
Comprehensive Transcriptomic Analysis of VISTA in Acute Myeloid Leukemia: Insights into Its Prognostic Value
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
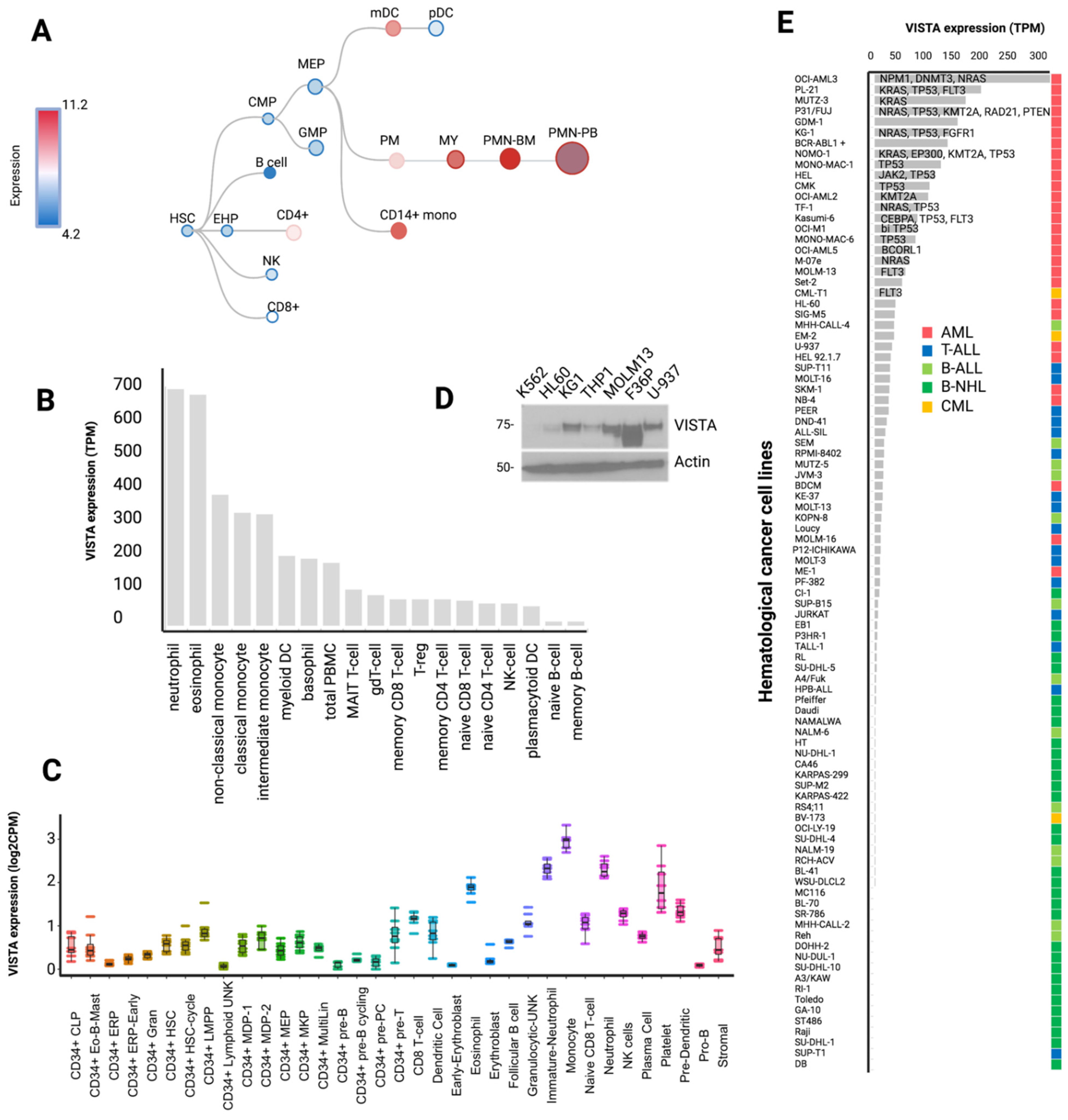
2.1. Cartography of VISTA Expression across Normal Hematopoiesis and Hematological Cancer Cell Lines
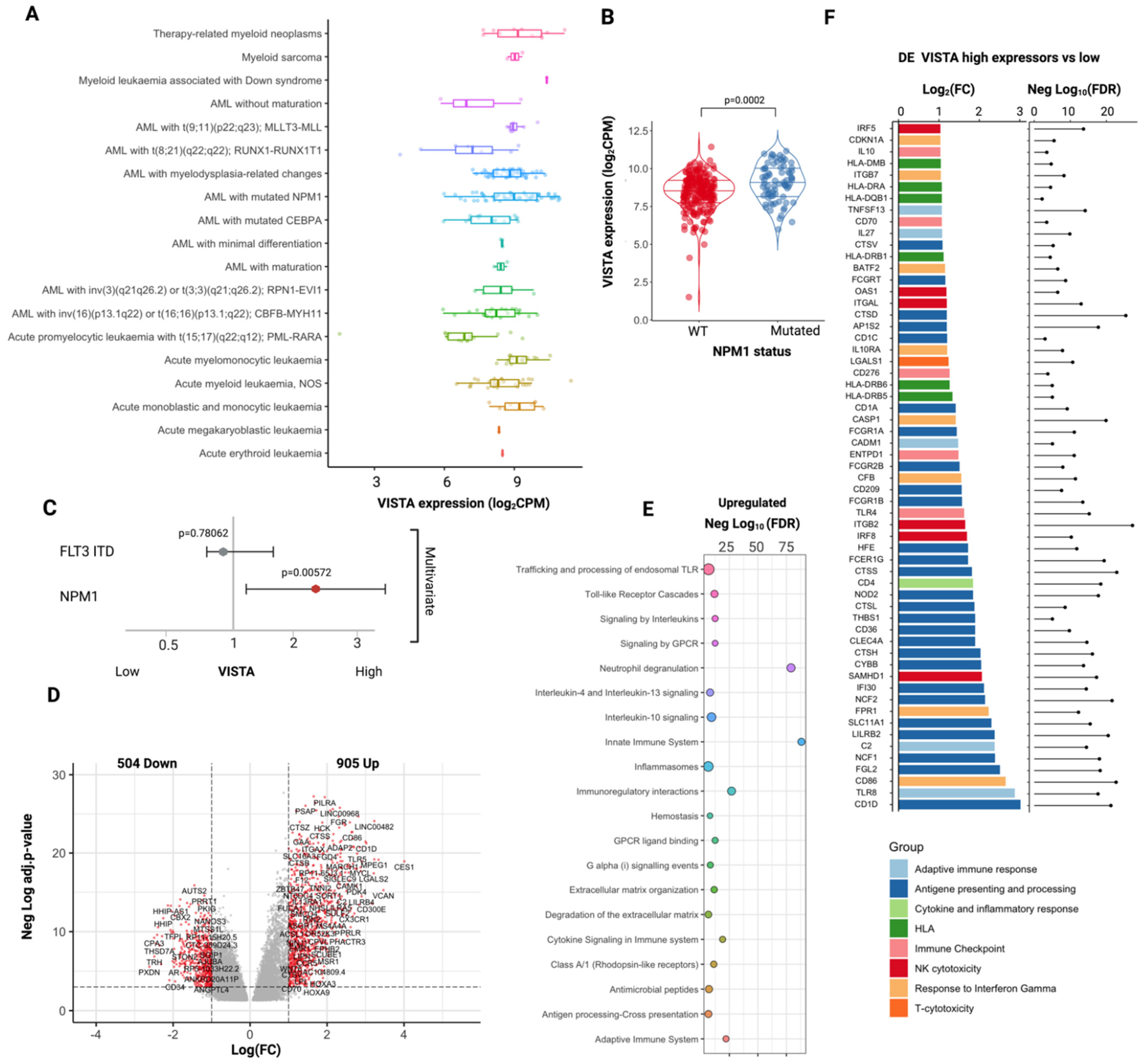
2.2. VISTA Expression Profile in AML
2.3. VISTA Expression Correlates with a Lower Mutational Burden and with a Specific Myeloid Landscape
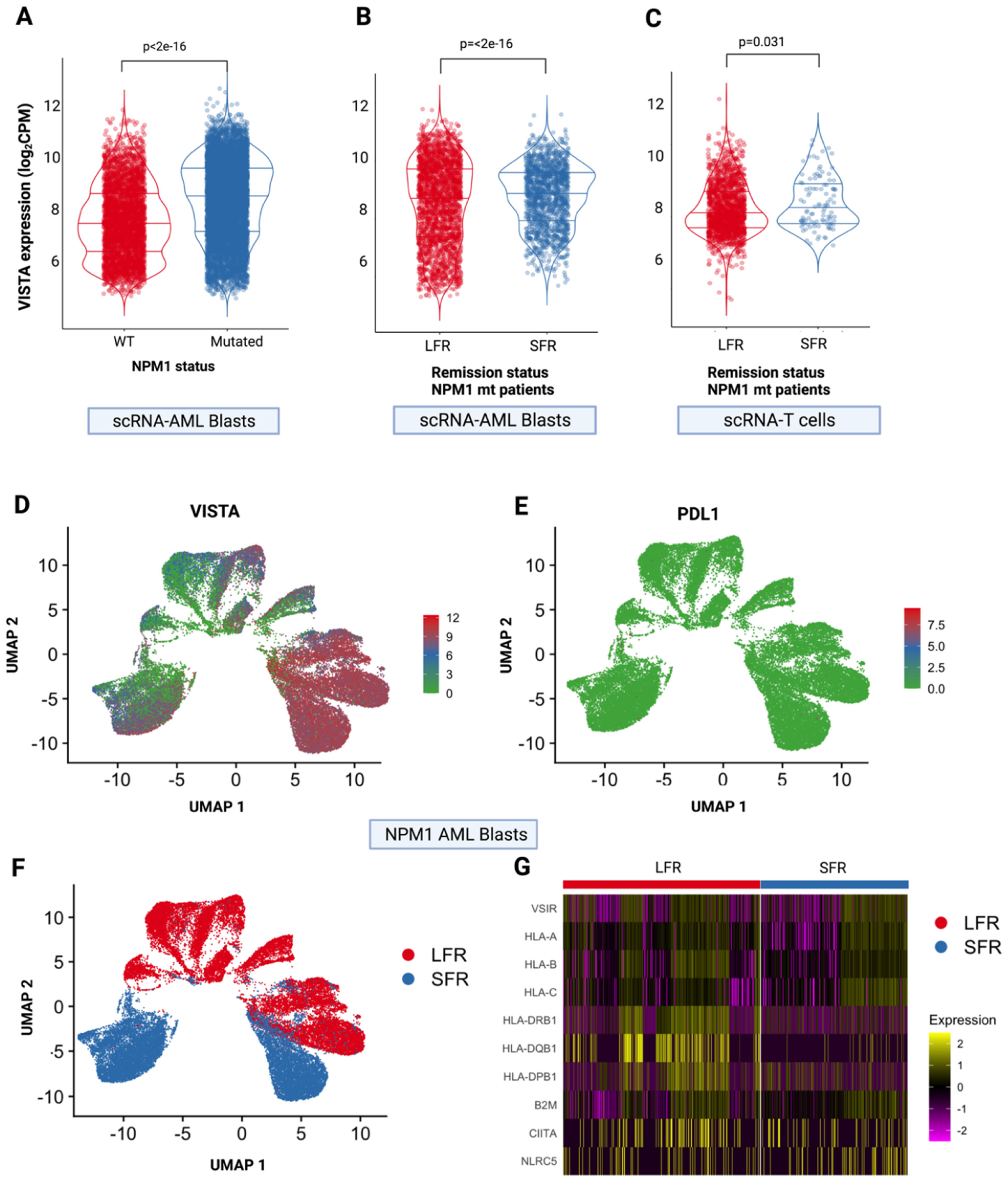
2.4. Leukemic T-Cell VISTA Interactions in NPM1 Mutated AML
3. Discussion
4. Methods and Materials
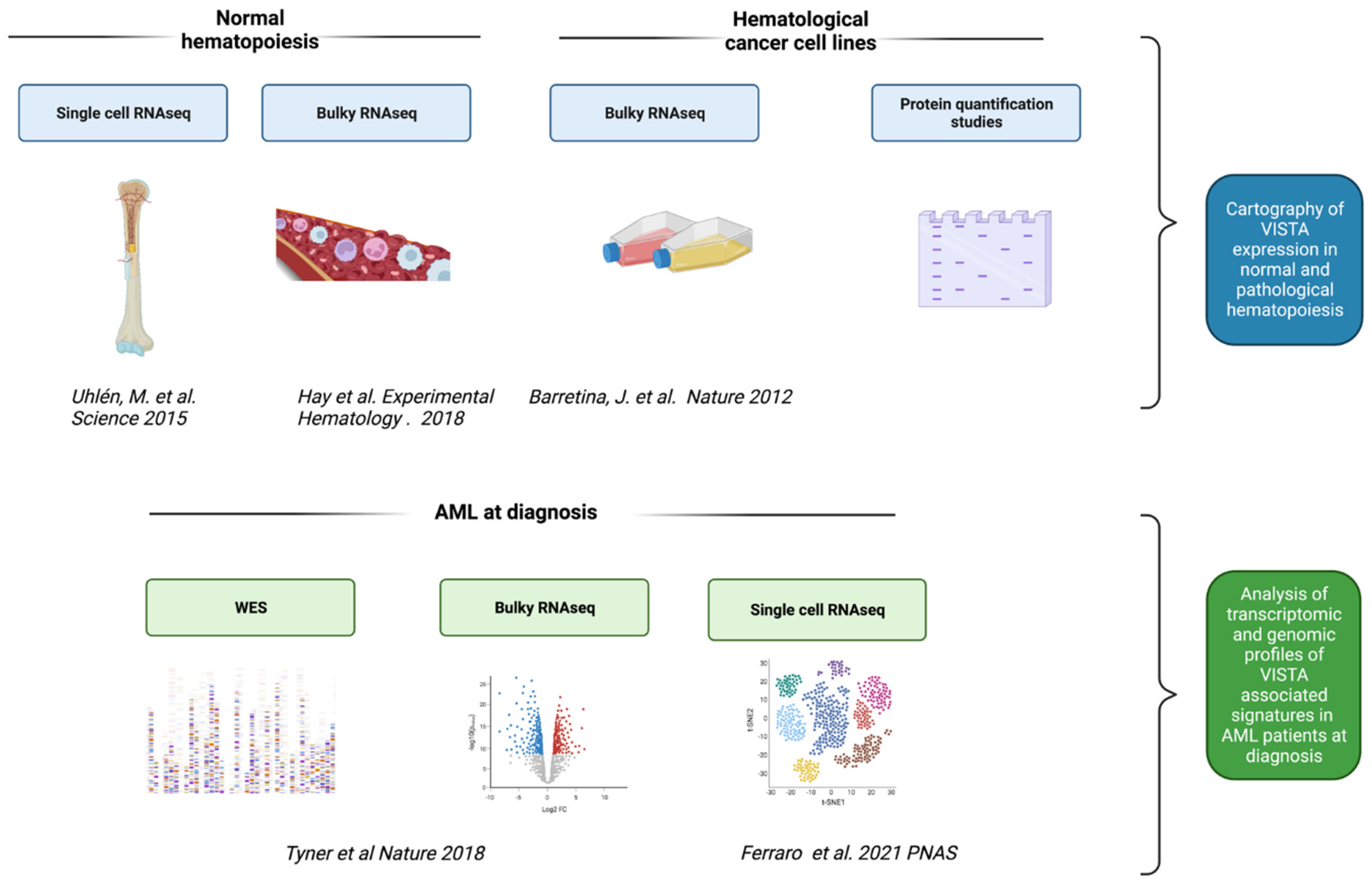
4.1. Study Overview
4.2. Transcriptomic and Differential Expression Analysis
4.3. Single-Cell RNA Sequencing and Analysis
4.4. Genomic Analysis
4.5. Western Blot Study
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, P.; Allison, J.P. The Future of Immune Checkpoint Therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef]
- Fares, C.M.; Van Allen, E.M.; Drake, C.G.; Allison, J.P.; Hu-Lieskovan, S. Mechanisms of Resistance to Immune Checkpoint Blockade: Why Does Checkpoint Inhibitor Immunotherapy Not Work for All Patients? Am. Soc. Clin. Oncol. Educ. Book 2019, 39, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Havel, J.J.; Chowell, D.; Chan, T.A. The Evolving Landscape of Biomarkers for Checkpoint Inhibitor Immunotherapy. Nat. Rev. Cancer 2019, 19, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Llobell, M.; Peleteiro Raíndo, A.; Climent Medina, J.; Gómez Centurión, I.; Mosquera Orgueira, A. Immune Checkpoint Inhibitors in Acute Myeloid Leukemia: A Meta-Analysis. Front. Oncol. 2022, 12, 882531. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, F.; Miller, C.A.; Christensen, K.A.; Helton, N.M.; O’Laughlin, M.; Fronick, C.C.; Fulton, R.S.; Kohlschmidt, J.; Eisfeld, A.-K.; Bloomfield, C.D.; et al. Immunosuppression and Outcomes in Adult Patients with de Novo Acute Myeloid Leukemia with Normal Karyotypes. Proc. Natl. Acad. Sci. USA 2021, 118, e2116427118. [Google Scholar] [CrossRef]
- Bewersdorf, J.P.; Stahl, M.; Zeidan, A.M. Immune Checkpoint-Based Therapy in Myeloid Malignancies: A Promise yet to Be Fulfilled. Expert Rev. Anticancer Ther. 2019, 19, 393–404. [Google Scholar] [CrossRef]
- Xu, W.; Hiếu, T.; Malarkannan, S.; Wang, L. The Structure, Expression, and Multifaceted Role of Immune-Checkpoint Protein VISTA as a Critical Regulator of Anti-Tumor Immunity, Autoimmunity, and Inflammation. Cell Mol. Immunol. 2018, 15, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Rubinstein, R.; Lines, J.L.; Wasiuk, A.; Ahonen, C.; Guo, Y.; Lu, L.-F.; Gondek, D.; Wang, Y.; Fava, R.A.; et al. VISTA, a Novel Mouse Ig Superfamily Ligand That Negatively Regulates T Cell Responses. J. Exp. Med. 2011, 208, 577–592. [Google Scholar] [CrossRef]
- Lines, J.L.; Sempere, L.F.; Broughton, T.; Wang, L.; Noelle, R. VISTA Is a Novel Broad-Spectrum Negative Checkpoint Regulator for Cancer Immunotherapy. Cancer Immunol. Res. 2014, 2, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.J.; Su, L.J.; Pinckney, J.; Critton, D.; Boyer, E.; Krishnakumar, A.; Corbett, M.; Rankin, A.L.; Dibella, R.; Campbell, L.; et al. VISTA Is an Acidic PH-Selective Ligand for PSGL-1. Nature 2019, 574, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Le Mercier, I.; Chen, W.; Lines, J.L.; Day, M.; Li, J.; Sergent, P.; Noelle, R.J.; Wang, L. VISTA Regulates the Development of Protective Antitumor Immunity. Cancer Res. 2014, 74, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yuan, Y.; Chen, W.; Putra, J.; Suriawinata, A.A.; Schenk, A.D.; Miller, H.E.; Guleria, I.; Barth, R.J.; Huang, Y.H.; et al. Immune-Checkpoint Proteins VISTA and PD-1 Nonredundantly Regulate Murine T-Cell Responses. Proc. Natl. Acad. Sci. USA 2015, 112, 6682–6687. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jia, B.; Claxton, D.F.; Ehmann, W.C.; Rybka, W.B.; Mineishi, S.; Naik, S.; Khawaja, M.R.; Sivik, J.; Han, J.; et al. VISTA Is Highly Expressed on MDSCs and Mediates an Inhibition of T Cell Response in Patients with AML. OncoImmunology 2018, 7, e1469594. [Google Scholar] [CrossRef]
- Xu, W.; Dong, J.; Zheng, Y.; Zhou, J.; Yuan, Y.; Ta, H.M.; Miller, H.E.; Olson, M.; Rajasekaran, K.; Ernstoff, M.S.; et al. Immune-Checkpoint Protein VISTA Regulates Antitumor Immunity by Controlling Myeloid Cell–Mediated Inflammation and Immunosuppression. Cancer Immunol. Res. 2019, 7, 1497–1510. [Google Scholar] [CrossRef]
- Dufva, O.; Pölönen, P.; Brück, O.; Keränen, M.A.I.; Klievink, J.; Mehtonen, J.; Huuhtanen, J.; Kumar, A.; Malani, D.; Siitonen, S.; et al. Immunogenomic Landscape of Hematological Malignancies. Cancer Cell 2020, 38, 380–399.e13. [Google Scholar] [CrossRef] [PubMed]
- Hay, S.B.; Ferchen, K.; Chetal, K.; Grimes, H.L.; Salomonis, N. The Human Cell Atlas Bone Marrow Single-Cell Interactive Web Portal. Exp. Hematol. 2018, 68, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehár, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia Enables Predictive Modelling of Anticancer Drug Sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef]
- Tyner, J.W.; Tognon, C.E.; Bottomly, D.; Wilmot, B.; Kurtz, S.E.; Savage, S.L.; Long, N.; Schultz, A.R.; Traer, E.; Abel, M.; et al. Functional Genomic Landscape of Acute Myeloid Leukaemia. Nature 2018, 562, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Gurnari, C.; Pagliuca, S.; Visconte, V. Deciphering the Therapeutic Resistance in Acute Myeloid Leukemia. Int. J. Mol. Sci. 2020, 21, 8505. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Döhner, H.; Estey, E.; Grimwade, D.; Amadori, S.; Appelbaum, F.R.; Büchner, T.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Larson, R.A.; et al. Diagnosis and Management of AML in Adults: 2017 ELN Recommendations from an International Expert Panel. Blood 2017, 129, 424–447. [Google Scholar] [CrossRef]
- Novershtern, N.; Subramanian, A.; Lawton, L.N.; Mak, R.H.; Haining, W.N.; McConkey, M.E.; Habib, N.; Yosef, N.; Chang, C.Y.; Shay, T.; et al. Densely Interconnected Transcriptional Circuits Control Cell States in Human Hematopoiesis. Cell 2011, 144, 296–309. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M.; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated Analysis of Multimodal Single-Cell Data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagliuca, S.; Gurnari, C.; Zhang, K.; Kewan, T.; Bahaj, W.; Mori, M.; Nautiyal, I.; Rubio, M.T.; Ferraro, F.; Maciejewski, J.P.; et al. Comprehensive Transcriptomic Analysis of VISTA in Acute Myeloid Leukemia: Insights into Its Prognostic Value. Int. J. Mol. Sci. 2022, 23, 14885. https://doi.org/10.3390/ijms232314885
Pagliuca S, Gurnari C, Zhang K, Kewan T, Bahaj W, Mori M, Nautiyal I, Rubio MT, Ferraro F, Maciejewski JP, et al. Comprehensive Transcriptomic Analysis of VISTA in Acute Myeloid Leukemia: Insights into Its Prognostic Value. International Journal of Molecular Sciences. 2022; 23(23):14885. https://doi.org/10.3390/ijms232314885
Chicago/Turabian StylePagliuca, Simona, Carmelo Gurnari, Keman Zhang, Tariq Kewan, Waled Bahaj, Minako Mori, Ishani Nautiyal, Marie Thérèse Rubio, Francesca Ferraro, Jaroslaw P. Maciejewski, and et al. 2022. "Comprehensive Transcriptomic Analysis of VISTA in Acute Myeloid Leukemia: Insights into Its Prognostic Value" International Journal of Molecular Sciences 23, no. 23: 14885. https://doi.org/10.3390/ijms232314885
APA StylePagliuca, S., Gurnari, C., Zhang, K., Kewan, T., Bahaj, W., Mori, M., Nautiyal, I., Rubio, M. T., Ferraro, F., Maciejewski, J. P., Wang, L., & Visconte, V. (2022). Comprehensive Transcriptomic Analysis of VISTA in Acute Myeloid Leukemia: Insights into Its Prognostic Value. International Journal of Molecular Sciences, 23(23), 14885. https://doi.org/10.3390/ijms232314885