
A Focused Review of Ras Guanine Nucleotide-Releasing Protein 1 in Immune Cells and Cancer
 , , , , and
, , , , and 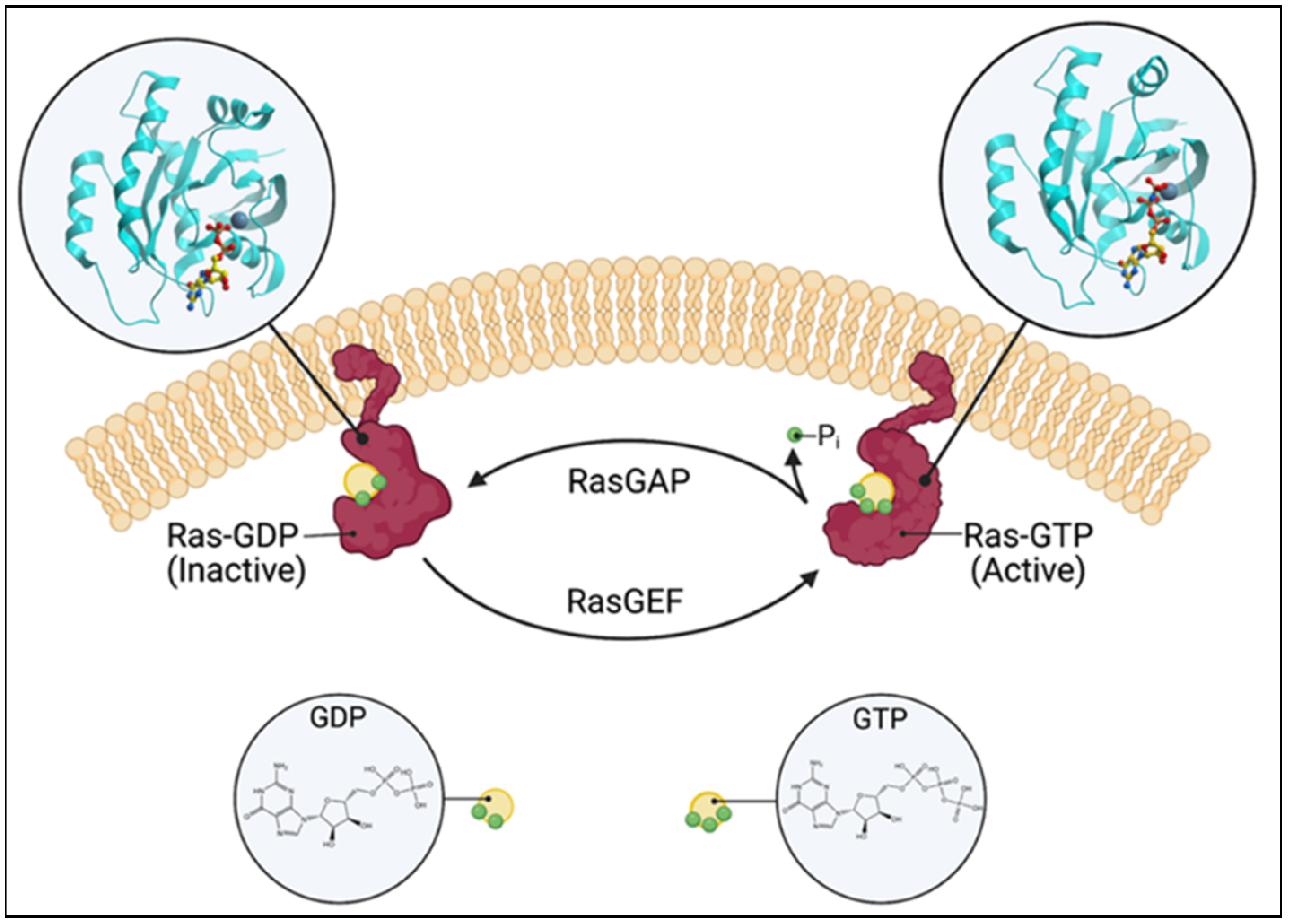{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
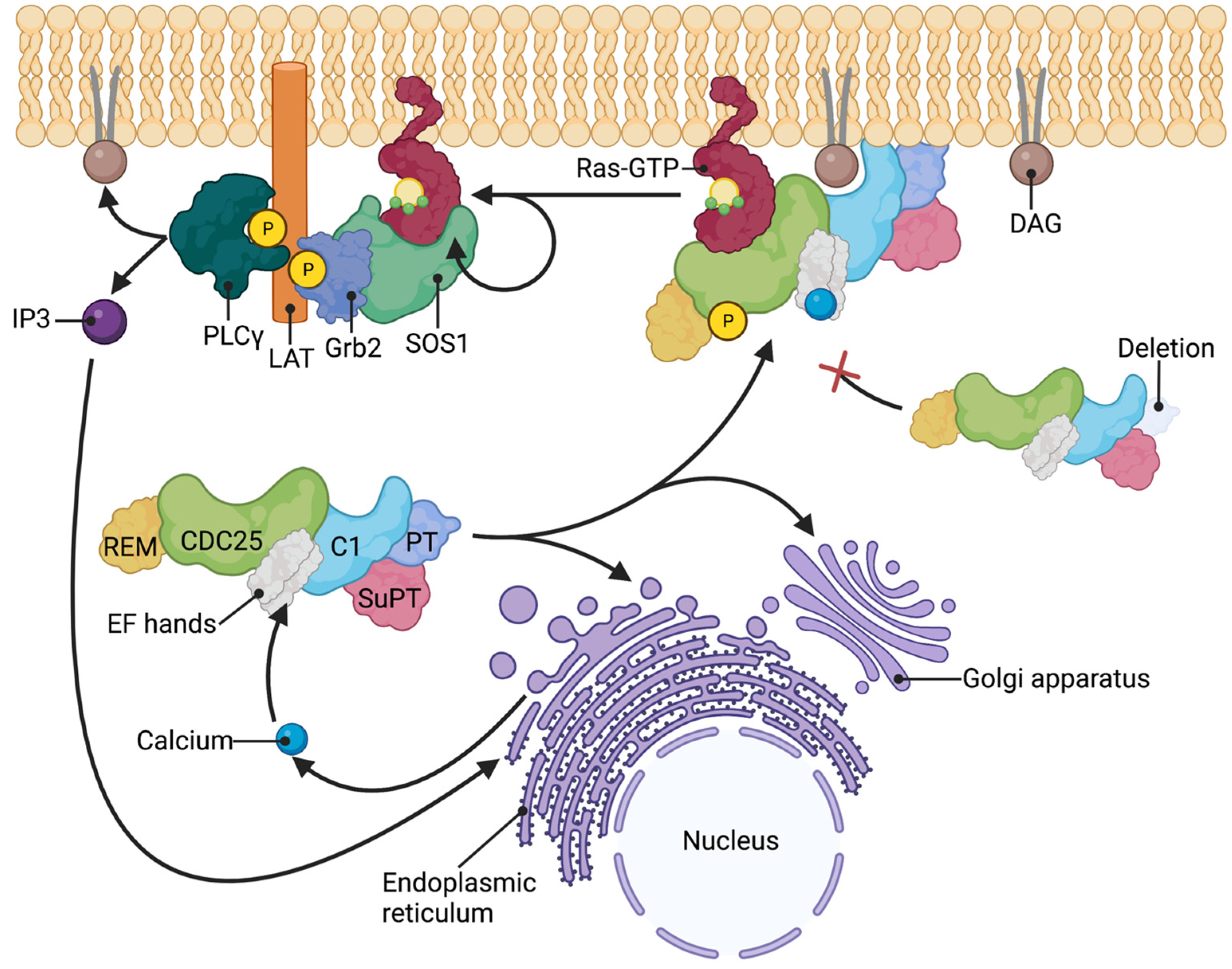
Abstract
1. Ras Guanine Nucleotide Exchange Factors: Introduction
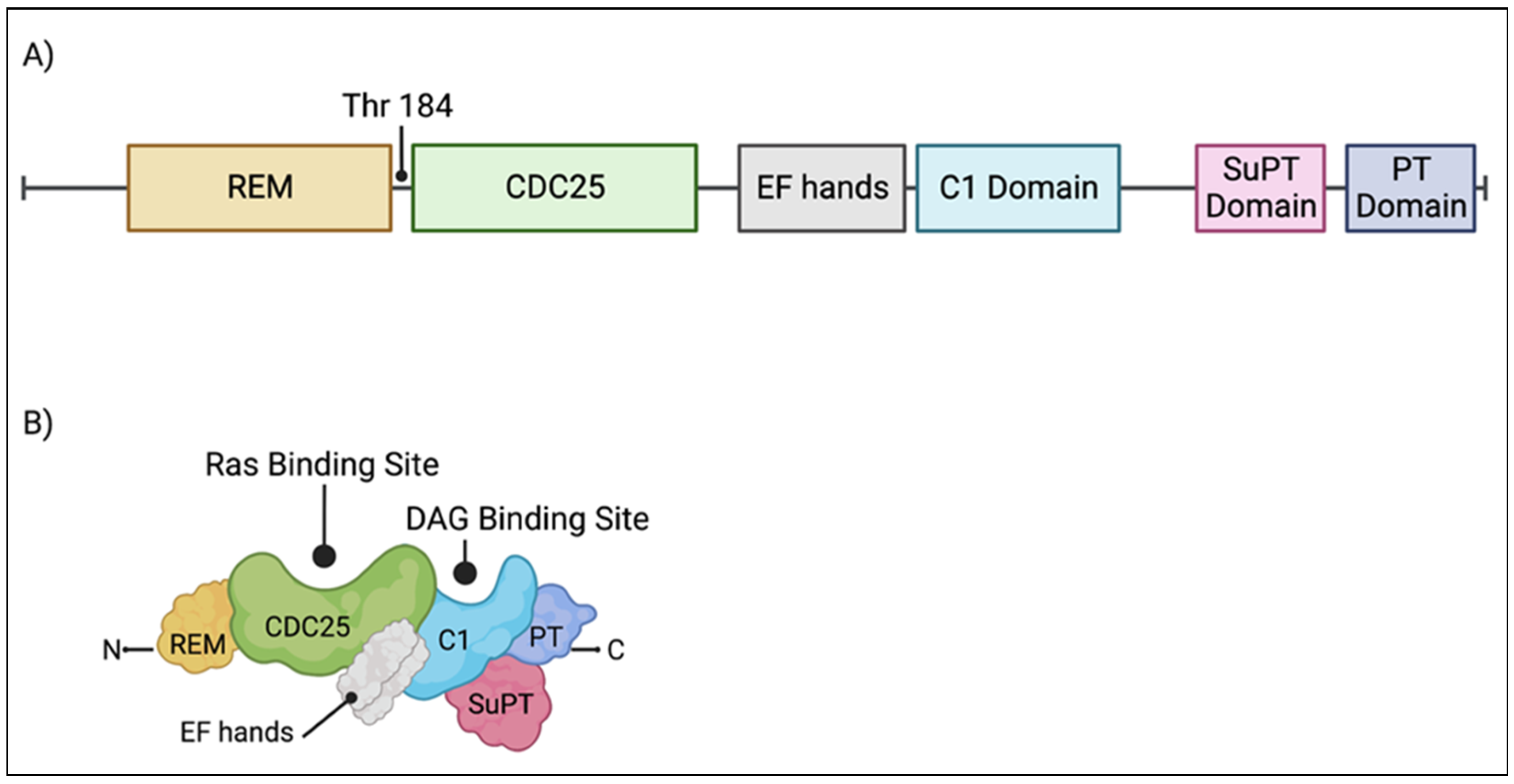
2. RasGRP1: Structure and Function
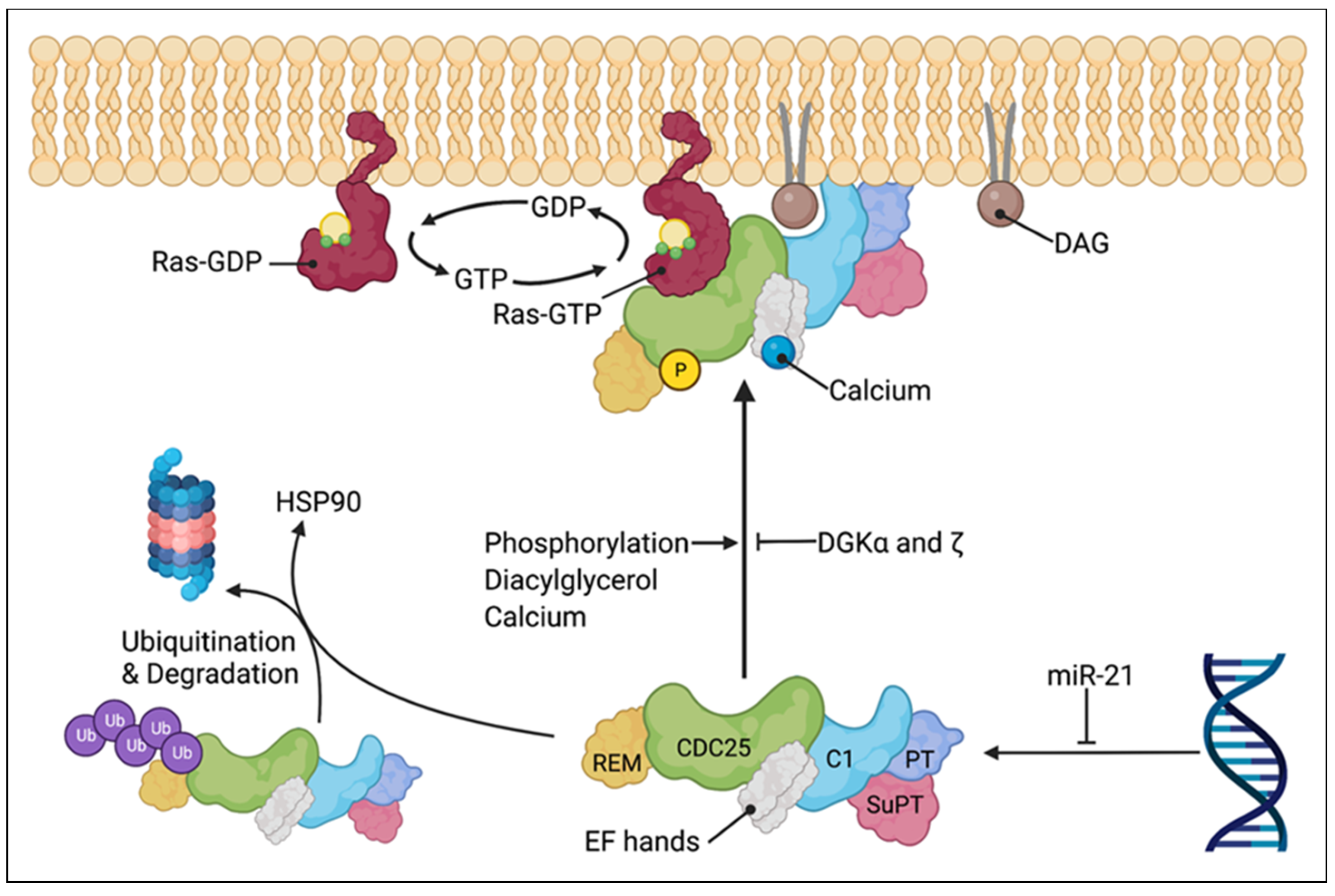
3. Ras Guanine Nucleotide-Releasing Protein 1: Regulation
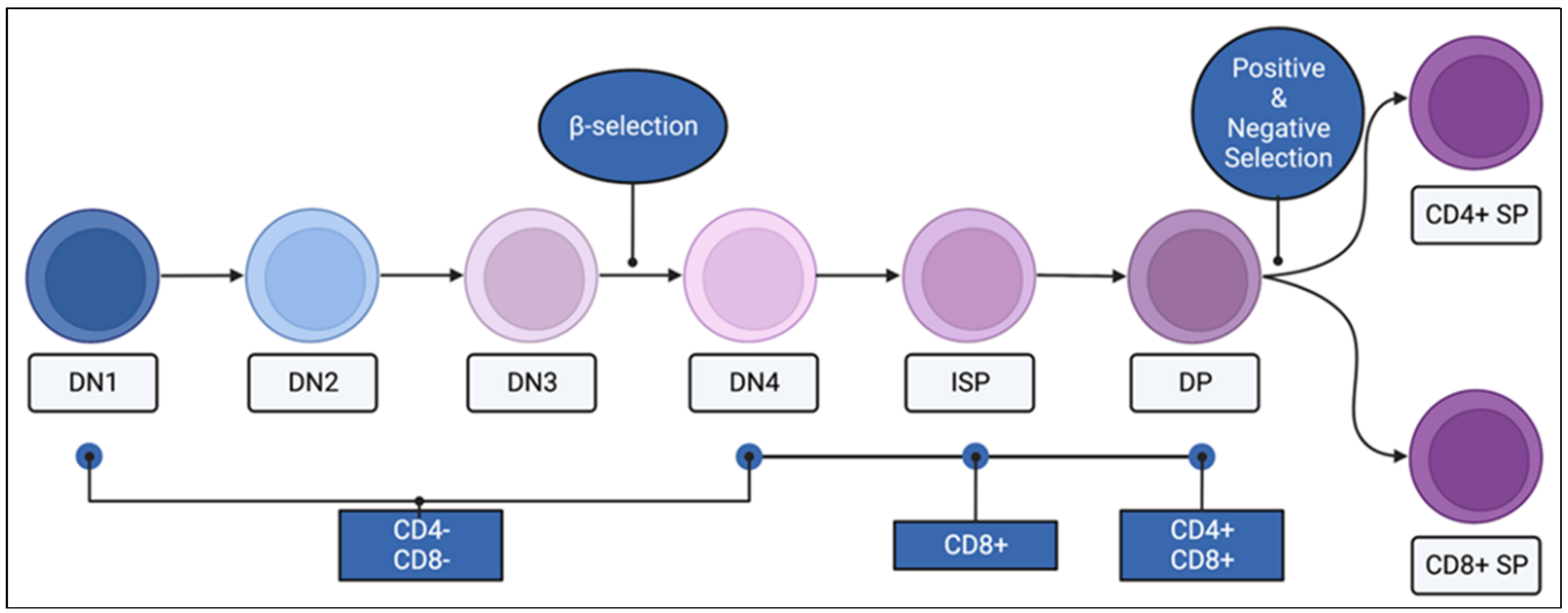
4. RasGRP1: Cell Development and Function
4.1. Immature Thymocytes
4.2. T Cells
4.3. B-Cells
4.4. NK Cells
4.5. Granulocytes
5. RasGRP1: Role in Cancer
5.1. Lymphoma and Leukemia
5.2. Squamous Cell Carcinoma
5.3. Colorectal Cancer
5.4. Hepatocellular Carcinoma
5.5. Breast Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Kawasaki, H.; Springett, G.M.; Mochizuki, N.; Toki, S.; Nakaya, M.; Matsuda, M.; Housman, D.E.; Graybiel, A.M. A family of cAMP-binding proteins that directly activate Rap1. Science 1998, 282, 2275–2279. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Philips, M.R. Compartmentalized Ras/MAPK signaling. Annu. Rev. Immunol. 2006, 24, 771–800. [Google Scholar] [CrossRef]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef]
- Stone, J.C. Regulation and Function of the RasGRP Family of Ras Activators in Blood Cells. Genes Cancer 2011, 2, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.E.; Rubio, I.; Roose, J.P. Regulation of ras exchange factors and cellular localization of ras activation by lipid messengers in T cells. Front. Immunol. 2013, 4, 239. [Google Scholar] [CrossRef]
- Ksionda, O.; Limnander, A.; Roose, J.P. RasGRP Ras guanine nucleotide exchange factors in cancer. Front. Biol. 2013, 8, 508–532. [Google Scholar] [CrossRef]
- Quilliam, L.A.; Rebhun, J.F.; Castro, A.F. A growing family of guanine nucleotide exchange factors is responsible for activation of Ras-family GTPases. Prog. Nucleic Acid Res. Mol. Biol. 2002, 71, 391–444. [Google Scholar] [CrossRef] [PubMed]
- Vanoni, M.; Bertini, R.; Sacco, E.; Fontanella, L.; Rieppi, M.; Colombo, S.; Martegani, E.; Carrera, V.; Moroni, A.; Bizzarri, C.; et al. Characterization and properties of dominant-negative mutants of the ras-specific guanine nucleotide exchange factor CDC25(Mm). J. Biol. Chem. 1999, 274, 36656–36662. [Google Scholar] [CrossRef]
- Bossu, P.; Vanoni, M.; Wanke, V.; Cesaroni, M.P.; Tropea, F.; Melillo, G.; Asti, C.; Porzio, S.; Ruggiero, P.; Di Cioccio, V.; et al. A dominant negative RAS-specific guanine nucleotide exchange factor reverses neoplastic phenotype in K-ras transformed mouse fibroblasts. Oncogene 2000, 19, 2147–2154. [Google Scholar] [CrossRef]
- Sacco, E.; Fantinato, S.; Manzoni, R.; Metalli, D.; De Gioia, L.; Fantucci, P.; Alberghina, L.; Vanoni, M. The isolated catalytic hairpin of the Ras-specific guanine nucleotide exchange factor Cdc25Mm retains nucleotide dissociation activity but has impaired nucleotide exchange activity. FEBS Lett. 2005, 579, 6851–6858. [Google Scholar] [CrossRef]
- Ebinu, J.O.; Bottorff, D.A.; Chan, E.Y.; Stang, S.L.; Dunn, R.J.; Stone, J.C. RasGRP, a Ras guanyl nucleotide- releasing protein with calcium- and diacylglycerol-binding motifs. Science 1998, 280, 1082–1086. [Google Scholar] [CrossRef] [PubMed]
- Tognon, C.E.; Kirk, H.E.; Passmore, L.A.; Whitehead, I.P.; Der, C.J.; Kay, R.J. Regulation of RasGRP via a phorbol ester-responsive C1 domain. Mol. Cell. Biol. 1998, 18, 6995–7008. [Google Scholar] [CrossRef]
- Ebinu, J.O.; Stang, S.L.; Teixeira, C.; Bottorff, D.A.; Hooton, J.; Blumberg, P.M.; Barry, M.; Bleakley, R.C.; Ostergaard, H.L.; Stone, J.C. RasGRP links T-cell receptor signaling to Ras. Blood 2000, 95, 3199–3203. [Google Scholar] [CrossRef]
- Tazmini, G.; Beaulieu, N.; Woo, A.; Zahedi, B.; Goulding, R.E.; Kay, R.J. Membrane localization of RasGRP1 is controlled by an EF-hand, and by the GEF domain. Biochim. Biophys. Acta 2009, 1793, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Guilbault, B.; Kay, R.J. RasGRP1 sensitizes an immature B cell line to antigen receptor-induced apoptosis. J. Biol. Chem. 2004, 279, 19523–19530. [Google Scholar] [CrossRef] [PubMed]
- Iwig, J.S.; Vercoulen, Y.; Das, R.; Barros, T.; Limnander, A.; Che, Y.; Pelton, J.G.; Wemmer, D.E.; Roose, J.P.; Kuriyan, J. Structural analysis of autoinhibition in the Ras-specific exchange factor RasGRP1. eLife 2013, 2, e00813. [Google Scholar] [CrossRef] [PubMed]
- Cullen, P.J.; Lockyer, P.J. Integration of calcium and Ras signalling. Nat. Rev. Mol. Cell Biol. 2002, 3, 339–348. [Google Scholar] [CrossRef]
- Feske, S. Calcium signalling in lymphocyte activation and disease. Nat. Rev. Immunol. 2007, 7, 690–702. [Google Scholar] [CrossRef]
- Rambaratsingh, R.A.; Stone, J.C.; Blumberg, P.M.; Lorenzo, P.S. RasGRP1 represents a novel non-protein kinase C phorbol ester signaling pathway in mouse epidermal keratinocytes. J. Biol. Chem. 2003, 278, 52792–52801. [Google Scholar] [CrossRef]
- Tuthill, M.C.; Oki, C.E.; Lorenzo, P.S. Differential effects of bryostatin 1 and 12-O-tetradecanoylphorbol-13-acetate on the regulation and activation of RasGRP1 in mouse epidermal keratinocytes. Mol. Cancer Ther. 2006, 5, 602–610. [Google Scholar] [CrossRef]
- Johnson, J.E.; Goulding, R.E.; Ding, Z.; Partovi, A.; Anthony, K.V.; Beaulieu, N.; Tazmini, G.; Cornell, R.B.; Kay, R.J. Differential membrane binding and diacylglycerol recognition by C1 domains of RasGRPs. Biochem. J. 2007, 406, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Elhalem, E.; Donadio, L.G.; Zhou, X.; Lewin, N.E.; Garcia, L.C.; Lai, C.C.; Kelley, J.A.; Peach, M.L.; Blumberg, P.M.; Comin, M.J. Exploring the influence of indololactone structure on selectivity for binding to the C1 domains of PKCalpha, PKCepsilon, and RasGRP. Bioorg. Med. Chem. 2017, 25, 2971–2980. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, N.; Zahedi, B.; Goulding, R.E.; Tazmini, G.; Anthony, K.V.; Omeis, S.L.; de Jong, D.R.; Kay, R.J. Regulation of RasGRP1 by B cell antigen receptor requires cooperativity between three domains controlling translocation to the plasma membrane. Mol. Biol. Cell 2007, 18, 3156–3168. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Campi, G.; Du, G.; Zheng, Y.; Foster, D.A.; Dustin, M.L.; Philips, M.R. The lymphocyte function-associated antigen-1 receptor costimulates plasma membrane Ras via phospholipase D2. Nat. Cell Biol. 2007, 9, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Caloca, M.J.; Zugaza, J.L.; Bustelo, X.R. Exchange factors of the RasGRP family mediate Ras activation in the Golgi. J. Biol. Chem. 2003, 278, 33465–33473. [Google Scholar] [CrossRef]
- Zugaza, J.L.; Caloca, M.J.; Bustelo, X.R. Inverted signaling hierarchy between RAS and RAC in T-lymphocytes. Oncogene 2004, 23, 5823–5833. [Google Scholar] [CrossRef]
- Bivona, T.G.; Perez De Castro, I.; Ahearn, I.M.; Grana, T.M.; Chiu, V.K.; Lockyer, P.J.; Cullen, P.J.; Pellicer, A.; Cox, A.D.; Philips, M.R. Phospholipase Cgamma activates Ras on the Golgi apparatus by means of RasGRP1. Nature 2003, 424, 694–698. [Google Scholar] [CrossRef]
- Choy, E.; Chiu, V.K.; Silletti, J.; Feoktistov, M.; Morimoto, T.; Michaelson, D.; Ivanov, I.E.; Philips, M.R. Endomembrane trafficking of ras: The CAAX motif targets proteins to the ER and Golgi. Cell 1999, 98, 69–80. [Google Scholar] [CrossRef]
- Chiu, V.K.; Bivona, T.; Hach, A.; Sajous, J.B.; Silletti, J.; Wiener, H.; Johnson, R.L., 2nd; Cox, A.D.; Philips, M.R. Ras signalling on the endoplasmic reticulum and the Golgi. Nat. Cell Biol. 2002, 4, 343–350. [Google Scholar] [CrossRef]
- Zahedi, B.; Goo, H.J.; Beaulieu, N.; Tazmini, G.; Kay, R.J.; Cornell, R.B. Phosphoinositide 3-kinase regulates plasma membrane targeting of the Ras-specific exchange factor RasGRP1. J. Biol. Chem. 2011, 286, 12712–12723. [Google Scholar] [CrossRef]
- Fuller, D.M.; Zhu, M.; Song, X.; Ou-Yang, C.W.; Sullivan, S.A.; Stone, J.C.; Zhang, W. Regulation of RasGRP1 function in T cell development and activation by its unique tail domain. PLoS ONE 2012, 7, e38796. [Google Scholar] [CrossRef]
- Sanjuan, M.A.; Jones, D.R.; Izquierdo, M.; Merida, I. Role of diacylglycerol kinase alpha in the attenuation of receptor signaling. J. Cell Biol. 2001, 153, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan, M.A.; Pradet-Balade, B.; Jones, D.R.; Martinez, A.C.; Stone, J.C.; Garcia-Sanz, J.A.; Merida, I. T cell activation in vivo targets diacylglycerol kinase alpha to the membrane: A novel mechanism for Ras attenuation. J. Immunol. 2003, 170, 2877–2883. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.R.; Sanjuan, M.A.; Stone, J.C.; Merida, I. Expression of a catalytically inactive form of diacylglycerol kinase alpha induces sustained signaling through RasGRP. FASEB J. 2002, 16, 595–597. [Google Scholar] [CrossRef] [PubMed]
- Zha, Y.; Marks, R.; Ho, A.W.; Peterson, A.C.; Janardhan, S.; Brown, I.; Praveen, K.; Stang, S.; Stone, J.C.; Gajewski, T.F. T cell anergy is reversed by active Ras and is regulated by diacylglycerol kinase-alpha. Nat. Immunol. 2006, 7, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Olenchock, B.A.; Guo, R.; Carpenter, J.H.; Jordan, M.; Topham, M.K.; Koretzky, G.A.; Zhong, X.P. Disruption of diacylglycerol metabolism impairs the induction of T cell anergy. Nat. Immunol. 2006, 7, 1174–1181. [Google Scholar] [CrossRef]
- Topham, M.K.; Prescott, S.M. Diacylglycerol kinase zeta regulates Ras activation by a novel mechanism. J. Cell Biol. 2001, 152, 1135–1143. [Google Scholar] [CrossRef]
- Zhong, X.P.; Hainey, E.A.; Olenchock, B.A.; Zhao, H.; Topham, M.K.; Koretzky, G.A. Regulation of T cell receptor-induced activation of the Ras-ERK pathway by diacylglycerol kinase zeta. J. Biol. Chem. 2002, 277, 31089–31098. [Google Scholar] [CrossRef]
- Sakane, F.; Imai, S.; Kai, M.; Yasuda, S.; Kanoh, H. Diacylglycerol kinases: Why so many of them? Biochim. Biophys. Acta 2007, 1771, 793–806. [Google Scholar] [CrossRef]
- Merida, I.; Arranz-Nicolas, J.; Torres-Ayuso, P.; Avila-Flores, A. Diacylglycerol Kinase Malfunction in Human Disease and the Search for Specific Inhibitors. Handb. Exp. Pharmacol. 2020, 259, 133–162. [Google Scholar] [CrossRef]
- Roose, J.P.; Mollenauer, M.; Gupta, V.A.; Stone, J.; Weiss, A. A diacylglycerol-protein kinase C-RasGRP1 pathway directs Ras activation upon antigen receptor stimulation of T cells. Mol. Cell. Biol. 2005, 25, 4426–4441. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, H.; Coughlin, J.; Zheng, J.; Li, L.; Stone, J.C. Phosphorylation of RasGRP3 on threonine 133 provides a mechanistic link between PKC and Ras signaling systems in B cells. Blood 2005, 105, 3648–3654. [Google Scholar] [CrossRef]
- Luo, B.; Prescott, S.M.; Topham, M.K. Association of diacylglycerol kinase zeta with protein kinase C alpha: Spatial regulation of diacylglycerol signaling. J. Cell Biol. 2003, 160, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Vercoulen, Y.; Kondo, Y.; Iwig, J.S.; Janssen, A.B.; White, K.A.; Amini, M.; Barber, D.L.; Kuriyan, J.; Roose, J.P. A Histidine pH sensor regulates activation of the Ras-specific guanine nucleotide exchange factor RasGRP1. eLife 2017, 6, e29002. [Google Scholar] [CrossRef] [PubMed]
- Griner, E.M.; Kazanietz, M.G. Protein kinase C and other diacylglycerol effectors in cancer. Nat. Rev. Cancer 2007, 7, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Peterson, K.L.; Correia, C.; Koh, B.; Schneider, P.A.; Nowakowski, G.S.; Kaufmann, S.H. Histone deacetylase inhibitors interrupt HSP90*RASGRP1 and HSP90*CRAF interactions to upregulate BIM and circumvent drug resistance in lymphoma cells. Leukemia 2017, 31, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Wasik, U.; Kempinska-Podhorodecka, A.; Bogdanos, D.P.; Milkiewicz, P.; Milkiewicz, M. Enhanced expression of miR-21 and miR-150 is a feature of anti-mitochondrial antibody-negative primary biliary cholangitis. Mol. Med. 2020, 26, 8. [Google Scholar] [CrossRef]
- Pan, W.; Zhu, S.; Yuan, M.; Cui, H.; Wang, L.; Luo, X.; Li, J.; Zhou, H.; Tang, Y.; Shen, N. MicroRNA-21 and microRNA-148a contribute to DNA hypomethylation in lupus CD4+ T cells by directly and indirectly targeting DNA methyltransferase 1. J. Immunol. 2010, 184, 6773–6781. [Google Scholar] [CrossRef]
- Wickramasinghe, N.S.; Manavalan, T.T.; Dougherty, S.M.; Riggs, K.A.; Li, Y.; Klinge, C.M. Estradiol downregulates miR-21 expression and increases miR-21 target gene expression in MCF-7 breast cancer cells. Nucleic Acids Res. 2009, 37, 2584–2595. [Google Scholar] [CrossRef]
- Swat, W.; Shinkai, Y.; Cheng, H.L.; Davidson, L.; Alt, F.W. Activated Ras signals differentiation and expansion of CD4+8+ thymocytes. Proc. Natl. Acad. Sci. USA 1996, 93, 4683–4687. [Google Scholar] [CrossRef]
- Dower, N.A.; Stang, S.L.; Bottorff, D.A.; Ebinu, J.O.; Dickie, P.; Ostergaard, H.L.; Stone, J.C. RasGRP essential for mouse thymocyte differentiation and TCR signaling. Nat. Immunol. 2000, 1, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Kortum, R.L.; Sommers, C.L.; Alexander, C.P.; Pinski, J.M.; Li, W.; Grinberg, A.; Lee, J.; Love, P.E.; Samelson, L.E. Targeted Sos1 deletion reveals its critical role in early T-cell development. Proc. Natl. Acad. Sci. USA 2011, 108, 12407–12412. [Google Scholar] [CrossRef] [PubMed]
- Kortum, R.L.; Sommers, C.L.; Pinski, J.M.; Alexander, C.P.; Merrill, R.K.; Li, W.; Love, P.E.; Samelson, L.E. Deconstructing Ras signaling in the thymus. Mol. Cell. Biol. 2012, 32, 2748–2759. [Google Scholar] [CrossRef] [PubMed]
- Priatel, J.J.; Teh, S.J.; Dower, N.A.; Stone, J.C.; Teh, H.S. RasGRP1 transduces low-grade TCR signals which are critical for T cell development, homeostasis, and differentiation. Immunity 2002, 17, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Daniels, M.A.; Teixeiro, E.; Gill, J.; Hausmann, B.; Roubaty, D.; Holmberg, K.; Werlen, G.; Hollander, G.A.; Gascoigne, N.R.; Palmer, E. Thymic selection threshold defined by compartmentalization of Ras/MAPK signalling. Nature 2006, 444, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Zikherman, J.; Das, J.; Roose, J.P.; Weiss, A.; Chakraborty, A.K. Origin of the sharp boundary that discriminates positive and negative selection of thymocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 528–533. [Google Scholar] [CrossRef]
- Das, J.; Ho, M.; Zikherman, J.; Govern, C.; Yang, M.; Weiss, A.; Chakraborty, A.K.; Roose, J.P. Digital signaling and hysteresis characterize ras activation in lymphoid cells. Cell 2009, 136, 337–351. [Google Scholar] [CrossRef]
- Salzer, E.; Cagdas, D.; Hons, M.; Mace, E.M.; Garncarz, W.; Petronczki, O.Y.; Platzer, R.; Pfajfer, L.; Bilic, I.; Ban, S.A.; et al. RASGRP1 deficiency causes immunodeficiency with impaired cytoskeletal dynamics. Nat. Immunol. 2016, 17, 1352–1360. [Google Scholar] [CrossRef]
- Coughlin, J.J.; Stang, S.L.; Dower, N.A.; Stone, J.C. RasGRP1 and RasGRP3 regulate B cell proliferation by facilitating B cell receptor-Ras signaling. J. Immunol. 2005, 175, 7179–7184. [Google Scholar] [CrossRef]
- Coughlin, J.J.; Stang, S.L.; Dower, N.A.; Stone, J.C. The role of RasGRPs in regulation of lymphocyte proliferation. Immunol. Lett. 2006, 105, 77–82. [Google Scholar] [CrossRef]
- Baars, M.J.D.; Douma, T.; Simeonov, D.R.; Myers, D.R.; Kulhanek, K.; Banerjee, S.; Zwakenberg, S.; Baltissen, M.P.; Amini, M.; de Roock, S.; et al. Dysregulated RASGRP1 expression through RUNX1 mediated transcription promotes autoimmunity. Eur. J. Immunol. 2021, 51, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Oh-hora, M.; Johmura, S.; Hashimoto, A.; Hikida, M.; Kurosaki, T. Requirement for Ras guanine nucleotide releasing protein 3 in coupling phospholipase C-gamma2 to Ras in B cell receptor signaling. J. Exp. Med. 2003, 198, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, H.; Takahashi, Y.; Hayashi, R.; Nakamura, T.; Ishii, K.; Matsuda, J.; Ogura, A.; Shirakata, Y.; Karasuyama, H.; Sudo, T.; et al. Ras mediates effector pathways responsible for pre-B cell survival, which is essential for the developmental progression to the late pre-B cell stage. J. Exp. Med. 2000, 192, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Iritani, B.M.; Forbush, K.A.; Farrar, M.A.; Perlmutter, R.M. Control of B cell development by Ras-mediated activation of Raf. EMBO J. 1997, 16, 7019–7031. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Nunes, J.A.; Vely, F. Natural killer cell signaling pathways. Science 2004, 306, 1517–1519. [Google Scholar] [CrossRef]
- Lee, S.H.; Yun, S.; Lee, J.; Kim, M.J.; Piao, Z.H.; Jeong, M.; Chung, J.W.; Kim, T.D.; Yoon, S.R.; Greenberg, P.D.; et al. RasGRP1 is required for human NK cell function. J. Immunol. 2009, 183, 7931–7938. [Google Scholar] [CrossRef]
- Hock, H.; Hamblen, M.J.; Rooke, H.M.; Traver, D.; Bronson, R.T.; Cameron, S.; Orkin, S.H. Intrinsic requirement for zinc finger transcription factor Gfi-1 in neutrophil differentiation. Immunity 2003, 18, 109–120. [Google Scholar] [CrossRef]
- Karsunky, H.; Zeng, H.; Schmidt, T.; Zevnik, B.; Kluge, R.; Schmid, K.W.; Duhrsen, U.; Moroy, T. Inflammatory reactions and severe neutropenia in mice lacking the transcriptional repressor Gfi1. Nat. Genet. 2002, 30, 295–300. [Google Scholar] [CrossRef]
- Person, R.E.; Li, F.Q.; Duan, Z.; Benson, K.F.; Wechsler, J.; Papadaki, H.A.; Eliopoulos, G.; Kaufman, C.; Bertolone, S.J.; Nakamoto, B.; et al. Mutations in proto-oncogene GFI1 cause human neutropenia and target ELA2. Nat. Genet. 2003, 34, 308–312. [Google Scholar] [CrossRef]
- Ward, A.C.; van Aesch, Y.M.; Gits, J.; Schelen, A.M.; de Koning, J.P.; van Leeuwen, D.; Freedman, M.H.; Touw, I.P. Novel point mutation in the extracellular domain of the granulocyte colony-stimulating factor (G-CSF) receptor in a case of severe congenital neutropenia hyporesponsive to G-CSF treatment. J. Exp. Med. 1999, 190, 497–507. [Google Scholar] [CrossRef]
- Liu, F.; Wu, H.Y.; Wesselschmidt, R.; Kornaga, T.; Link, D.C. Impaired production and increased apoptosis of neutrophils in granulocyte colony-stimulating factor receptor-deficient mice. Immunity 1996, 5, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Grail, D.; Hodgson, G.; Metcalf, D.; Stanley, E.; Cheers, C.; Fowler, K.J.; Basu, S.; Zhan, Y.F.; Dunn, A.R. Mice lacking granulocyte colony-stimulating factor have chronic neutropenia, granulocyte and macrophage progenitor cell deficiency, and impaired neutrophil mobilization. Blood 1994, 84, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- de la Luz Sierra, M.; Sakakibara, S.; Gasperini, P.; Salvucci, O.; Jiang, K.; McCormick, P.J.; Segarra, M.; Stone, J.; Maric, D.; Zhu, J.; et al. The transcription factor Gfi1 regulates G-CSF signaling and neutrophil development through the Ras activator RasGRP1. Blood 2010, 115, 3970–3979. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, M.; Nishida, K.; Hirano, T.; Zhang, W. An essential role for RasGRP1 in mast cell function and IgE-mediated allergic response. J. Exp. Med. 2007, 204, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Yang, W.; Latour, S.; Yang, J.; Winter, S.; Zheng, J.; Ni, K.; Lv, M.; Liu, C.; Huang, H.; et al. RASGRP1 mutation in autoimmune lymphoproliferative syndrome-like disease. J. Allergy Clin. Immunol. 2018, 142, 595–604.e516. [Google Scholar] [CrossRef]
- Somekh, I.; Marquardt, B.; Liu, Y.; Rohlfs, M.; Hollizeck, S.; Karakukcu, M.; Unal, E.; Yilmaz, E.; Patiroglu, T.; Cansever, M.; et al. Novel Mutations in RASGRP1 are Associated with Immunodeficiency, Immune Dysregulation, and EBV-Induced Lymphoma. J. Clin. Immunol. 2018, 38, 699–710. [Google Scholar] [CrossRef]
- Winter, S.; Martin, E.; Boutboul, D.; Lenoir, C.; Boudjemaa, S.; Petit, A.; Picard, C.; Fischer, A.; Leverger, G.; Latour, S. Loss of RASGRP1 in humans impairs T-cell expansion leading to Epstein-Barr virus susceptibility. EMBO Mol. Med. 2018, 10, 188–199. [Google Scholar] [CrossRef]
- Oki, T.; Kitaura, J.; Watanabe-Okochi, N.; Nishimura, K.; Maehara, A.; Uchida, T.; Komeno, Y.; Nakahara, F.; Harada, Y.; Sonoki, T.; et al. Aberrant expression of RasGRP1 cooperates with gain-of-function NOTCH1 mutations in T-cell leukemogenesis. Leukemia 2012, 26, 1038–1045. [Google Scholar] [CrossRef]
- Hartzell, C.; Ksionda, O.; Lemmens, E.; Coakley, K.; Yang, M.; Dail, M.; Harvey, R.C.; Govern, C.; Bakker, J.; Lenstra, T.L.; et al. Dysregulated RasGRP1 responds to cytokine receptor input in T cell leukemogenesis. Sci. Signal. 2013, 6, ra21. [Google Scholar] [CrossRef]
- Kim, R.; Trubetskoy, A.; Suzuki, T.; Jenkins, N.A.; Copeland, N.G.; Lenz, J. Genome-based identification of cancer genes by proviral tagging in mouse retrovirus-induced T-cell lymphomas. J. Virol. 2003, 77, 2056–2062. [Google Scholar] [CrossRef]
- Dupuy, A.J.; Morgan, K.; von Lintig, F.C.; Shen, H.; Acar, H.; Hasz, D.E.; Jenkins, N.A.; Copeland, N.G.; Boss, G.R.; Largaespada, D.A. Activation of the Rap1 guanine nucleotide exchange gene, CalDAG-GEF I, in BXH-2 murine myeloid leukemia. J. Biol. Chem. 2001, 276, 11804–11811. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Shen, H.; Akagi, K.; Morse, H.C.; Malley, J.D.; Naiman, D.Q.; Jenkins, N.A.; Copeland, N.G. New genes involved in cancer identified by retroviral tagging. Nat. Genet. 2002, 32, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Klinger, M.B.; Guilbault, B.; Goulding, R.E.; Kay, R.J. Deregulated expression of RasGRP1 initiates thymic lymphomagenesis independently of T-cell receptors. Oncogene 2005, 24, 2695–2704. [Google Scholar] [CrossRef] [PubMed]
- Ksionda, O.; Melton, A.A.; Bache, J.; Tenhagen, M.; Bakker, J.; Harvey, R.; Winter, S.S.; Rubio, I.; Roose, J.P. RasGRP1 overexpression in T-ALL increases basal nucleotide exchange on Ras rendering the Ras/PI3K/Akt pathway responsive to protumorigenic cytokines. Oncogene 2016, 35, 3658–3668. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Yun, G.; Hebert, A.; Kong, G.; Ranheim, E.A.; Finn, R.; Rajagoplan, A.; Li, S.; Zhou, Y.; Yu, M.; et al. Nras Q61R/+ and Kras-/- cooperate to downregulate Rasgrp1 and promote lympho-myeloid leukemia in early T-cell precursors. Blood 2021, 137, 3259–3271. [Google Scholar] [CrossRef] [PubMed]
- Karra, L.; Romero-Moya, D.; Ksionda, O.; Krush, M.; Gu, Z.; Mues, M.; Depeille, P.; Mullighan, C.; Roose, J.P. Increased baseline RASGRP1 signals enhance stem cell fitness during native hematopoiesis. Oncogene 2020, 39, 6920–6934. [Google Scholar] [CrossRef]
- Knudson, A.G., Jr. Mutation and cancer: Statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [Google Scholar] [CrossRef]
- Guo, R.; Wan, C.K.; Carpenter, J.H.; Mousallem, T.; Boustany, R.M.; Kuan, C.T.; Burks, A.W.; Zhong, X.P. Synergistic control of T cell development and tumor suppression by diacylglycerol kinase alpha and zeta. Proc. Natl. Acad. Sci. USA 2008, 105, 11909–11914. [Google Scholar] [CrossRef]
- Lauchle, J.O.; Kim, D.; Le, D.T.; Akagi, K.; Crone, M.; Krisman, K.; Warner, K.; Bonifas, J.M.; Li, Q.; Coakley, K.M.; et al. Response and resistance to MEK inhibition in leukaemias initiated by hyperactive Ras. Nature 2009, 461, 411–414. [Google Scholar] [CrossRef]
- Ksionda, O.; Mues, M.; Wandler, A.M.; Donker, L.; Tenhagen, M.; Jun, J.; Ducker, G.S.; Matlawska-Wasowska, K.; Shannon, K.; Shokat, K.M.; et al. Comprehensive analysis of T cell leukemia signals reveals heterogeneity in the PI3 kinase-Akt pathway and limitations of PI3 kinase inhibitors as monotherapy. PLoS ONE 2018, 13, e0193849. [Google Scholar] [CrossRef]
- Vardiman, J.W.; Thiele, J.; Arber, D.A.; Brunning, R.D.; Borowitz, M.J.; Porwit, A.; Harris, N.L.; Le Beau, M.M.; Hellstrom-Lindberg, E.; Tefferi, A.; et al. The 2008 revision of the World Health Organization (WHO) classification of myeloid neoplasms and acute leukemia: Rationale and important changes. Blood 2009, 114, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.P.; Cai, J.; Ma, S.Y.; Fang, Y.; Huang, H.Q.; Lin, T.Y.; Rao, H.L.; Li, M.; Xia, Z.J.; Kang, T.B.; et al. BRD2 induces drug resistance through activation of the RasGRP1/Ras/ERK signaling pathway in adult T-cell lymphoblastic lymphoma. Cancer Commun. 2020, 40, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.C.; Stang, S.L.; Zheng, Y.; Dower, N.A.; Brenner, S.E.; Baryza, J.L.; Wender, P.A. Synthetic bryostatin analogues activate the RasGRP1 signaling pathway. J. Med. Chem. 2004, 47, 6638–6644. [Google Scholar] [CrossRef]
- Stang, S.L.; Lopez-Campistrous, A.; Song, X.; Dower, N.A.; Blumberg, P.M.; Wender, P.A.; Stone, J.C. A proapoptotic signaling pathway involving RasGRP, Erk, and Bim in B cells. Exp. Hematol. 2009, 37, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Leo, I.R.; Aswad, L.; Stahl, M.; Kunold, E.; Post, F.; Erkers, T.; Struyf, N.; Mermelekas, G.; Joshi, R.N.; Gracia-Villacampa, E.; et al. Integrative multi-omics and drug response profiling of childhood acute lymphoblastic leukemia cell lines. Nat. Commun. 2022, 13, 1691. [Google Scholar] [CrossRef]
- Oki-Idouchi, C.E.; Lorenzo, P.S. Transgenic overexpression of RasGRP1 in mouse epidermis results in spontaneous tumors of the skin. Cancer Res. 2007, 67, 276–280. [Google Scholar] [CrossRef]
- Diez, F.R.; Garrido, A.A.; Sharma, A.; Luke, C.T.; Stone, J.C.; Dower, N.A.; Cline, J.M.; Lorenzo, P.S. RasGRP1 transgenic mice develop cutaneous squamous cell carcinomas in response to skin wounding: Potential role of granulocyte colony-stimulating factor. Am. J. Pathol. 2009, 175, 392–399. [Google Scholar] [CrossRef]
- Obermueller, E.; Vosseler, S.; Fusenig, N.E.; Mueller, M.M. Cooperative autocrine and paracrine functions of granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor in the progression of skin carcinoma cells. Cancer Res. 2004, 64, 7801–7812. [Google Scholar] [CrossRef]
- Luke, C.T.; Oki-Idouchi, C.E.; Cline, J.M.; Lorenzo, P.S. RasGRP1 overexpression in the epidermis of transgenic mice contributes to tumor progression during multistage skin carcinogenesis. Cancer Res. 2007, 67, 10190–10197. [Google Scholar] [CrossRef]
- Sharma, A.; Fonseca, L.L.; Rajani, C.; Yanagida, J.K.; Endo, Y.; Cline, J.M.; Stone, J.C.; Ji, J.; Ramos, J.W.; Lorenzo, P.S. Targeted deletion of RasGRP1 impairs skin tumorigenesis. Carcinogenesis 2014, 35, 1084–1091. [Google Scholar] [CrossRef]
- Sharma, A.; Luke, C.T.; Dower, N.A.; Stone, J.C.; Lorenzo, P.S. RasGRP1 is essential for ras activation by the tumor promoter 12-O-tetradecanoylphorbol-13-acetate in epidermal keratinocytes. J. Biol. Chem. 2010, 285, 15724–15730. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, L.L.; Yang, W.S.; Geerts, D.; Turkson, J.; Ji, J.; Ramos, J.W. RasGRP1 induces autophagy and transformation-associated changes in primary human keratinocytes. Transl. Oncol. 2021, 14, 100880. [Google Scholar] [CrossRef] [PubMed]
- Gbenedio, O.M.; Bonnans, C.; Grun, D.; Wang, C.Y.; Hatch, A.J.; Mahoney, M.R.; Barras, D.; Matli, M.; Miao, Y.; Garcia, K.C.; et al. RasGRP1 is a potential biomarker to stratify anti-EGFR therapy response in colorectal cancer. JCI Insight 2019, 5, 127552. [Google Scholar] [CrossRef] [PubMed]
- Depeille, P.; Henricks, L.M.; van de Ven, R.A.; Lemmens, E.; Wang, C.Y.; Matli, M.; Werb, Z.; Haigis, K.M.; Donner, D.; Warren, R.; et al. RasGRP1 opposes proliferative EGFR-SOS1-Ras signals and restricts intestinal epithelial cell growth. Nat. Cell Biol. 2015, 17, 804–815. [Google Scholar] [CrossRef]
- Zhang, X.; Zhuang, H.; Han, F.; Shao, X.; Liu, Y.; Ma, X.; Wang, Z.; Qiang, Z.; Li, Y. Sp1-regulated transcription of RasGRP1 promotes hepatocellular carcinoma (HCC) proliferation. Liver Int. 2018, 38, 2006–2017. [Google Scholar] [CrossRef]
- Moon, H.; Ro, S.W. MAPK/ERK Signaling Pathway in Hepatocellular Carcinoma. Cancers 2021, 13, 3026. [Google Scholar] [CrossRef]
- Wang, S.; Beeghly-Fadiel, A.; Cai, Q.; Cai, H.; Guo, X.; Shi, L.; Wu, J.; Ye, F.; Qiu, Q.; Zheng, Y.; et al. Gene expression in triple-negative breast cancer in relation to survival. Breast Cancer Res. Treat. 2018, 171, 199–207. [Google Scholar] [CrossRef]
- Chou, C.W.; Huang, Y.M.; Chang, Y.J.; Huang, C.Y.; Hung, C.S. Identified the novel resistant biomarkers for taxane-based therapy for triple-negative breast cancer. Int. J. Med. Sci. 2021, 18, 2521–2531. [Google Scholar] [CrossRef]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337.e310. [Google Scholar] [CrossRef]
- Latour, S.; Fischer, A. Signaling pathways involved in the T-cell-mediated immunity against Epstein-Barr virus: Lessons from genetic diseases. Immunol. Rev. 2019, 291, 174–189. [Google Scholar] [CrossRef]
- Rchiad, Z.; Haidar, M.; Ansari, H.R.; Tajeri, S.; Mfarrej, S.; Ben Rached, F.; Kaushik, A.; Langsley, G.; Pain, A. Novel tumour suppressor roles for GZMA and RASGRP1 in Theileria annulata-transformed macrophages and human B lymphoma cells. Cell. Microbiol. 2020, 22, e13255. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, A.; Di Maio, A.; Torretta, S.; Garofalo, M.; Giorgelli, V.; Masellis, R.; Nuzzo, T.; Errico, F.; Bertolino, A.; Subramaniam, S.; et al. Abnormal RasGRP1 Expression in the Post-Mortem Brain and Blood Serum of Schizophrenia Patients. Biomolecules 2022, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Kim, S.Y.; Gil, J.E.; Byun, J.S.; Cha, D.W.; Ku, B.; Lee, W.; Kim, W.K.; Oh, K.J.; Lee, E.W.; et al. Nurr1 performs its anti-inflammatory function by regulating RasGRP1 expression in neuro-inflammation. Sci. Rep. 2020, 10, 10755. [Google Scholar] [CrossRef] [PubMed]
- Molineros, J.E.; Singh, B.; Terao, C.; Okada, Y.; Kaplan, J.; McDaniel, B.; Akizuki, S.; Sun, C.; Webb, C.F.; Looger, L.L.; et al. Mechanistic Characterization of RASGRP1 Variants Identifies an hnRNP-K-Regulated Transcriptional Enhancer Contributing to SLE Susceptibility. Front. Immunol. 2019, 10, 1066. [Google Scholar] [CrossRef] [PubMed]
- Eshraghi, M.; Ramirez-Jarquin, U.N.; Shahani, N.; Nuzzo, T.; De Rosa, A.; Swarnkar, S.; Galli, N.; Rivera, O.; Tsaprailis, G.; Scharager-Tapia, C.; et al. RasGRP1 is a causal factor in the development of l-DOPA-induced dyskinesia in Parkinson’s disease. Sci. Adv. 2020, 6, eaaz7001. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, M.; Yu, M.; Shen, J.; Zhou, J.; Hu, J.; Zhou, Y.; Zhang, W. RasGRP1 is a target for VEGF to induce angiogenesis and involved in the endothelial-protective effects of metformin under high glucose in HUVECs. IUBMB Life 2019, 71, 1391–1400. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, T.C.; Rodrigues, G.O.L.; Winer, H.; Hixon, J.A.; Li, W.; Tarasova, N.I.; Durum, S.K. A Focused Review of Ras Guanine Nucleotide-Releasing Protein 1 in Immune Cells and Cancer. Int. J. Mol. Sci. 2023, 24, 1652. https://doi.org/10.3390/ijms24021652
Hsu TC, Rodrigues GOL, Winer H, Hixon JA, Li W, Tarasova NI, Durum SK. A Focused Review of Ras Guanine Nucleotide-Releasing Protein 1 in Immune Cells and Cancer. International Journal of Molecular Sciences. 2023; 24(2):1652. https://doi.org/10.3390/ijms24021652
Chicago/Turabian StyleHsu, Tu Chun, Gisele O. L. Rodrigues, Hila Winer, Julie A. Hixon, Wenqing Li, Nadya I. Tarasova, and Scott K. Durum. 2023. "A Focused Review of Ras Guanine Nucleotide-Releasing Protein 1 in Immune Cells and Cancer" International Journal of Molecular Sciences 24, no. 2: 1652. https://doi.org/10.3390/ijms24021652
APA StyleHsu, T. C., Rodrigues, G. O. L., Winer, H., Hixon, J. A., Li, W., Tarasova, N. I., & Durum, S. K. (2023). A Focused Review of Ras Guanine Nucleotide-Releasing Protein 1 in Immune Cells and Cancer. International Journal of Molecular Sciences, 24(2), 1652. https://doi.org/10.3390/ijms24021652