
Fermentation of Murta (Ugni molinae) Juice: Effect on Antioxidant Activity and Control of Enzymes Associated with Glucose Assimilation


Abstract
:1. Introduction
2. Results and Discussion
2.1. Fermented Fruit Juices Characteristics
2.2. Total Phenolic Content (TPC) and Antioxidant Activity
2.2.1. Total Polyphenol Content (TPC)
2.2.2. DPPH Free Radical Scavenging Method
2.2.3. Oxygen Radical Absorbance Capacity (ORAC)
2.3. Enzyme Inhibition Activities
2.3.1. Inhibitory Activity of α-Amylase Enzyme
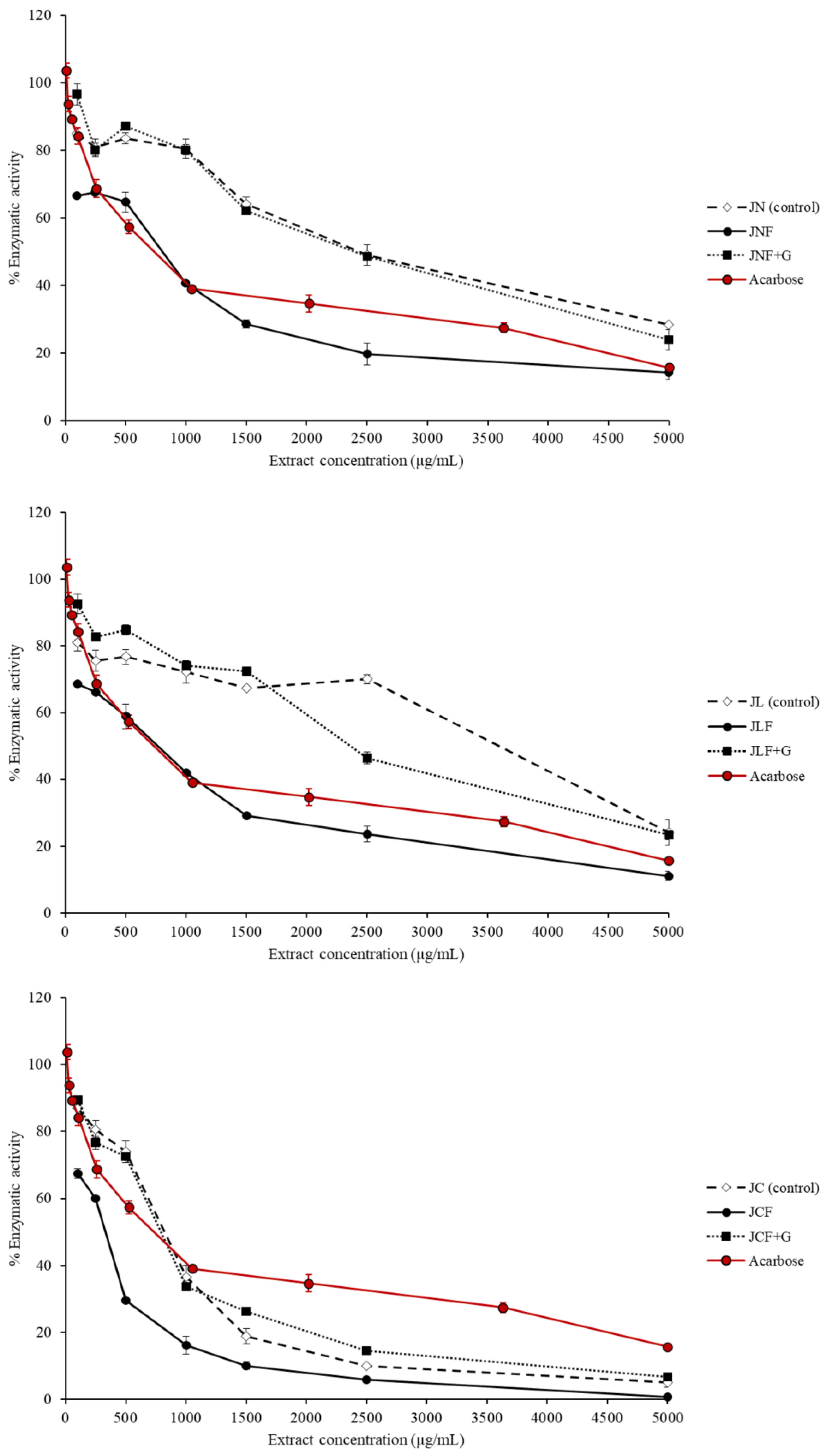
2.3.2. Inhibitory Activity of α-Glucosidase
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Juices Preparation
3.2.1. Natural Juice
3.2.2. Commercial Juice
3.2.3. Freeze-Dried Juice
3.3. Juices Fermentation
3.4. Total Polyphenols Content (TPC)
3.5. Antioxidant Activity
3.5.1. DPPH Radical Scavenging Activity
3.5.2. Oxygen Radical Absorbance Capacity (ORAC)
3.6. Inhibition of Carbohydrate Hydrolytic Enzymes
3.6.1. Pretreatment of Samples
3.6.2. Anti α-Amylase Activity
3.6.3. Anti α-Glucosidase Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ali, A.; Rahut, D.B. Healthy foods as proxy for functional foods: Consumers’ awareness, perception, and demand for natural functional foods in Pakistan. Int. J. Food Sci. 2019, 2019, 6390650. [Google Scholar] [CrossRef] [PubMed]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.G.R.; Gasga, V.M.Z.; Pescuma, M.; Van Nieuwenhove, C.; Mozzi, F.; Burgos, J.A.S. Fruits and fruit by-products as sources of bioactive compounds. Benefits and trends of lactic acid fermentation in the development of novel fruit-based functional beverages. Food Res. Int. 2021, 140, 109854. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Li, T.; Qi, J.; Jiang, T.; Xu, H.; Lei, H. Effects of lactic acid fermentation-based biotransformation on phenolic profiles, antioxidant capacity and flavor volatiles of apple juice. LWT 2020, 122, 109064. [Google Scholar] [CrossRef]
- Sun, C.; Liu, Y.; Zhang, L.; Rayat, G.R.; Xiao, J.; Jiang, H.; Li, X.; Chen, K. Anti-diabetic effects of natural antioxidants from fruits. Trends Food Sci. Technol. 2021, 117, 3–14. [Google Scholar] [CrossRef]
- Edirisinghe, I.; Burton-Freeman, B. Anti-diabetic actions of Berry polyphenols—Review on proposed mechanisms of action. J. Berry Res. 2016, 6, 237–250. [Google Scholar] [CrossRef]
- Gayathry, K.S.; John, J.A. Functional beverages: Special focus on anti-diabetic potential. J. Food Process. Preserv. 2021, 45, e15974. [Google Scholar] [CrossRef]
- Pap, N.; Fidelis, M.; Azevedo, L.; Carmo, M.A.V.D.; Wang, D.; Mocan, A.; Pereira, E.P.R.; Xavier-Santos, D.; Sant’Ana, A.S.; Yang, B.; et al. Berry polyphenols and human health: Evidence of antioxidant, anti-inflammatory, microbiota modulation, and cell-protecting effects. Curr. Opin. Food Sci. 2021, 42, 167–186. [Google Scholar] [CrossRef]
- Sivapragasam, N.; Neelakandan, N.; Rupasinghe, H.P.V. Potential health benefits of fermented blueberry: A review of current scientific evidence. Trends Food Sci. Technol. 2023, 132, 103–120. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, F.; Chai, Z.; Liu, M.; Battino, M.; Meng, X. Mixed fermentation of blueberry pomace with L. rhamnosus GG and L. plantarum-1: Enhance the active ingredient, antioxidant activity and health-promoting benefits. Food Chem. Toxicol. 2019, 131, 110541. [Google Scholar] [CrossRef]
- Fredes, C.; Parada, A.; Salinas, J.; Robert, P. Phytochemicals and Traditional Use of Two Southernmost Chilean Berry Fruits: Murta (Ugni molinae Turcz) and Calafate (Berberis buxifolia Lam.). Foods 2020, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Vega-Galvez, A.; Rodríguez, A.; Stucken, K. Antioxidant, functional properties and health-promoting potential of native South American berries: A review. J. Sci. Food Agric. 2021, 101, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In Vitro and In Vivo Anti-Diabetic Effects of Anthocyanins from Maqui Berry (Aristotelia chilensis). Food Chem. 2012, 131, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Tovar, P.R.; Mariotti-Celis, M.S.; Pérez-Correa, J.R. Maqui (Aristotelia chilensis (Mol.) Stuntz) and murta (Ugni molinae Turcz): Native Chilean sources of polyphenol compounds. Mini-Rev. Org. Chem. 2019, 16, 261–276. [Google Scholar] [CrossRef]
- López, J.; Vera, C.; Bustos, R.; Florez-Mendez, J. Native berries of Chile: A comprehensive review on nutritional aspects, functional properties, and potential health benefits. J. Food Meas. Charact. 2020, 15, 1139–1160. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Erpel, F.; Mariotti-Celis, M.S.; Parada, J.; Pedreschi, F.; Pérez-Correa, J.R. Pressurized Hot Liquid Extraction with 15% v/v Glycerol-Water as An Effective Environment-Friendly Process to Obtain Durvillaea incurvata and Lessonia spicata Phlorotannin Extracts with Antioxidant and Antihyperglycemic Potential. Antioxidants 2021, 10, 1105. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Szutowska, J. Functional properties of lactic acid bacteria in fermented fruit and vegetable juices: A systematic literature review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Blancas-Benítez, F.J.; Sáyago-Ayerdi, S.G. Polyphenols associated with dietary fibers in plant foods: Molecular interactions and bioaccessibility. Curr. Opin. Food Sci. 2017, 13, 84–88. [Google Scholar] [CrossRef]
- Rocchetti, G.; Gregorio, R.P.; Lorenzo, J.M.; Barba, F.J.; Oliveira, P.G.; Prieto, M.A.; Simal-Gandara, J.; Mosele, J.I.; Motilva, M.J.; Tomas, M.; et al. Functional implications of bound phenolic compounds and phenolics–food interaction: A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 811–842. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Y.; Xiang, J.; Wang, C.; Johnson, J.B.; Beta, T. Diverse polyphenol components contribute to antioxidant activity and hypoglycemic potential of mulberry varieties. LWT 2023, 173, 114308. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Vincentini, O.; Cantatore, V.; Cavoski, I.; Gobbetti, M. Fermented Portulaca oleracea L. Juice: A Novel Functional Beverage with Potential Ameliorating Effects on the Intestinal Inflammation and Epithelial Injury. Nutrients 2019, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jang, H. Characteristics of Vegetable Juice Fermented with Lactobacillus plantarum MKHA15 and Leuconostoc mesenteroids MKSR. J. Korean Diet. Assoc. 2019, 25, 281–294. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, M.; Chang, L. Effects of lactic acid bacteria fermentation on the phytochemicals content, taste and aroma of blended edible rose and shiitake beverage. Food Chem. 2023, 405, 134722. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef]
- Ye, M.; Yue, T.; Yuan, Y. Evolution of polyphenols and organic acids during the fermentation of apple cider. J. Sci. Food Agric. 2014, 94, 2951–2957. [Google Scholar] [CrossRef]
- de Dicastillo, C.L.; Bustos, F.; Valenzuela, X.; López-Carballo, G.; Vilariño, J.M.; Galotto, M.J. Chilean berry Ugni molinae Turcz. fruit and leaves extracts with interesting antioxidant, antimicrobial and tyrosinase inhibitory properties. Food Res. Int. 2017, 102, 119–128. [Google Scholar] [CrossRef]
- Rubilar, M.; Jara, C.; Poo, Y.; Acevedo, F.; Gutierrez, C.; Sineiro, J.; Shene, C. Extracts of maqui (Aristotelia chilensis) and murta (Ugni molinae Turcz.): Sources of antioxidant compounds and α-glucosidase/α-amylase inhibitors. J. Agric. Food Chem. 2011, 59, 1630–1637. [Google Scholar] [CrossRef]
- Shene, C.; Reyes, A.K.; Villarroel, M.; Sineiro, J.; Pinelo, M.; Rubilar, M. Plant location and extraction procedure strongly alter the antimicrobial activity of murta extracts. Eur. Food Res. Technol. 2009, 228, 467–475. [Google Scholar] [CrossRef]
- Ah-Hen, K.S.; Mathias-Rettig, K.; Gómez-Pérez, L.S.; Riquelme-Asenjo, G.; Lemus-Mondaca, R.; Muñoz-Fariña, O. Bioaccessibility of bioactive compounds and antioxidant activity in murta (Ugni molinae T.) berries juices. J. Food Meas. Charact. 2018, 12, 602–615. [Google Scholar] [CrossRef]
- Hauser, C.; Peñaloza, A.; Rodríguez, F.; Guarda, A.; Galotto, M.J. Promising antimicrobial and antioxidant extracts of Murta leaves (Ugni molinae Turcz): Shelf-life extension and food safety. Food Packag. Shelf. Life 2014, 1, 77–85. [Google Scholar] [CrossRef]
- Chen, W.; Xie, C.; He, Q.; Sun, J.; Bai, W. Improvement in color expression and antioxidant activity of strawberry juice fermented with lactic acid bacteria: A phenolic-based research. Food Chem. X 2023, 17, 100535. [Google Scholar] [CrossRef] [PubMed]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- López, J.; Vega-Gálvez, A.; Rodríguez, A.; Uribe, E.; Bilbao-Sainz, C. Murta (Ugni molinae Turcz.): A review on chemical composition, functional components and biological activities of leaves and fruits. Chil. J. Agric. Anim. Sci. 2018, 34, 43–56. [Google Scholar] [CrossRef]
- Speisky, H.; López-Alarcón, C.; Gómez, M.; Fuentes, J.; Sandoval-Acuña, C. First web-based database on total phenolics and oxygen radical absorbance capacity (ORAC) of fruits produced and consumed within the south Andes region of South America. J. Agric. Food Chem. 2012, 60, 8851–8859. [Google Scholar] [CrossRef]
- Junqueira-Gonçalves, M.P.; Yáñez, L.; Morales, C.; Navarro, M.; Contreras, R.A.; Zúñiga, G.E. Isolation and Characterization of Phenolic Compounds and Anthocyanins from Murta (Ugni molinae Turcz.) Fruits. Assessment of Antioxidant and Antibacterial Activity. Molecules 2015, 20, 5698–5713. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Sidro, B.; Ullate, M.; Frias, J.; Vidal-Valverde, C. White cabbage fermentation improves ascorbigen content, antioxidant and nitric oxide production inhibitory activity in LPS-induced macrophages. LWT Food Sci. Technol. 2012, 46, 77–83. [Google Scholar] [CrossRef]
- Piekarska-Radzik, L.; Klewicka, E. Mutual influence of polyphenols and Lactobacillus spp. bacteria in food: A review. Eur. Food Res. Technol. 2021, 247, 9–24. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, C.; Guo, Y.; Wang, X.; Meng, Y. Polyphenols in fermented apple juice: Beneficial effects on human health. J. Funct. Foods 2021, 76, 104294. [Google Scholar] [CrossRef]
- Yoon, J.; Kim, M. In Vitro evaluation of antidiabetic, antidementia, and antioxidant activity of Artemisia capillaris fermented by Leuconostoc spp. LWT 2022, 172, 114163. [Google Scholar] [CrossRef]
- Kalaycıoğlu, Z.; Erim, F.B. Total phenolic contents, antioxidant activities, and bioactive ingredients of juices from pomegranate cultivars worldwide. Food Chem. 2017, 221, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; De Magalhães Carrapeiro, M.; Fogliano, V.; Van Ruth, S.M. Effects of geographical origin, varietal and farming system on the chemical composition and functional properties of purple grape juices: A review. Trends Food Sci. Technol. 2016, 52, 31–48. [Google Scholar] [CrossRef]
- Gaur, G.; Gänzle, M.G. Conversion of (poly)phenolic compounds in food fermentations by lactic acid bacteria: Novel insights into metabolic pathways and functional metabolites. Curr. Res. Food Sci. 2023, 6, 100448. [Google Scholar] [CrossRef]
- Yang, F.; Chen, C.; Ni, D.; Yang, Y.; Tian, J.; Li, Y.; Chen, S.; Ye, X.; Wang, L. Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review. Foods 2023, 12, 3315. [Google Scholar] [CrossRef]
- Suthanthangjai, W.; Davies, K.; Phillips, A.; Ansell, J.; Kilmartin, P. Biotransformation of blueberry anthocyanins to bioavailable phenolic compounds by Lactobacillus. Planta Med. 2013, 79, PJ46. [Google Scholar] [CrossRef]
- Gao, B.; Wang, J.; Wang, Y.; Xu, Z.; Li, B.; Meng, X.; Sun, X.; Zhu, J. Influence of fermentation by lactic acid bacteria and in vitro digestion on the biotransformations of blueberry juice phenolics. Food Control 2022, 133, 108603. [Google Scholar] [CrossRef]
- Wu, Y.; Li, S.; Tao, Y.; Li, D.; Han, Y.; Show, P.L.; Wen, G.; Zhou, J. Fermentation of blueberry and blackberry juices using Lactobacillus plantarum, Streptococcus thermophilus and Bifidobacterium bifidum: Growth of probiotics, metabolism of phenolics, antioxidant capacity in vitro and sensory evaluation. Food Chem. 2021, 348, 129083. [Google Scholar] [CrossRef]
- Cheng, J.R.; Liu, X.M.; Chen, Z.Y.; Zhang, Y.S.; Zhang, Y.H. Mulberry anthocyanin biotransformation by intestinal probiotics. Food Chem. 2016, 213, 721–727. [Google Scholar] [CrossRef]
- Ávila, M.; Hidalgo, M.; Sánchez-Moreno, C.; Pelaez, C.; Requena, T.; de Pascual-Teresa, S. Bioconversion of anthocyanin glycosides by Bifidobacteria and Lactobacillus. Food Res. Int. 2009, 42, 1453–1461. [Google Scholar] [CrossRef]
- Wu, X.; Hu, M.; Hu, X.; Ding, H.; Gong, D.; Zhang, G. Inhibitory mechanism of epicatechin gallate on α-amylase and α-glucosidase and its combinational effect with acarbose or epigallocatechin gallate. J. Mol. Liq. 2019, 290, 111202. [Google Scholar] [CrossRef]
- McDougall, G.J.; Stewart, D. The inhibitory effects of berry polyphenols on digestive enzymes. BioFactors 2005, 23, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Natural Products as α-Amylase and α-Glucosidase Inhibitors and their Hypoglycaemic Potential in the Treatment of Diabetes: An Update. Mini-Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, J.; Li, H.; Shi, S.; Peng, X. Mechanistic study and synergistic effect on inhibition of α-amylase by structurally similar flavonoids. J. Mol. Liq. 2022, 360, 119485. [Google Scholar] [CrossRef]
- Proença, C.; Freitas, M.; Ribeiro, D.; Tomé, S.M.; Oliveira, E.F.T.; Viegas, M.F.; Araújo, A.N.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; et al. Evaluation of a flavonoids library for inhibition of pancreatic α-amylase towards a structure–activity relationship. J. Enzyme Inhib. Med. Chem. 2019, 34, 577–588. [Google Scholar] [CrossRef]
- Lee, S.; Kim, M. Leuconostoc mesenteroides MKSR isolated from kimchi possesses α-glucosidase inhibitory activity, antioxidant activity, and cholesterol-lowering effects. LWT Food Sci. Technol. 2019, 116, 108570. [Google Scholar] [CrossRef]
- Yang, J.; Wang, X.; Zhang, C.; Ma, L.; Wei, T.; Zhao, Y.; Peng, X. Comparative study of inhibition mechanisms of structurally different flavonoid compounds on α-glucosidase and synergistic effect with acarbose. Food Chem. 2021, 347, 129056. [Google Scholar] [CrossRef]
- Hossain, U.; Das, A.K.; Ghosh, S.; Sil, P.C. An overview on the role of bioactive α-glucosidase inhibitors in ameliorating diabetic complications. Food Chem. Toxicol. 2020, 145, 111738. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Yu, W.; Gao, H.; Szeto, I.M.Y.; Feng, H.; Liu, X.; Wang, Y.; Sun, L. Soluble dietary fibres decrease α-glucosidase inhibition of epigallocatechin gallate through affecting polyphenol-enzyme binding interactions. Food Chem. 2023, 409, 135327. [Google Scholar] [CrossRef]
- Paray, B.A.; Rather, I.A.; Al-Sadoon, M.K.; Hamad, A.S.F. Pharmaceutical significance of Leuconostoc mesenteroides KS-TN11 isolated from Nile Tilapia, Oreochromis niloticus. Saudi Pharm. J. 2018, 26, 509–514. [Google Scholar] [CrossRef]
- Singh, M.; Thrimawithana, T.; Shukla, R.; Adhikari, B. Managing obesity through natural polyphenols: A review. Future Foods 2020, 1–2, 100002. [Google Scholar] [CrossRef]
- Liu, S.; Yu, J.; Guo, S.; Fang, H.; Chang, X. Inhibition of pancreatic α-amylase by Lonicera caerulea berry polyphenols in vitro and their potential as hyperglycemic agents. LWT 2020, 126, 109288. [Google Scholar] [CrossRef]
- De Silva, A.B.K.H.; Rupasinghe, H.P.V. Polyphenols composition and anti-diabetic properties in vitro of haskap (Lonicera caerulea L.) berries in relation to cultivar and harvesting date. J. Food Compos. Anal. 2020, 88, 103402. [Google Scholar] [CrossRef]
- Maia, G.A.; Da Silva, L.M.R.; Prado, G.M.D.; Fonseca, A.V.V.; De Sousa, P.H.M.; de Figueiredo, R.W. Development of Mixed Beverages Based on Tropical Fruits. In Non-Alcoholic Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 129–162. [Google Scholar] [CrossRef]
- Darniadi, S.; Ifie, I.; Ho, P.; Murray, B.S. Evaluation of total monomeric anthocyanin, total phenolic content and individual anthocyanins of foam-mat freeze-dried and spray-dried blueberry powder. J. Food Meas. Charact. 2019, 13, 1599–1606. [Google Scholar] [CrossRef]
- Estupiñan-Amaya, M.; Fuenmayor, C.A.; López-Córdoba, A. New Freeze-Dried Andean Blueberry Juice Powders for Potential Application as Functional Food Ingredients: Effect of Maltodextrin on Bioactive and Morphological Features. Molecules 2020, 25, 5635. [Google Scholar] [CrossRef]
- Isas, A.S.; Celis, M.S.M.; Correa, J.R.P.; Fuentes, E.; Rodríguez, L.; Palomo, I.; Mozzi, F.; Van Nieuwenhove, C. Functional fermented cherimoya (Annona cherimola Mill.) juice using autochthonous lactic acid bacteria. Food Res. Int. 2020, 138, 109729. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Juice Type | Glucose (g/L) | Lactic Acid (mg/L) | L. mesenteroides (Log CFU/mL) |
|---|---|---|---|
| JN | 4.7 ± 0.2 a | 22.1 ± 0.1 c | BDL |
| JNF | 1.2 ± 0.1 c | 30.4 ± 0.2 b | 6.8 ± 0.1 a |
| JNF + G | 2.80 ± 0.04 b | 47.9 ± 0.1 a | 6.5 ± 0.2 b |
| JL | 0.77 ± 0.02 a | ND | BDL |
| JLF | 0.1 ± 0.1 c | 13.5 ± 0.2 b | 6.7 ± 0.2 a |
| JLF + G | 0.57 ± 0.04 b | 21.7 ± 0.1 a | 6.9 ± 0.2 a |
| JC | 3.0 ± 0.1 a | 22.3 ± 0.1 c | BDL |
| JCF | 1.74 ± 0.03 c | 31.8 ± 0.1 b | 6.7 ± 0.2 a |
| JCF + G | 2.1 ± 0.1 b | 20.6 ± 0.2 a | 6.8 ± 0.2 a |
| Juice Type | TPC (mg GAE/L) | DPPH IC50 (mg GAE/L) | DPPH % Inhibition | ORAC (µmol TE/mL) |
|---|---|---|---|---|
| JN | 467 ± 5 d | 272 ± 10 a | 71 ± 3 e | 44 ± 2 h |
| JNF | 618 ± 1 a | 123 ± 5 c,d | 80 ± 3 d | 93 ± 4 f |
| JNF + G | 635 ± 5 a | 136 ± 1 c | 74 ± 3 e | 201 ± 5 c |
| JL | 131 ± 2 g | N.D. | 33 ± 1 h | 62 ± 3 g |
| JLF | 322 ± 1 e | 115 ± 2 d | 59.2 ± 0.1 f | 161 ± 30 d |
| JLF + G | 242 ± 1 f | 126 ± 3 c,d | 53.2 ± 0.4 g | 253 ± 30 b |
| JC | 550 ± 3 c | 160 ± 5 b | 86.6 ± 0.4 c | 65 ± 2 g |
| JCF | 479 ± 3 d | 74 ± 10 e | 92.4 ± 0.2 a | 135 ± 30 e |
| JCF + G | 593 ± 2 b | 89 ± 4 e | 87 ± 1 b | 261 ± 30 a |
| Bioactive Compound | |
|---|---|
| Anthocyanins | Delphinidin Cyanidin Petunidin Peonidin Malvidin |
| Flavonoids | Quercertin Rutin Myricetin Kaempferol Luteolin Catechin |
| Hydroxycinnamic acids | Caffeic acid |
| Hydroxybenzoic acids | Gallic acid |
| Juice Type | IC50 (µg/mL) α-Glucosidase |
|---|---|
| JN | 2430 ± 10 c |
| JNF | 805 ± 20 d |
| JNF + G | 2390 ± 20 c |
| JL | 3588 ± 20 a |
| JLF | 760 ± 20 e |
| JLF + G | 2663 ± 20 b |
| JC | 820 ± 10 d |
| JCF | 332 ± 4 g |
| JCF + G | 790 ± 10 e |
| Acarbose (control) | 699 ± 4 f |
| Juice Type | pH | TSS | TTA (% Ascorbic Acid) | Glucose (g/L) | Fructose (g/L) |
|---|---|---|---|---|---|
| JN | 3.5 ± 0.1 b | 1.7 ± 0.4 a | 0.09 ± 0.02 a | 4.7 ± 0.1 a | 2.8 ± 0.1 b |
| JL | 4.8 ± 0.1 a | 1.6 ± 0.1 a | 0.06 ± 0.02 a | 0.77 ± 0.02 c | 1.47 ± 0.04 c |
| JC | 3.6 ± 0.1 b | 1.7 ± 0.3 a | 0.09 ± 0.03 a | 3.0 ± 0.1 b | 3.8 ± 0.1 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escobar-Beiza, N.; Pérez-Correa, J.R.; Franco, W. Fermentation of Murta (Ugni molinae) Juice: Effect on Antioxidant Activity and Control of Enzymes Associated with Glucose Assimilation. Int. J. Mol. Sci. 2023, 24, 15197. https://doi.org/10.3390/ijms242015197
Escobar-Beiza N, Pérez-Correa JR, Franco W. Fermentation of Murta (Ugni molinae) Juice: Effect on Antioxidant Activity and Control of Enzymes Associated with Glucose Assimilation. International Journal of Molecular Sciences. 2023; 24(20):15197. https://doi.org/10.3390/ijms242015197
Chicago/Turabian StyleEscobar-Beiza, Natalia, José R. Pérez-Correa, and Wendy Franco. 2023. "Fermentation of Murta (Ugni molinae) Juice: Effect on Antioxidant Activity and Control of Enzymes Associated with Glucose Assimilation" International Journal of Molecular Sciences 24, no. 20: 15197. https://doi.org/10.3390/ijms242015197
APA StyleEscobar-Beiza, N., Pérez-Correa, J. R., & Franco, W. (2023). Fermentation of Murta (Ugni molinae) Juice: Effect on Antioxidant Activity and Control of Enzymes Associated with Glucose Assimilation. International Journal of Molecular Sciences, 24(20), 15197. https://doi.org/10.3390/ijms242015197