
Structural Basis of the Inhibition of L-Methionine γ-Lyase from Fusobacterium nucleatum

 , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Results
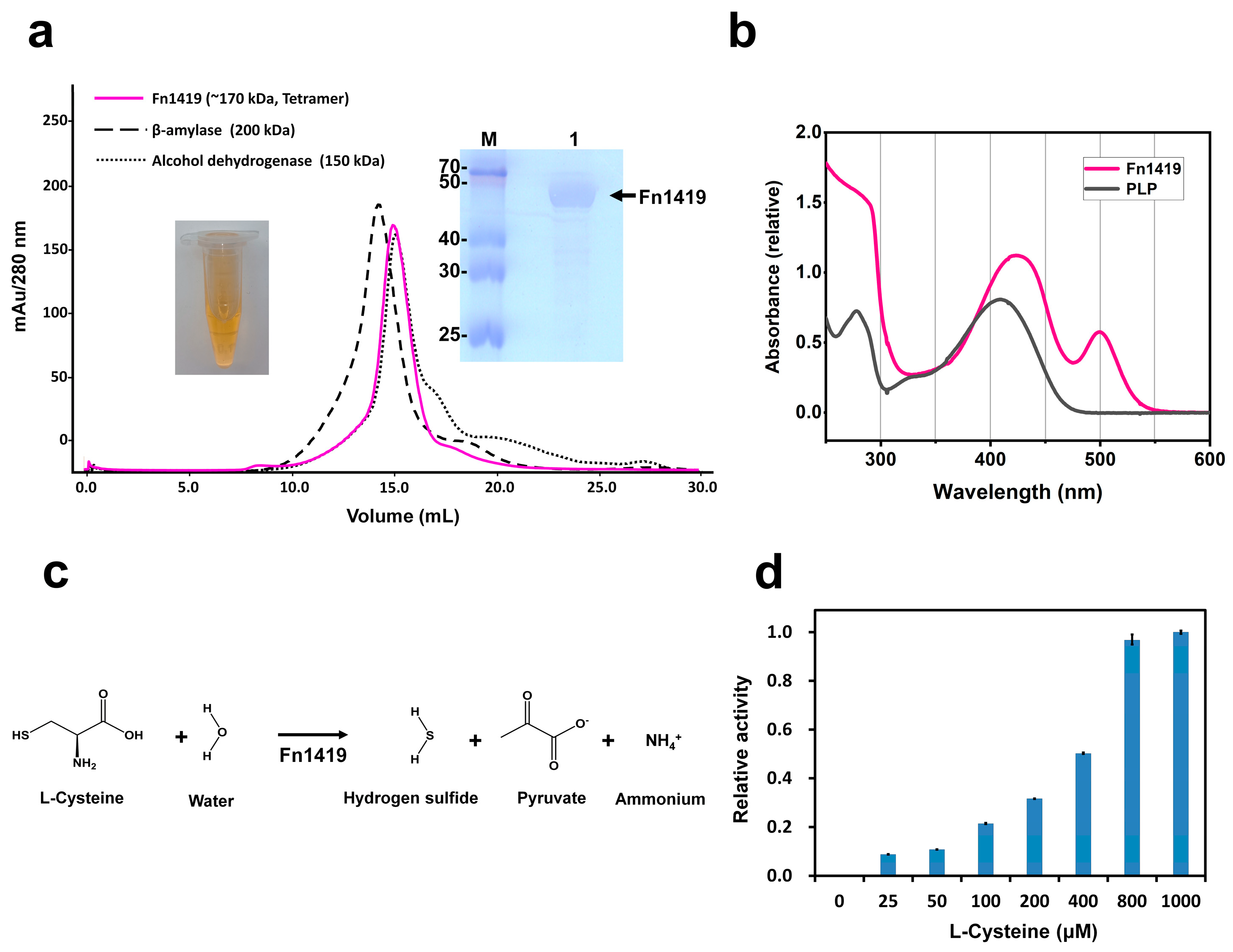
2.1. Characterization of Fn1419
2.2. Crystal Structure of Fn1419
2.3. Substrate- and Cofactor-Binding Sites of Fn1419
2.4. Screening of Leading Compounds for Fn1419 Inhibition
2.5. Identification of Fn1419-Binding Site for Trihydroxybenzene-Based Lead Compounds
3. Discussion
4. Materials and Methods
4.1. Protein Preparation
4.2. Analytical Size-Exclusion Chromatography
4.3. PLP Binding Analysis
4.4. Enzyme Assays
4.5. Crystallization, Data Collection, and Structure Determination
4.6. Inhibitor Screening Assay
4.7. IC50 Determination
4.8. Molecular Docking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brook, I. Spectrum and treatment of anaerobic infections. J. Infect. Chemother. 2016, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hecht, D.W. Prevalence of antibiotic resistance in anaerobic bacteria: Worrisome developments. Clin. Infect. Dis. 2004, 39, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, X.; Huang, Y.; Liao, B.; Cheng, L.; Ren, B. Reactive Oxygen Species in Pathogen Clearance: The Killing Mechanisms, the Adaption Response, and the Side Effects. Front. Microbiol. 2020, 11, 622534. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Ying, J.; Xiang, L.; Zhang, C. The biologic effect of hydrogen sulfide and its function in various diseases. Medicine 2018, 97, e13065. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, R.; Jeong, N.Y.; Jung, J. Protective effect of hydrogen sulfide on oxidative stress-induced neurodegenerative diseases. Neural Regen. Res. 2020, 15, 232–241. [Google Scholar]
- Shatalin, K.; Nuthanakanti, A.; Kaushik, A.; Shishov, D.; Peselis, A.; Shamovsky, I.; Pani, B.; Lechpammer, M.; Vasilyev, N.; Shatalina, E.; et al. Inhibitors of bacterial H2S biogenesis targeting antibiotic resistance and tolerance. Science 2021, 372, 1169–1175. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef]
- Sun, C.H.; Li, B.B.; Wang, B.; Zhao, J.; Zhang, X.Y.; Li, T.T.; Li, W.B.; Tang, D.; Qiu, M.J.; Wang, X.C.; et al. The role of Fusobacterium nucleatum in colorectal cancer: From carcinogenesis to clinical management. Chronic Dis. Transl. Med. 2019, 5, 178–187. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Kim, E. Cysteine-derived hydrogen sulfide and gut health: A matter of endogenous or bacterial origin. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 68–75. [Google Scholar] [CrossRef]
- Kabil, O.; Vitvitsky, V.; Xie, P.; Banerjee, R. The quantitative significance of the transsulfuration enzymes for H2S production in murine tissues. Antioxid. Redox Signal. 2011, 15, 363–372. [Google Scholar] [CrossRef]
- Paul, B.D.; Snyder, S.H. H2S signalling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Bio. 2012, 13, 499–507. [Google Scholar] [CrossRef]
- Zhao, Y.; Cerda, M.M.; Pluth, M.D. Fluorogenic hydrogen sulfide (H2S) donors based on sulfenyl thiocarbonates enable H2S tracking and quantification. Chem. Sci. 2019, 10, 1873–1878. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Enzymology of H2S biogenesis, decay and signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Braccia, D.J.; Jiang, X.; Pop, M.; Hall, A.B. The Capacity to Produce Hydrogen Sulfide (H2S) via Cysteine Degradation Is Ubiquitous in the Human Gut Microbiome. Front. Microbiol. 2021, 12, 705583. [Google Scholar] [CrossRef]
- Suwabe, K.; Yoshida, Y.; Nagano, K.; Yoshimura, F. Identification of an L-methionine gamma-lyase involved in the production of hydrogen sulfide from L-cysteine in Fusobacterium nucleatum subsp. nucleatum ATCC 25586. Microbiology 2011, 157 Pt 10, 2992–3000. [Google Scholar] [CrossRef]
- Mothersole, R.G.; Wolthers, K.R. Structural and Kinetic Insight into the Biosynthesis of H2S and l-Lanthionine from l-Cysteine by a Pyridoxal l-Phosphate-Dependent Enzyme from Fusobacterium nucleatum. Biochemistry 2019, 58, 3592–3603. [Google Scholar] [CrossRef]
- Sato, D.; Yamagata, W.; Harada, S.; Nozaki, T. Kinetic characterization of methionine gamma-lyases from the enteric protozoan parasite Entamoeba histolytica against physiological substrates and trifluoromethionine, a promising lead compound against amoebiasis. FEBS J. 2008, 275, 548–560. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, Y.; Zhou, H.; Luo, H.B.; Zhan, C.G. Catalytic Roles of Coenzyme Pyridoxal-5′-phosphate (PLP) in PLP-dependent Enzymes: Reaction Pathway for Methionine-gamma-lyase-catalyzed L-methionine Depletion. ACS Catal. 2020, 10, 2198–2210. [Google Scholar] [CrossRef]
- Toney, M.D. Reaction specificity in pyridoxal phosphate enzymes. Arch. Biochem. Biophys. 2005, 433, 279–287. [Google Scholar] [CrossRef]
- He, S.; Chen, Y.; Wang, L.; Bai, X.; Bu, T.; Zhang, J.; Lu, M.; Ha, N.C.; Quan, C.; Nam, K.H.; et al. Structural and Functional Analysis of the Pyridoxal Phosphate Homeostasis Protein YggS from Fusobacterium nucleatum. Molecules 2022, 27, 4781. [Google Scholar] [CrossRef] [PubMed]
- Sato, D.; Shiba, T.; Yunoto, S.; Furutani, K.; Fukumoto, M.; Kudou, D.; Tamura, T.; Inagaki, K.; Harada, S. Structural and mechanistic insights into homocysteine degradation by a mutant of methionine gamma-lyase based on substrate-assisted catalysis. Protein Sci. 2017, 26, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Revtovich, S.V.; Faleev, N.G.; Morozova, E.A.; Anufrieva, N.V.; Nikulin, A.D.; Demidkina, T.V. Crystal structure of the external aldimine of Citrobacter freundii methionine gamma-lyase with glycine provides insight in mechanisms of two stages of physiological reaction and isotope exchange of alpha- and beta-protons of competitive inhibitors. Biochimie 2014, 101, 161–167. [Google Scholar] [CrossRef]
- Revtovich, S.; Anufrieva, N.; Morozova, E.; Kulikova, V.; Nikulin, A.; Demidkina, T. Structure of methionine gamma-lyase from Clostridium sporogenes. Acta Crystallogr. F Struct. Biol. Commun. 2016, 72 Pt 1, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences, T.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.; Seregina, T.; Nagornykh, M.; Luhachack, L.G.; Korolkova, N.; Lopes, L.E.; Kotova, V.; Zavilgelsky, G.; Shakulov, R.; Shatalin, K.; et al. Mechanism of H2S-mediated protection against oxidative stress in Escherichia coli. Proc. Natl. Acad. Sci. USA 2017, 114, 6022–6027. [Google Scholar] [CrossRef]
- Toliver-Kinsky, T.; Cui, W.; Toro, G.; Lee, S.J.; Shatalin, K.; Nudler, E.; Szabo, C. H2S, a Bacterial Defense Mechanism against the Host Immune Response. Infect. Immun. 2019, 87, e00272-18. [Google Scholar] [CrossRef]
- Mendes, S.S.; Miranda, V.; Saraiva, L.M. Hydrogen Sulfide and Carbon Monoxide Tolerance in Bacteria. Antioxidants 2021, 10, 729. [Google Scholar] [CrossRef]
- Yoshida, Y.; Suwabe, K.; Nagano, K.; Kezuka, Y.; Kato, H.; Yoshimura, F. Identification and enzymic analysis of a novel protein associated with production of hydrogen sulfide and L-serine from L-cysteine in Fusobacterium nucleatum subsp. nucleatum ATCC 25586. Microbiology 2011, 157 Pt 7, 2164–2171. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef]
- Park, S.; Imlay, J.A. High levels of intracellular cysteine promote oxidative DNA damage by driving the fenton reaction. J. Bacteriol. 2003, 185, 1942–1950. [Google Scholar] [CrossRef]
- Somerville, G.A.; Proctor, R.A. At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci. Microbiol. Mol. Biol. Rev. 2009, 73, 233–248. [Google Scholar] [CrossRef]
- Poole, K. Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 2012, 67, 2069–2089. [Google Scholar] [CrossRef]
- Zhang, H.; Caprioli, G.; Hussain, H.; Le, N.P.K.; Farag, M.A.; Xiao, J. A multifaceted review on dihydromyricetin resources, extraction, bioavailability, biotransformation, bioactivities, and food applications with future perspectives to maximize its value. eFood 2021, 2, 164–184. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, L.; Liao, P.; Xiao, Z.; Zhang, F.; Sindaye, D.; Xin, Z.; Tan, C.; Deng, J.; Yin, Y.; et al. Impact of Gallic Acid on Gut Health: Focus on the Gut Microbiome, Immune Response, and Mechanisms of Action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef]
- Rajamanickam, K.; Yang, J.; Sakharkar, M.K. Gallic Acid Potentiates the Antimicrobial Activity of Tulathromycin against Two Key Bovine Respiratory Disease (BRD) Causing-Pathogens. Front. Pharmacol. 2018, 9, 1486. [Google Scholar] [CrossRef]
- Cota, D.; Patil, D. Antibacterial potential of ellagic acid and gallic acid against IBD bacterial isolates and cytotoxicity against colorectal cancer. Nat. Prod. Res. 2022, 15, 1–5. [Google Scholar] [CrossRef]
- Allemandou, F.; Nussberger, J.; Brunner, H.R.; Brakch, N. Rapid Site-Directed Mutagenesis Using Two-PCR-Generated DNA Fragments Reproducing the Plasmid Template. J. Biomed. Biotechnol. 2003, 2003, 202–207. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]
- Evans, P.; McCoy, A. An introduction to molecular replacement. Acta Crystallogr. D Biol. Crystallogr. 2008, 64 Pt 1, 1–10. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Grosse-Kunstleve, R.W.; Hung, L.W.; Ioerger, T.R.; McCoy, A.J.; Moriarty, N.W.; Read, R.J.; Sacchettini, J.C.; Sauter, N.K.; Terwilliger, T.C. PHENIX: Building new software for automated crystallographic structure determination. Acta Crystallogr. D 2002, 58, 1948–1954. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. Use of PYMOL as a communications tool for molecular science. Abstr. Pap. Am. Chem. Soc. 2004, 228, U313–U314. [Google Scholar]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- De Vries, S.J.; van Dijk, M.; Bonvin, A.M.J.J. The HADDOCK web server for data-driven biomolecular docking. Nat. Protoc. 2010, 5, 883–897. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data collection | Fn1419 |
| Beamline | Beamline BL19U1 at SSRF |
| Resolution range (Å) | 42.99–2.50 (2.589–2.5) |
| Space group | P3121 |
| a, b, c (Å) | 95.21, 95.21, 302.25 |
| α, β, γ (o) | 90.00, 90.00, 120.00 |
| Rsym (%) | 6.2 (30.7) |
| Completeness (%) | 99.55 (98.89) |
| Multiplicity | 2.1 (2.1) |
| Average I/σ(I) | 17.27 (3.15) |
| Refinement | |
| Rwork/Rfree (%) | 21.60 (24.61)/27.88 (35.42) |
| Protein residues | 395 |
| Protein | 52.73 |
| Water | 46.92 |
| R.m.s.d. from ideal | |
| RMS (Bond) | 0.009 |
| RMS (Angles) | 1.240 |
| Ramachandran plot (%) | |
| Favored regions | 94.73 |
| Allowed regions | 4.69 |
| Disallowed regions | 0.58 |
| PDB code | 7BQW |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, T.; Lan, J.; Jo, I.; Zhang, J.; Bai, X.; He, S.; Jin, X.; Wang, L.; Jin, Y.; Jin, X.; et al. Structural Basis of the Inhibition of L-Methionine γ-Lyase from Fusobacterium nucleatum. Int. J. Mol. Sci. 2023, 24, 1651. https://doi.org/10.3390/ijms24021651
Bu T, Lan J, Jo I, Zhang J, Bai X, He S, Jin X, Wang L, Jin Y, Jin X, et al. Structural Basis of the Inhibition of L-Methionine γ-Lyase from Fusobacterium nucleatum. International Journal of Molecular Sciences. 2023; 24(2):1651. https://doi.org/10.3390/ijms24021651
Chicago/Turabian StyleBu, Tingting, Jing Lan, Inseong Jo, Jie Zhang, Xue Bai, Shanru He, Xiaoling Jin, Lulu Wang, Yu Jin, Xiaoyu Jin, and et al. 2023. "Structural Basis of the Inhibition of L-Methionine γ-Lyase from Fusobacterium nucleatum" International Journal of Molecular Sciences 24, no. 2: 1651. https://doi.org/10.3390/ijms24021651
APA StyleBu, T., Lan, J., Jo, I., Zhang, J., Bai, X., He, S., Jin, X., Wang, L., Jin, Y., Jin, X., Zhang, L., Piao, H., Ha, N.-C., Quan, C., Nam, K. H., & Xu, Y. (2023). Structural Basis of the Inhibition of L-Methionine γ-Lyase from Fusobacterium nucleatum. International Journal of Molecular Sciences, 24(2), 1651. https://doi.org/10.3390/ijms24021651