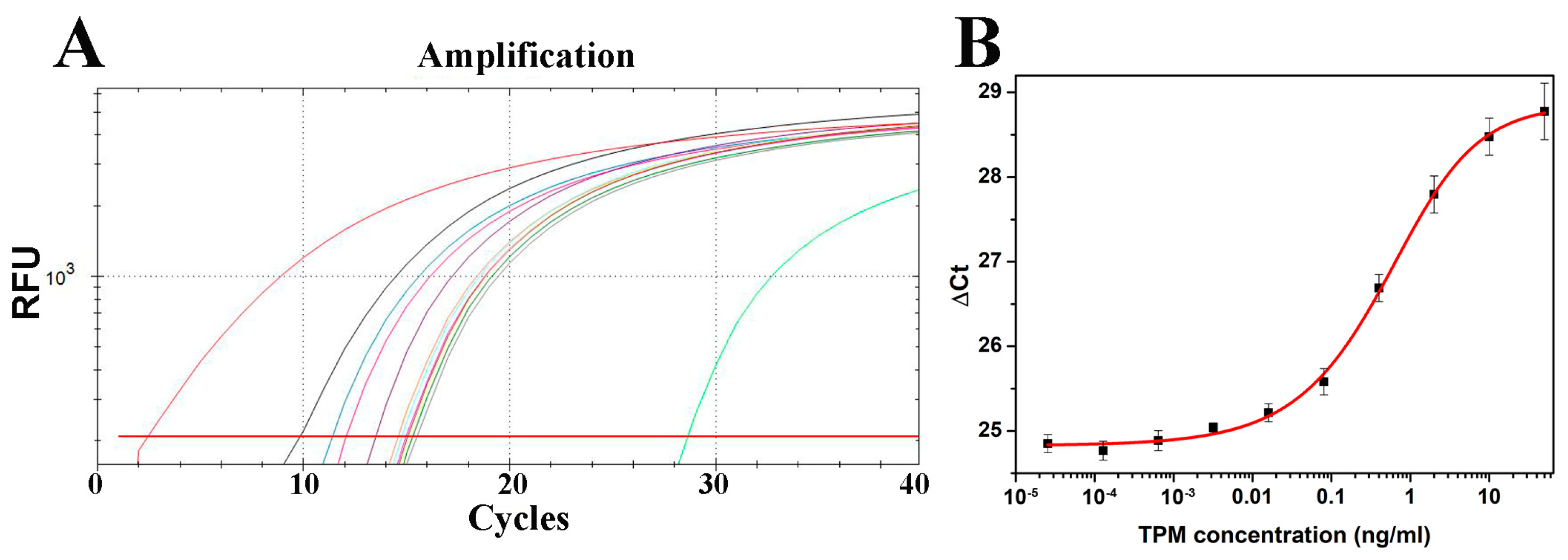
2.3. Development of Immuno-PCR for Crustacean Tropomyosin Quantification
2.4. Method Validation
2.4.1. Sensitivity and Linear Range
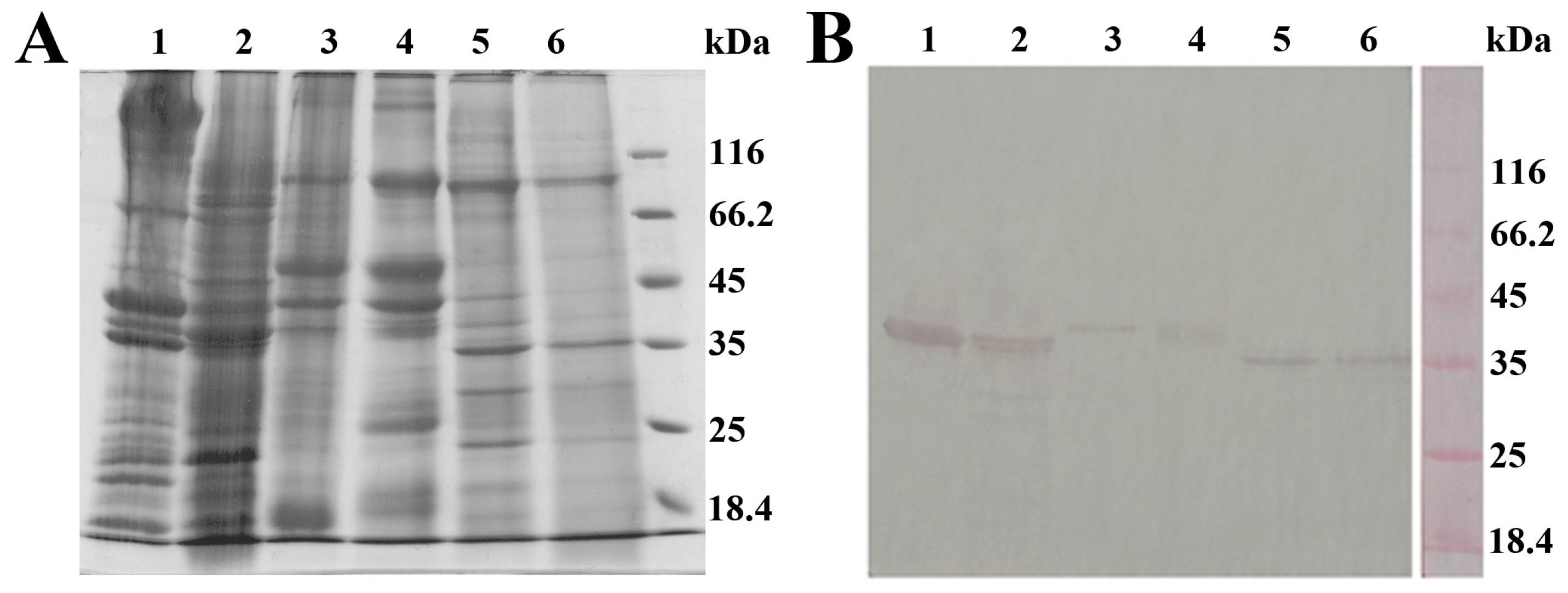
2.4.2. Specificity
2.4.3. Precision
2.4.4. Method Accuracy
2.4.5. Food Sample Analysis
3. Materials and Methods
3.1. Materials
3.2. Sandwich ELISA
3.3. DNA Preparation and Covalent Conjugation to Secondary Antibodies
3.4. Sandwich Immuno-PCR
3.5. Food Sample Preparation
3.6. SDS-PAGE and Western Blot Analysis of Shellfish Extracts
3.7. Limit of Detection and Limit of Quantification Determination
3.8. Precision Calculation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fiocchi, A.; Risso, D.; DunnGalvin, A.; González Díaz, S.N.; Monaci, L.; Fierro, V.; Ansotegui, I.J. Food labeling issues for severe food allergic patients. World Allergy Organ. J. 2021, 14, 100598. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, H.; Sun, D. An overview of tropomyosin as an important seafood allergen: Structure, cross-reactivity, epitopes, allergenicity, and processing modifications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Moonesinghe, H.; Mackenzie, H.; Venter, C.; Kilburn, S.; Turner, P.; Weir, K.; Dean, T. Prevalence of fish and shellfish allergy. Ann. Allergy Asthma Immunol. 2016, 117, 264–272.e4. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Johnston, E.B.; Nugraha, R.; Le, T.T.K.; Kalic, T.; McLean, T.R.; Kamath, S.D.; Lopata, A.L. Seafood allergy: A comprehensive review of fish and shellfish allergens. Mol. Immunol. 2018, 100, 28–57. [Google Scholar] [CrossRef]
- Behrmann, E.; Müller, M.; Penczek, P.A.; Mannherz, H.G.; Manstein, D.J.; Raunser, S. Structure of the rigor actin-tropomyosin-myosin complex. Cell 2012, 150, 327–338. [Google Scholar] [CrossRef]
- Kamath, S.D.; Rahman, A.M.A.; Voskamp, A.; Komoda, T.; Rolland, J.M.; O’Hehir, R.E.; Lopata, A.L. Effect of heat processing on antibody reactivity to allergen variants and fragments of black tiger prawn: A comprehensive allergenomic approach. Mol. Nutr. Food Res. 2014, 58, 1144–1155. [Google Scholar] [CrossRef]
- Zeng, L.; Song, S.; Zheng, Q.; Luo, P.; Wu, X.; Kuang, H. Development of a sandwich ELISA and immunochromatographic strip for the detection of shrimp tropomyosin. Food Agric. Immunol. 2019, 30, 606–619. [Google Scholar] [CrossRef]
- Fuller, H.R.; Goodwin, P.R.; Morris, G.E. An enzyme-linked immunosorbent assay (ELISA) for the major crustacean allergen, tropomyosin, in food. Food Agric. Immunol. 2006, 17, 43–52. [Google Scholar] [CrossRef]
- Werner, M.T.; Fæste, C.K.; Egaas, E. Quantitative Sandwich ELISA for the Determination of Tropomyosin from Crustaceans in Foods. J. Agric. Food Chem. 2007, 55, 8025–8032. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, Y.; Li, Z.; Pramod, S.N.; Zhang, L.; Lin, H. Development of ELISA Method for Detecting Crustacean Major Allergen Tropomyosin in Processed Food Samples. Food Anal. Methods 2019, 12, 2719–2729. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y.; Li, R.; Timira, V.; Dasanayaka, B.P.; Zhang, Z.; Zhang, J.; Lin, H.; Li, Z. Evaluation of poly- and monoclonal antibody-based sandwich enzyme-linked immunosorbent assay (ELISA) for their performance to detect crustacean residues in processed foods. Food Control 2022, 138, 108983. [Google Scholar] [CrossRef]
- Seiki, K.; Oda, H.; Yoshioka, H.; Sakai, S.; Urisu, A.; Akiyama, H.; Ohno, Y. A Reliable and Sensitive Immunoassay for the Determination of Crustacean Protein in Processed Foods. J. Agric. Food Chem. 2007, 55, 9345–9350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lu, Y.; Ushio, H.; Shiomi, K. Development of sandwich ELISA for detection and quantification of invertebrate major allergen tropomyosin by a monoclonal antibody. Food Chem. 2014, 150, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-Y.; Lee, C.-H.; Huang, E.S.; Sheu, S.-C.; Yu, H.-S. Quantification of Crustacean Tropomyosin, a Major Food Allergen, in Eight Species of Taiwanese Shrimp Based on Immunoassay. Food Anal. Methods 2018, 11, 2607–2613. [Google Scholar] [CrossRef]
- Angulo-Ibáñez, A.; Eletxigerra, U.; Lasheras, X.; Campuzano, S.; Merino, S. Electrochemical tropomyosin allergen immunosensor for complex food matrix analysis. Anal. Chim. Acta 2019, 1079, 94–102. [Google Scholar] [CrossRef]
- Wang, Y.; Rao, Z.; Zhou, J.; Zheng, L.; Fu, L. A chiral assembly of gold nanoparticle trimer-based biosensors for ultrasensitive detection of the major allergen tropomyosin in shellfish. Biosens. Bioelectron. 2019, 132, 84–89. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, Y.; Qian, Y.; Zhang, T.; Zheng, L.; Fu, L. Quantification of shellfish major allergen tropomyosin by SPR biosensor with gold patterned Biochips. Food Control 2020, 107, 106547. [Google Scholar] [CrossRef]
- Torre, R.; Freitas, M.; Costa-Rama, E.; Nouws, H.P.A.; Delerue-Matos, C. Food allergen control: Tropomyosin analysis through electrochemical immunosensing. Food Chem. 2022, 396, 133659. [Google Scholar] [CrossRef]
- Amouzadeh Tabrizi, M.; Shamsipur, M.; Saber, R.; Sarkar, S.; Ebrahimi, V. A high sensitive visible light-driven photoelectrochemical aptasensor for shrimp allergen tropomyosin detection using graphitic carbon nitride-TiO2 nanocomposite. Biosens. Bioelectron. 2017, 98, 113–118. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Wei, X.; Zhang, J.; Mo, S. DNA aptamer for use in a fluorescent assay for the shrimp allergen tropomyosin. Microchim. Acta 2017, 184, 633–639. [Google Scholar] [CrossRef]
- Chinnappan, R.; Rahamn, A.A.; AlZabn, R.; Kamath, S.; Lopata, A.L.; Abu-Salah, K.M.; Zourob, M. Aptameric biosensor for the sensitive detection of major shrimp allergen, tropomyosin. Food Chem. 2020, 314, 126133. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Li, H.; Peng, Y.; Fu, L. A fluorometric sandwich biosensor based on rationally imprinted magnetic particles and aptamer modified carbon dots for the detection of tropomyosin in seafood products. Food Control 2022, 132, 108552. [Google Scholar] [CrossRef]
- Jiang, D.; Ji, J.; An, L.; Sun, X.; Zhang, Y.; Zhang, G.; Tang, L. Mast cell-based electrochemical biosensor for quantification of the major shrimp allergen Pen a 1 (tropomyosin). Biosens. Bioelectron. 2013, 50, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhu, P.; Jiang, H.; Ji, J.; Sun, X.; Gu, W.; Zhang, G. Fluorescent magnetic bead-based mast cell biosensor for electrochemical detection of allergens in foodstuffs. Biosens. Bioelectron. 2015, 70, 482–490. [Google Scholar] [CrossRef]
- Shi, L.; Wang, X.C.; Liu, Y.; Lu, Y. Rapid Detection of Shellfish Major Allergen Tropomyosin Using Superparamagnetic Nanoparticle-Based Lateral Flow Immunoassay. Adv. Mater. Res. 2011, 311–313, 436–445. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Lin, H.; Siddanakoppalu, P.N.; Zhou, J.; Chen, G.; Yu, Z. Quantum-dot-based lateral flow immunoassay for the rapid detection of crustacean major allergen tropomyosin. Food Control 2019, 106, 106714. [Google Scholar] [CrossRef]
- Li, R.; Zhang, Y.; Zhao, J.; Wang, Y.; Wang, H.; Zhang, Z.; Lin, H.; Li, Z. Quantum-dot-based sandwich lateral flow immunoassay for the rapid detection of shrimp major allergen tropomyosin. J. Food Compos. Anal. 2022, 114, 104776. [Google Scholar] [CrossRef]
- Coulomb, B.; Robert-Peillard, F.; Ben Ali Gam, N.; Sadok, S.; Boudenne, J.-L. Multisyringe Flow Injection Analysis of Tropomyosin Allergens in Shellfish Samples. Molecules 2021, 26, 5809. [Google Scholar] [CrossRef]
- Korte, R.; Monneuse, J.-M.; Gemrot, E.; Metton, I.; Humpf, H.-U.; Brockmeyer, J. New High-Performance Liquid Chromatography Coupled Mass Spectrometry Method for the Detection of Lobster and Shrimp Allergens in Food Samples via Multiple Reaction Monitoring and Multiple Reaction Monitoring Cubed. J. Agric. Food Chem. 2016, 64, 6219–6227. [Google Scholar] [CrossRef]
- Ho, C.-W.; Hsu, J.-L.; Chen, S.-H.; Liaw, E.-T.; Liu, S.-S.; Huang, E.S.; Chen, Y.-K.; Jean Huang, C.-C.; Yu, H.-S. Development and validation of mass spectrometry-based method for detecting shrimp allergen tropomyosin. LWT 2021, 152, 112367. [Google Scholar] [CrossRef]
- Wang, J.; Ge, M.; Sun, L.; Ahmed, I.; Li, W.; Lin, H.; Lin, H.; Li, Z. Quantification of crustacean tropomyosin in foods using high-performance liquid chromatography–tandem mass spectrometry method. J. Sci. Food Agric. 2021, 101, 5278–5285. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Ma, J.; Li, C.; Wang, Y.; Zeng, W.; Li, Q.; Zhou, J.; Wang, L.; Wang, Y.; Zhang, Y. Determination of Tropomyosin in Shrimp and Crab by Liquid Chromatography–Tandem Mass Spectrometry Based on Immunoaffinity Purification. Front. Nutr. 2022, 9, 848294. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, T.; Wang, Y.; Zhang, S.; Sheng, H.; Fu, L. Liquid chromatography coupled to tandem mass spectrometry for comprehensive quantification of crustacean tropomyosin and arginine kinase in food matrix. Food Control 2022, 140, 109137. [Google Scholar] [CrossRef]
- Kim, M.-J.; Kim, H.-I.; Kim, J.-H.; Suh, S.-M.; Kim, H.-Y. Rapid on-site detection of shrimp allergen tropomyosin using a novel ultrafast PCR system. Food Sci. Biotechnol. 2019, 28, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Smith, C.L.; Cantor, C.R. Immuno-PCR: Very Sensitive Antigen Detection by Means of Specific Antibody-DNA Conjugates. Science 1992, 258, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Rajkovic, A.; El Moualij, B.; Fikri, Y.; Dierick, K.; Zorzi, W.; Heinen, E.; Uner, A.; Uyttendaele, M. Detection of Clostridium botulinum neurotoxins A and B in milk by ELISA and immuno-PCR at higher sensitivity than mouse bio-assay. Food Anal. Methods 2012, 5, 319–326. [Google Scholar] [CrossRef]
- Rajkovic, A.; El Moualij, B.; Uyttendaele, M.; Brolet, P.; Zorzi, W.; Heinen, E.; Foubert, E.; Debevere, J. Immunoquantitative Real-Time PCR for Detection and Quantification of Staphylococcus aureus Enterotoxin B in Foods. Appl. Environ. Microbiol. 2006, 72, 6593–6599. [Google Scholar] [CrossRef]
- Monjezi, R.; Tan, S.W.; Tey, B.T.; Sieo, C.C.; Tan, W.S. Detection of hepatitis B virus core antigen by phage display mediated TaqMan real-time immuno-PCR. J. Virol. Methods 2013, 187, 121–126. [Google Scholar] [CrossRef]
- Johnston, E.B.; Kamath, S.D.; Lopata, A.L.; Schaeffer, P.M. Tus- Ter -lock immuno-PCR assays for the sensitive detection of tropomyosin-specific IgE antibodies. Bioanalysis 2014, 6, 465–476. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPM (ng/mL) | RSD 1 (%) | RSD 2 (%) | RSD 3 (%) | Inter-Day RSD (%) |
|---|---|---|---|---|
| 50 | 1.2 | 1.3 | 1.7 | 2 |
| 10 | 0.8 | 0.2 | 0.5 | 1.8 |
| 2 | 0.8 | 3 | 1.9 | 1.6 |
| 0.4 | 0.6 | 3.8 | 0.8 | 1.7 |
| 0.08 | 0.6 | 0.3 | 0.6 | 2.2 |
| 0.016 | 0.4 | 0.2 | 0 | 2.1 |
| 0.0032 | 0.2 | 0.4 | 0.4 | 2.2 |
| 0.00064 | 0.5 | 0.6 | 0.1 | 1.5 |
| 0.000128 | 0.4 | 0.9 | 0.7 | 1.8 |
| 0.0000256 | 0.4 | 0 | 0.3 | 1.7 |
| Recovery of the Method | |||
|---|---|---|---|
| Spike Level | Mean ± SD (ng/mL) | Recovery (%) | RSD (%) |
| 10 LOQ—0.2 ng/mL | 0.22 ± 0.04 | 108.7 | 19.3 |
| 50 LOQ—1 ng/mL | 1.16 ± 0.24 | 115.6 | 24.5 |
| 100 LOQ—2 ng/mL | 1.75 ± 0.10 | 87.7 | 5 |
| Quantification of crustacean tropomyosin | |||
| Sample | TPM (ng/g) | Declared by manufacturer | |
| Octopus in tomato sauce | 135 ± 16 | yes | |
| Spicy noodles | 116 ± 46 | yes | |
| Mild noodles | ND | yes | |
| Fish sticks | ND | no | |
| Tuna pate | ND | no | |
| LOD | LOQ | Method | Reference |
|---|---|---|---|
| 0.4 ng/mL | 0.6 ng/mL | ELISA, c: mAb, d: pAb | [11] |
| 0.8 ng/mL | 1 ng/mL | ELISA, c: pAb, d: pAb | [11] |
| 6.8 ng/mL | 13.67 ng/mL | ELISA, c: pAb, d:pAb | [10] |
| 30 pg/mL | 64 pg/mL | ELISA, c: mAb, d: pAb | [14] |
| 2 ng/mL | 3 ng/mL | ELISA, c: pAB, d: pAb | [9] |
| 0.71 ng/mL | 2.25 ng/mL | ELISA, c: mAb, d: pAb | [12] |
| 3.58 µg/g | 7.16 µg/g | UPLC-MS/MS after purification of extracts via immunoaffinity column and digestion by trypsin, with isotope-labeled standard | [32] |
| 72 ng/mL | 219 ng/mL | LC-MS/MS after trypsin digestion and SPE, with isotope-labeled standard | [30] |
| 18 ng/mL | 59 ng/mL | UHPLC-MS/MS after trypsin digestion and SPE, with isotope-labeled standard | [33] |
| NA, depends on quality of MS and the removal of impurities | 1.6 µg/g | HPLC–MS/MS, after trypsin digestion and SPE, with isotope-labeled standard | [31] |
| 1.3 µM of Glu (about 0.85 µg/mL TPM *) | 4.4 µM of Glu (about 2.85 µg/mL TPM *) | Fluorimetric determination of Glu after protein hydrolysis, conversion of Glu to pyroglutamic acid and its derivatization | [28] |
| 0.47 ng/mL | 1.6 ng/mL | Electrochemical Immunosensing, c: mAb, d: pAb | [18] |
| 1 µg/mL | 2.5 µg/mL | SPR biosensor, c: mAb | [17] |
| 21 pg/mL | 70 pg/mL | A biosensor based on a chiral assembly of polymer of gold nanoparticle (AuNP) trimers, c: mAb | [16] |
| 30.76 ng/mL | 102.53 ng/mL | Sandwich biosensor based on imprinted magnetic particles and aptamer-modified carbon dots | [22] |
| 0.15 µg/mL | 0.5 µg/mL | DNA aptamer assay | [20] |
| 230 pg/mL | 1 ng/mL | Visible light-driven photoelectrochemical aptasensor | [19] |
| 0.15 µg/mL | 0.5 µg/mL | Mast cell-based electrochemical biosensor | [23] |
| 30 ng/mL | 100 ng/mL | Fluorescent magnetic bead-based mast cell biosensor with electrochemical detection | [24] |
| 0.75 ng/mL | NA | ELISA, c: pAb, d: pAb | [8] |
| 0.45 ng/mL | NA | ELISA, c: mAb, d: mAb | [7] |
| 90 pg/mL | NA | ELISA, c: mAb, d: pAb | [13] |
| 25 µg/g | NA | LC-MS after trypsin digestion and SPE, label-free quantification | [29] |
| Visual 50 ng/mL Instrumental 10 ng/mL | NA | Quantum-dot-based sandwich lateral flow immunoassay, c: pAb | [27] |
| 15.6 ng/mL | NA | Superparamagnetic nanoparticle-based lateral flow immunoassay, c: mAb | [25] |
| Visual 500 ng/mL Instrumental 50 ng/mL | NA | Quantum-dot-based lateral flow immunoassay, c: pAb | [26] |
| 0.5 ng/mL | NA | Immunochromatographic assay strip, c: mAb, d: mAb | [7] |
| 2 nM (75 ng/mL) | NA | Aptameric biosensor | [21] |
| 47 pg/mL | NA | Amperometric immunosensor based on a sandwich immunoassay, c; mAb, d: pAb | [15] |
| 0.32 ng/mL TPM DNA | NA | Ultrafast PCR | [34] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radomirović, M.; Gligorijević, N.; Stanić-Vučinić, D.; Rajković, A.; Ćirković Veličković, T. Ultrasensitive Quantification of Crustacean Tropomyosin by Immuno-PCR. Int. J. Mol. Sci. 2023, 24, 15410. https://doi.org/10.3390/ijms242015410
Radomirović M, Gligorijević N, Stanić-Vučinić D, Rajković A, Ćirković Veličković T. Ultrasensitive Quantification of Crustacean Tropomyosin by Immuno-PCR. International Journal of Molecular Sciences. 2023; 24(20):15410. https://doi.org/10.3390/ijms242015410
Chicago/Turabian StyleRadomirović, Mirjana, Nikola Gligorijević, Dragana Stanić-Vučinić, Andreja Rajković, and Tanja Ćirković Veličković. 2023. "Ultrasensitive Quantification of Crustacean Tropomyosin by Immuno-PCR" International Journal of Molecular Sciences 24, no. 20: 15410. https://doi.org/10.3390/ijms242015410
APA StyleRadomirović, M., Gligorijević, N., Stanić-Vučinić, D., Rajković, A., & Ćirković Veličković, T. (2023). Ultrasensitive Quantification of Crustacean Tropomyosin by Immuno-PCR. International Journal of Molecular Sciences, 24(20), 15410. https://doi.org/10.3390/ijms242015410