
Effect of Antibiotic Exposure on Staphylococcus epidermidis Responsible for Catheter-Related Bacteremia
 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
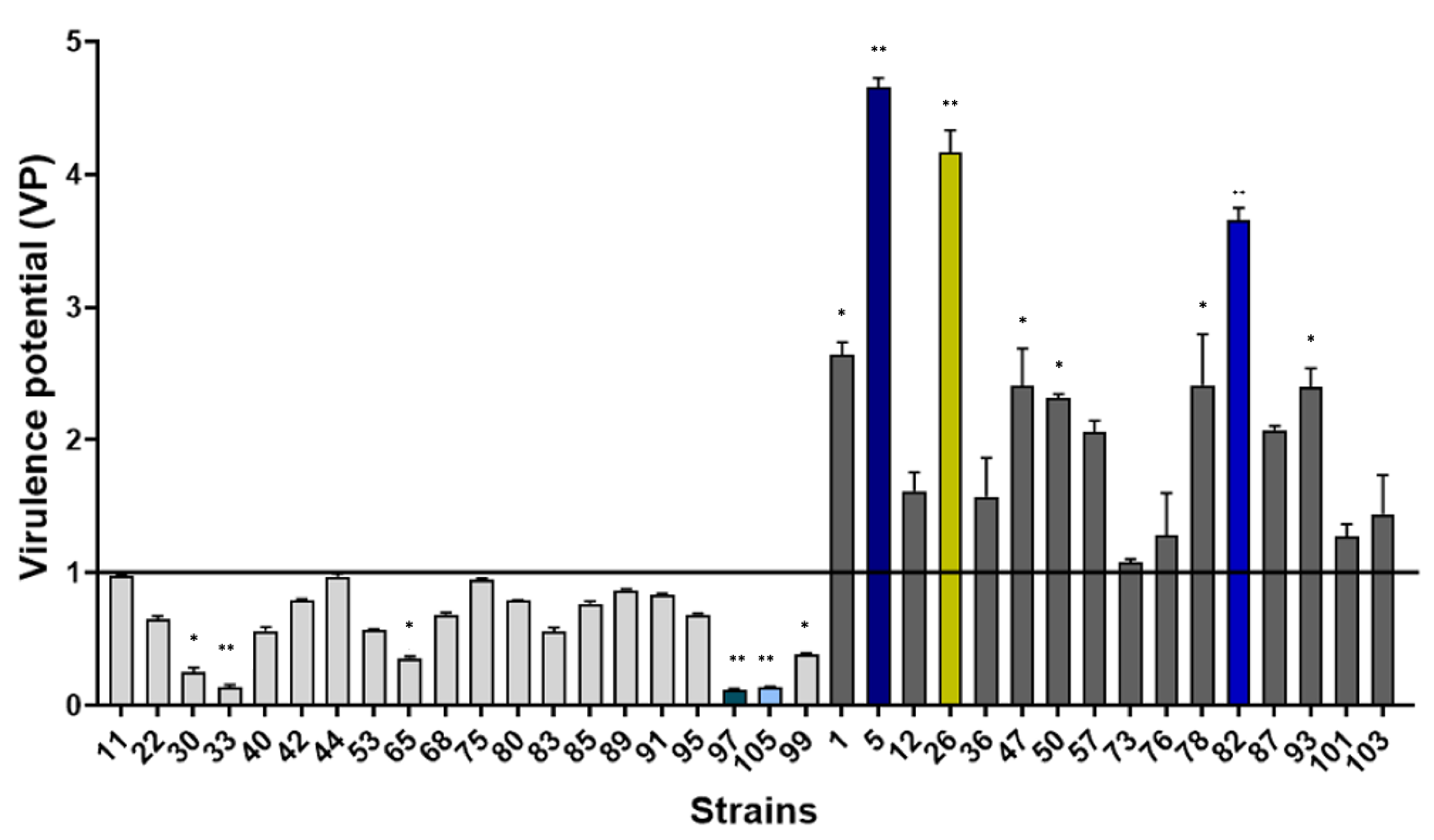
2.1. Potential Virulence of S. epidermidis Isolated from Catheter-Related Bacteremia
2.1.1. Evaluation of Virulence in an In Vivo Caenorhabditis Elegans Model
2.1.2. Evaluation of Biofilm Formation Ability
2.2. Effects of Exposure to Sub-Inhibitory Concentrations of Antibiotics on Virulence and Resistance of S. epidermidis
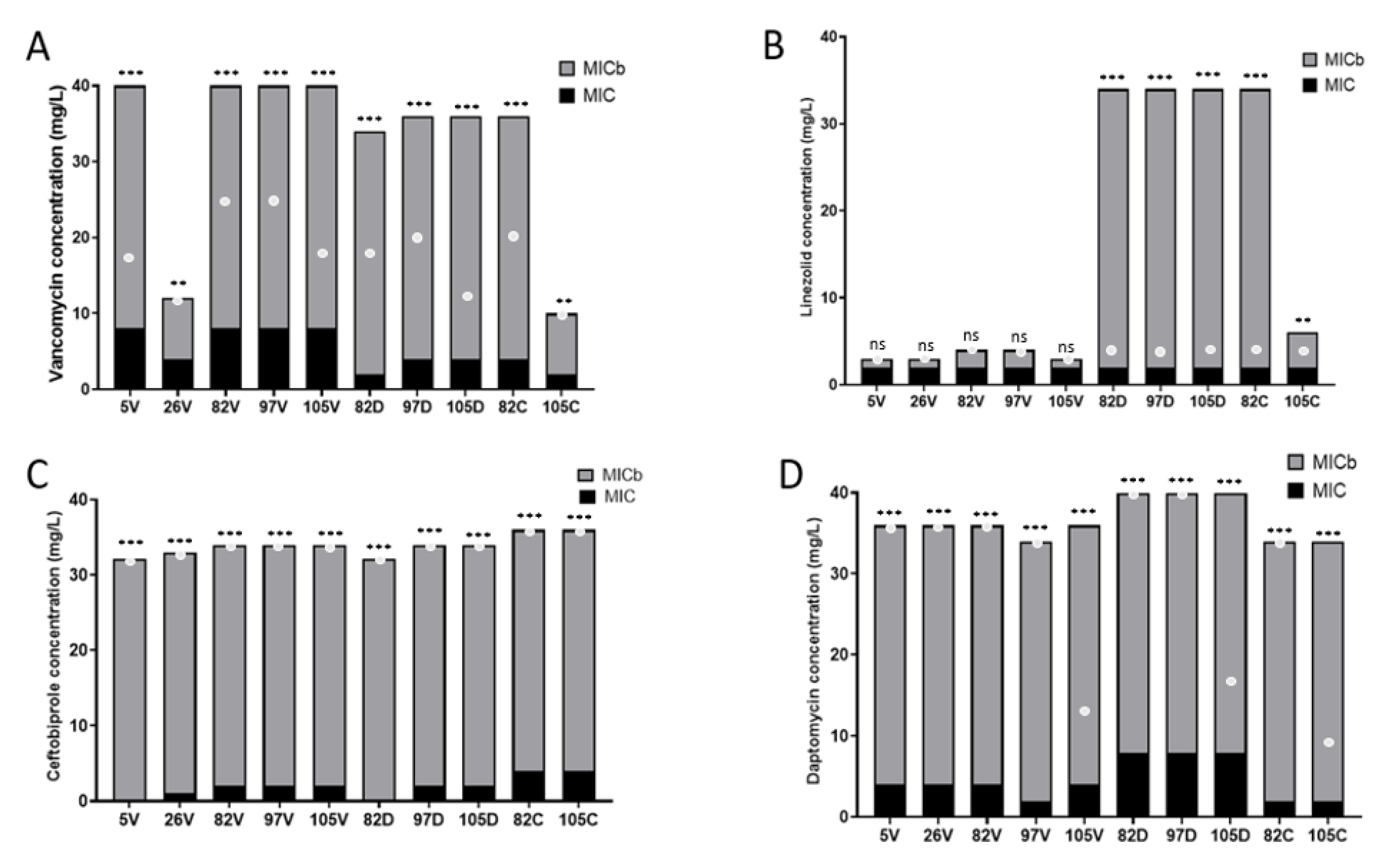
2.2.1. Effect on Antibiotic Susceptibility of S. epidermidis
2.2.2. Effect on Biofilm Formation of S. epidermidis
2.2.3. Effect of Antibiotics on Their Ability to Inhibit S. epidermidis Biofilm Formation
2.2.4. Effect on the Genomes of S. epidermidis after Antibiotic Exposure
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Culture Conditions and Antimicrobial Susceptibility Testing
4.2. Nematode Killing Assay
4.3. Biofilm Formation
4.4. Antibiotic Exposure
4.5. Antibiofilmogram®
4.6. Whole-Genome Sequencing
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costa, S.F.; Miceli, M.H.; Anaissie, E.J. Mucosa or skin as source of coagulase-negative staphylococcal bacteraemia? Lancet Infect. Dis. 2004, 4, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis–the ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Severn, M.M.; Horswill, A.R. Staphylococcus epidermidis and its dual lifestyle in skin health and infection. Nat. Rev. Microbiol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.; Sartelli, M.; McKimm, J.; Abu Bakar, M. Health care-associated infections–an overview. Infect. Drug Resist. 2018, 11, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Annual Epidemiological Report for 2016: Healthcare-Associated Infections in Intensive Care Units. European Centre for Disease Prevention and Control. 2018. Available online: https://ecdc.europa.eu/sites/portal/files/documents/AER_for_2016-HAI_0.pdf (accessed on 12 August 2022).
- Allegranzi, B.; Bagheri Nejad, S.; Combescure, C.; Graafmans, W.; Attar, H.; Donaldson, L.; Pittet, D. Burden of endemic health-care-associated infection in developing countries: Systematic review and meta-analysis. Lancet 2011, 377, 228–241. [Google Scholar] [CrossRef]
- Vincent, J.L. Nosocomial Infections in Adult Intensive-Care Units. Lancet 2003, 361, 2068–2077. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef]
- Zarb, P.; Coignard, B.; Griskeviciene, J.; Muller, A.; Vankerckhoven, V.; Weist, K.; Goossens, M.; Vaerenberg, S.; Hopkins, S.; Catry, B.; et al. The European Centre for Disease Prevention and Control (ECDC) Pilot Point Prevalence Survey of Healthcare-Associated Infections and Antimicrobial Use. Eurosurveillance 2012, 17, 20316. [Google Scholar] [CrossRef]
- Águila-Arcos, S.; Álvarez-Rodríguez, I.; Garaiyurrebaso, O.; Garbisu, C.; Grohmann, E.; Alkorta, I. Biofilm-forming clinical Staphylococcus isolates harbor horizontal transfer and antibiotic resistance genes. Front. Microbiol. 2017, 8, 2018. [Google Scholar] [CrossRef]
- Dengler Haunreiter, V.; Boumasmoud, M.; Häffner, N.; Wipfli, D.; Leimer, N.; Rachmühl, C.; Kühnert, D.; Achermann, Y.; Zbinden, R.; Benussi, S.; et al. In-host evolution of Staphylococcus epidermidis in a pacemaker-associated endocarditis resulting in increased antibiotic tolerance. Nat. Commun. 2019, 10, 1149. [Google Scholar] [CrossRef]
- Götz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef]
- Mack, D.; Becker, P.; Chatterjee, I.; Dobinsky, S.; Knobloch, J.K.; Peters, G.; Rohde, H.; Herrmann, M. Mechanisms of biofilm formation in Staphylococcus epidermidis and Staphylococcus aureus: Functional molecules, regulatory circuits, and adaptive responses. Int. J. Med. Microbiol. 2004, 294, 203–212. [Google Scholar] [CrossRef]
- Costerton, W. The Biofilm Primer; University of Exeter, Montana State University: Bozeman, MT, USA, 2007; Volume 1. [Google Scholar]
- Lindsay, D.; Von Holy, A. Bacterial biofilms within the clinical setting: What healthcare professionals should know. J. Hosp. Infect. 2006, 64, 313–325. [Google Scholar] [CrossRef]
- Musk, D.J., Jr.; Hergenrother, P.J. Chemical countermeasures for the control of bacterial biofilms: Effective compounds and promising targets. Curr. Med. Chem. 2006, 13, 2163–2177. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms on central venous catheters: Is eradication possible? Curr. Top. Microbiol. Immunol. 2008, 322, 133–161. [Google Scholar]
- Heilmann, C.; Hussain, M.; Peters, G.; Götz, F. Evidence for autolysin-mediated primary attachment of Staphylococcus epidermidis to a polystyrene surface. Mol. Microbiol. 1997, 24, 1013–1024. [Google Scholar] [CrossRef]
- Williams, R.J.; Henderson, B.; Sharp, L.J.; Nair, S.P. Identification of a fibronectin-binding protein from Staphylococcus epidermidis. Infect. Immun. 2002, 70, 6805–6810. [Google Scholar] [CrossRef]
- Heilmann, C.; Schweitzer, O.; Gerke, C.; Vanittanakom, N.; Mack, D.; Götz, F. Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol. Microbiol. 1996, 20, 1083–1091. [Google Scholar] [CrossRef]
- Haque, N.; Bari, M.S.; Haque, N.; Khan, R.A.; Haque, S.; Kabir, M.R.; Yasmin, T. Methicillin resistant Staphylococcus epidermidis. Mymensingh Med. J. 2011, 20, 326–331. [Google Scholar]
- May, L.; Klein, E.Y.; Rothman, R.E.; Laxminarayan, R. Trends in antibiotic resistance in coagulase-negative staphylococci in the United States, 1999 to 2012. Antimicrob. Agents Chemother. 2014, 58, 1404–1409. [Google Scholar] [CrossRef]
- Gill, S.R.; Fouts, D.E.; Archer, G.L.; Mongodin, E.F.; Deboy, R.T.; Ravel, J.; Paulsen, I.T.; Kolonay, J.F.; Brinkac, L.; Beanan, M.; et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J. Bacteriol. 2005, 187, 2426–2438. [Google Scholar] [CrossRef] [PubMed]
- CA-SFM–Antibiogram Committee of French Society of Microbiology–May 2021 v1.0. Available online: https://www.sfm-microbiologie.org (accessed on 22 October 2022).
- Ankrum, A.; Hall, B.G. Population dynamics of Staphylococcus aureus in cystic fibrosis patients to determine transmission events by use of whole-genome sequencing. J. Clin. Microbiol. 2017, 55, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed]
- Baddour, L.M.; Christensen, G.D. Prosthetic valve endocarditis due to small-colony staphylococcal variants. Rev. Infect. Dis. 1987, 9, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The worm has turned--microbial virulence modeled in Caenorhabditis elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar] [CrossRef]
- Pacha-Olivenza, M.Á.; Rodríguez-Cano, A.; González-Martín, M.L.; Gallardo-Moreno, A.M. Kinetic of adhesion of S. epidermidis with different EPS production on Ti6Al4V surfaces. Biomed. Res. Int. 2019, 2019, 1437806. [Google Scholar] [CrossRef]
- Montanaro, L.; Speziale, P.; Campoccia, D.; Ravaioli, S.; Cangini, I.; Pietrocola, G.; Giannini, S.; Arciola, C.R. Scenery of Staphylococcus implant infections in orthopedics. Future Microbiol. 2011, 6, 1329–1349. [Google Scholar] [CrossRef]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, M.; Liu, Q.; Qin, J.; Dai, Y.; He, L.; Li, T.; Zheng, B.; Zhou, F.; Fang, J.; et al. Role of the ESAT-6 secretion system in virulence of the emerging community-associated Staphylococcus aureus lineage ST398. Sci. Rep. 2016, 6, 25163. [Google Scholar] [CrossRef]
- Even, S.; Leroy, S.; Charlier, C.; Zakour, N.B.; Chacornac, J.P.; Lebert, I.; Jamet, E.; Desmonts, M.H.; Coton, E.; Pochet, S.; et al. Low occurrence of safety hazards in coagulase negative staphylococci isolated from fermented food stuffs. Int. J. Food Microbiol. 2010, 139, 87–95. [Google Scholar] [CrossRef]
- Okee, M.S.; Joloba, M.L.; Okello, M.; Najjuka, F.C.; Katabazi, F.A.; Bwanga, F.; Nanteza, A.; Kateete, D.P. Prevalence of virulence determinants in Staphylococcus epidermidis from ICU patients in Kampala, Uganda. J. Infect. Dev. Ctries 2012, 6, 242–250. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Coelho, S.M.O.; Reinoso, E.; Pereira, I.A.; Soares, L.C.; Demo, M.; Bogni, C.; Souza, M.M.S. Virulence factors and antimicrobial resistance of Staphylococcus aureus isolated from bovine mastitis in Rio de Janeiro. Pesq. Vet. Bras. 2009, 29, 369–374. [Google Scholar] [CrossRef]
- Pinheiro, L.; Brito, C.I.; de Oliveira, A.; Martins, P.Y.; Pereira, V.C.; da Cunha, M.L. Staphylococcus epidermidis and Staphylococcus haemolyticus: Molecular detection of cytotoxin and enterotoxin genes. Toxins 2015, 7, 3688–3699. [Google Scholar] [CrossRef]
- De Lastours, V.; Chau, F.; Roy, C.; Larroque, B.; Fantin, B. Emergence of quinolone resistance in the microbiota of hospitalized patients treated or not with a fluoroquinolone. J. Antimicrob. Chemother. 2014, 69, 3393–3400. [Google Scholar] [CrossRef]
- Kolár, M.; Urbánek, K.; Látal, T. Antibiotic selective pressure and development of bacterial resistance. Int. J. Antimicrob. Agents 2001, 17, 357–363. [Google Scholar] [CrossRef]
- Cui, L.; Ma, X.; Sato, K.; Okuma, K.; Tenover, F.C.; Mamizuka, E.M.; Gemmell, C.G.; Kim, M.N.; Ploy, M.C.; El-Solh, N.; et al. Cell wall thickening is a common feature of vancomycin resistance in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 5–14. [Google Scholar] [CrossRef]
- Butin, M.; Martins-Simões, P.; Picaud, J.C.; Kearns, A.; Claris, O.; Vandenesch, F.; Laurent, F.; Rasigade, J.P. Adaptation to vancomycin pressure of multiresistant Staphylococcus capitis NCRS-A involved in neonatal sepsis. J. Antimicrob. Chemother. 2015, 70, 3027–3031. [Google Scholar] [CrossRef]
- Blake, K.L.; O’Neill, A.J.; Mengin-Lecreulx, D.; Henderson, P.J.; Bostock, J.M.; Dunsmore, C.J.; Simmons, K.J.; Fishwick, C.W.; Leeds, J.A.; Chopra, I. The nature of Staphylococcus aureus MurA and MurZ and approaches for detection of peptidoglycan biosynthesis inhibitors. Mol. Microbiol. 2009, 72, 335–343. [Google Scholar] [CrossRef]
- Bæk, K.T.; Thøgersen, L.; Mogenssen, R.G.; Mellergaard, M.; Thomsen, L.E.; Petersen, A.; Skov, S.; Cameron, D.R.; Peleg, A.Y.; Frees, D. Stepwise decrease in daptomycin susceptibility in clinical Staphylococcus aureus isolates associated with an initial mutation in rpoB and a compensatory inactivation of the clpX gene. Antimicrob. Agents Chemother. 2015, 59, 6983–6991. [Google Scholar] [CrossRef]
- Kelley, W.L.; Lew, D.P.; Renzoni, A. Antimicrobial peptide exposure and reduced susceptibility to daptomycin: Insights into a complex genetic puzzle. J. Infect. Dis. 2012, 206, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.B.; Jevitt, L.A.; Hageman, J.; McDonald, L.C.; Tenover, F.C. An association between reduced susceptibility to daptomycin and reduced susceptibility to vancomycin in Staphylococcus aureus. Clin. Infect. Dis. 2006, 42, 1652–1653. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Huntelman, N.A.; Singh, N.; Valencia, A.; Mira, P.; Sakayan, M.; Boucher, I.; Tang, S.; Brennan, K.; Gianvecchio, C.; Fitz-Gibbon, S.; et al. Evolution of antibiotic cross-resistance and collateral sensitivity in Staphylococcus epidermidis using the mutant prevention concentration and the mutant selection window. Evol. Appl. 2020, 13, 808–823. [Google Scholar] [CrossRef] [PubMed]
- Roch, M.; Gagetti, P.; Davis, J.; Ceriana, P.; Errecalde, L.; Corso, A.; Rosato, A.E. Daptomycin resistance in clinical MRSA strains is associated with a high biological fitness cost. Front. Microbiol. 2017, 8, 2303. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Feng, L.; Qian, C.; Chen, T.; Wang, S.; Zhang, Y.; Zheng, X.; Wang, L.; Liu, S.; Zhou, T.; et al. Acquisition of Daptomycin Resistance by Enterococcus faecium Confers Collateral Sensitivity to Glycopeptides. Front. Microbiol. 2022, 13, 815600. [Google Scholar] [CrossRef]
- Su, J.; Iehara, M.; Yasukawa, J.; Matsumoto, Y.; Hamamoto, H.; Sekimizu, K. A novel mutation in the vraS gene of Staphylococcus aureus contributes to reduce susceptibility against daptomycin. J. Antibiot. 2015, 68, 646–648. [Google Scholar] [CrossRef]
- Wu, Y.; Meng, Y.; Qian, L.; Ding, B.; Han, H.; Chen, H.; Bai, L.; Qu, D.; Wu, Y. The Vancomycin Resistance-Associated Regulatory System VraSR Modulates Biofilm Formation of Staphylococcus epidermidis in an ica-Dependent Manner. mSphere 2021, 6, e0064121. [Google Scholar] [CrossRef]
- Cafiso, V.; Bertuccio, T.; Purrello, S.; Campanile, F.; Mammina, C.; Sartor, A.; Raglio, A.; Stefani, S. dltA overexpression: A strain-independent keystone of daptomycin resistance in methicillin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2014, 43, 26–31. [Google Scholar] [CrossRef]
- Neoh, H.M.; Cui, L.; Yuzawa, H.; Takeuchi, F.; Matsuo, M.; Hiramatsu, K. Mutated response regulator graR is responsible for phenotypic conversion of Staphylococcus aureus from heterogeneous vancomycin-intermediate resistance to vancomycin-intermediate resistance. Antimicrob. Agents Chemother. 2008, 52, 45–53. [Google Scholar] [CrossRef]
- Thitiananpakorn, K.; Aiba, Y.; Tan, X.E.; Watanabe, S.; Kiga, K.; Sato’o, Y.; Boonsiri, T.; Li, F.Y.; Sasahara, T.; Taki, Y.; et al. Association of mprF mutations with cross-resistance to daptomycin and vancomycin in methicillin-resistant Staphylococcus aureus (MRSA). Sci. Rep. 2020, 10, 16107. [Google Scholar] [CrossRef]
- Cameron, D.R.; Lin, Y.H.; Trouillet-Assant, S.; Tafani, V.; Kostoulias, X.; Mouhtouris, E.; Skinner, N.; Visvanathan, K.; Baines, S.L.; Howden, B.; et al. Vancomycin-intermediate Staphylococcus aureus isolates are attenuated for virulence when compared with susceptible progenitors. Clin. Microbiol. Infect. 2017, 23, 767–773. [Google Scholar] [CrossRef]
- Morroni, G.; Brenciani, A.; Brescini, L.; Fioriti, S.; Simoni, S.; Pocognoli, A.; Mingoia, M.; Giovanetti, E.; Barchiesi, F.; Giacometti, A.; et al. High rate of ceftobiprole resistance among clinical methicillin-resistant Staphylococcus aureus isolates from a hospital in Central Italy. Antimicrob. Agents Chemother. 2018, 62, e01663-18. [Google Scholar] [CrossRef]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol Mol Biol Rev. 2020, 3, e00026-19. [Google Scholar] [CrossRef]
- Cargill, J.S.; Upton, M. Low concentrations of vancomycin stimulate biofilm formation in some clinical isolates of Staphylococcus epidermidis. J. Clin. Pathol. 2009, 62, 1112–1116. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, F.J.; Liu, Y.; Xiong, L.R.; Xie, L.L.; Xia, P.Y. Enhancement of biofilm formation by subinhibitory concentrations of macrolides in icaADBC-positive and -negative clinical isolates of Staphylococcus epidermidis. Antimicrob. Agents Chemother. 2010, 54, 2707–2711. [Google Scholar] [CrossRef]
- Pérez-Giraldo, C.; Rodríguez-Benito, A.; Morán, F.J.; Hurtado, C.; Blanco, M.T.; Gómez-García, A.C. In-vitro slime production by Staphylococcus epidermidis in presence of subinhibitory concentrations of ciprofloxacin, ofloxacin and sparfloxacin. J. Antimicrob. Chemother. 1994, 33, 845–848. [Google Scholar] [CrossRef]
- Rachid, S.; Cho, S.; Ohlsen, K.; Hacker, J.; Ziebuhr, W. Induction of Staphylococcus epidermidis biofilm formation by environmental factors: The possible involvement of the alternative transcription factor sigB. Adv. Exp. Med. Biol. 2000, 485, 159–166. [Google Scholar]
- Both, A.; Huang, J.; Qi, M.; Lausmann, C.; Weißelberg, S.; Büttner, H.; Lezius, S.; Failla, A.V.; Christner, M.; Stegger, M.; et al. Distinct clonal lineages and within-host diversification shape invasive Staphylococcus epidermidis populations. PLoS Pathog. 2021, 17, e1009304. [Google Scholar] [CrossRef]
- Meeker, D.G.; Beenken, K.E.; Mills, W.B.; Loughran, A.J.; Spencer, H.J.; Lynn, W.B.; Smeltzer, M.S. Evaluation of antibiotics active against Methicillin-Resistant Staphylococcus aureus based on activity in an established biofilm. Antimicrob. Agents Chemother. 2016, 60, 5688–5694. [Google Scholar] [CrossRef]
- Marquès, C.; Tasse, J.; Pracros, A.; Collin, V.; Franceschi, C.; Laurent, F.; Chatellier, S.; Forestier, C. Effects of antibiotics on biofilm and unattached cells of a clinical Staphylococcus aureus isolate from bone and joint infection. J. Med. Microbiol. 2015, 64, 1021–1026. [Google Scholar] [CrossRef]
- Henriksen, A.S.; Smart, J.; Hamed, K. Comparative activity of ceftobiprole against coagulase-negative staphylococci from the BSAC Bacteraemia Surveillance Programme, 2013–2015. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Boudet, A.; Sorlin, P.; Pouget, C.; Chiron, R.; Lavigne, J.P.; Dunyach-Remy, C.; Marchandin, H. Biofilm Formation in Methicillin-Resistant Staphylococcus aureus Isolated in Cystic Fibrosis Patients Is Strain-Dependent and Differentially Influenced by Antibiotics. Front. Microbiol. 2021, 12, 750489. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A.; Nguyen, V.L.; Chamberlain, N.R. Evidence for persisters in Staphylococcus epidermidis RP62a planktonic cultures and biofilms. J. Med. Microbiol. 2011, 60, 950–960. [Google Scholar] [CrossRef]
- Cui, L.; Isii, T.; Fukuda, M.; Ochiai, T.; Neoh, H.M.; Camargo, I.L.; Watanabe, Y.; Shoji, M.; Hishinuma, T.; Hiramatsu, K. An RpoB mutation confers dual heteroresistance to daptomycin and vancomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 5222–5233. [Google Scholar] [CrossRef] [PubMed]
- Kaldalu, N.; Hauryliuk, V.; Turnbull, K.J.; La Mensa, A.; Putrinš, M.; Tenson, T. In Vitro Studies of Persister Cells. Microbiol. Mol. Biol. Rev. 2020, 84, e00070-20. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Chua, K.; Davies, J.K.; Newton, H.J.; Seemann, T.; Harrison, P.F.; Holmes, N.E.; Rhee, H.W.; Hong, J.I.; Hartland, E.L.; et al. Two novel point mutations in clinical Staphylococcus aureus reduce linezolid susceptibility and switch on the stringent response to promote persistent infection. PLoS Pathog. 2010, 6, e1000944. [Google Scholar] [CrossRef]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef]
- Conlon, K.M.; Humphreys, H.; O’Gara, J.P. icaR encodes a transcriptional repressor involved in environmental regulation of ica operon expression and biofilm formation in Staphylococcus epidermidis. J. Bacteriol. 2002, 184, 4400–4408. [Google Scholar] [CrossRef]
- Cerna, N.; Brooks, J.L.; Jefferson, K.K. Regulation of the intercellular adhesin locus regulator (icaR) by SarA, σB, and IcaR in Staphylococcus aureus. J. Bacteriol. 2008, 190, 6530–6533. [Google Scholar]
- Lavigne, J.P.; Nicolas-Chanoine, M.H.; Bourg, G.; Moreau, J.; Sotto, A. Virulent synergistic effect between Enterococcus faecalis and Escherichia coli assayed by using the Caenorhabditis elegans model. PLoS ONE 2008, 3, e3370. [Google Scholar] [CrossRef]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hébraud, M.; Bernardi, T. A new device for rapid evaluation of biofilm formation potential by bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef]
- Tasse, J.; Croisier, D.; Badel-Berchoux, S.; Chavanet, P.; Bernardi, T.; Provot, C.; Laurent, F. Preliminary results of a new antibiotic susceptibility test against biofilm installation in device-associated infections: The Antibiofilmogram®. Pathog. Dis. 2016, 74, ftw057. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarlov, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing Whole-Genome Sequencing with Sanger Sequencing for Spa Typing of Methicillin-Resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef]
- Snippy: Fast Bacterial Variant Calling from NGS Reads. Available online: https://github.com/tseemann/snippy (accessed on 24 June 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vancomycin MIC (mg/L) | Daptomycin MIC (mg/L) | Linezolid MIC (mg/L) | Ceftobiprole MIC (mg/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Before (D0) | After ATB (D15) | After No ATB (D30) | Before (D0) | After ATB (D15) | After No ATB (D30) | Before (D0) | After (D15) | After No ATB (D30) | Before (D0) | After ATB (D15) | After No ATB (D30) |
| 5 | 1 (S) * | 4 (R) | 4 (R) | 0.5 (S) | 0.5 (S) | 1 (S) | 1 (S) | 1 (S) | 1 (S) | 0.25 (S) | 1 (S) | 1 (S) |
| 26 | 1 (S) | 4 (R) | 4 (R) | 1 (S) | 1 (S) | 1 (S) | 1 (S) | 4 (S) | 4 (S) | 0.5 (S) | 1 (S) | 2 (S) |
| 82 | 1 (S) | 4 (R) | 4 (R) | 0.5 (S) | 2 (R) | 2 (R) | 1 (S) | 2 (S) | 2 (S) | 1 (S) | 4 (R) | 8 (R) |
| 97 | 1 (S) | 4 (R) | 8 (R) | 0.5 (S) | 2 (R) | 4 (R) | 1 (S) | 2 (S) | 4 (S) | 0.5 (S) | 1 (S) | 1 (S) |
| 105 | 1 (S) | 4 (R) | 4 (R) | 0.5 (S) | 2 (R) | 2 (R) | 1 (S) | 4 (S) | 2 (S) | 0.5 (S) | 4 (R) | 4(R) |
| Strain 5 | Strain 26 | Strain 82 | Strain 97 | Strain 105 | ||
|---|---|---|---|---|---|---|
| Genotyping | Genome Size (bp) | 2,517,447 | 2,509,906 | 2,553,400 | 2,475,519 | 2,528,716 |
| No. of contigs | 23 | 21 | 56 | 41 | 40 | |
| No. of ORFs * | 2262 | 2264 | 2309 | 2242 | 2267 | |
| Sequence type | 35 | 640 | 22 | 35 | 87 | |
| Virulome | agr, hld, psmα, icaA,B, IS256, bhp, atlE and esaAB | agr, hld, psmα, icaAB, IS256, bhp, atlE and esaAB | agr, hld, psmα, psm mec, icaAB, IS256, bhp and atlE | agr, hld, psmα, icaA,B, IS256, bhp, atlE and esaAB | agr, hld, psmα, psm mec, IS256, bhp, atlE and esaAB | |
| Resistome | blaZ, mecA, dfrC, norA, fusB, tetK, fosB, msrA, aad, ant(4″)-Ib and mphC | mecA, msrA, fusB, dfrC and norA, fosB | blaZ, mecA, aph(2″)-Ia, fusB, dfrC, norA, qacA and fosB | blaZ, mecA, mphC, ant(4″)-Ib, aad, msrA, fusB, dfrC, tetK, morA, mgrA and fosB | blaZ, mecA, ant(9)-Ia, aph(2″)-Ia, vatB, ermA, fusB, vgaA, dfrC, norA, fosB and qacA |
| Number of SNP | Strain 5 | Strain 26 | Strain 82 | Strain 97 | Strain 105 |
|---|---|---|---|---|---|
| After vancomycin exposure | 24 | 6 | 3 | 11 | 2 |
| After ceftobiprole exposure | - | - | 1 | - | 4 |
| After daptomycin exposure | - | - | 2 | 3 | 3 |
| Localization of SNP concerning known genes | rpoB, IS256 and icaR | murA | murA (vancomycin), pbp2 (ceftobiprole) and graR (daptomycin) | murA, rpoB (vancomycin) and vraS (daptomycin) | murA (vancomycin), pbp2 (ceftobiprole) and dltA (daptomycin) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouget, C.; Chatre, C.; Lavigne, J.-P.; Pantel, A.; Reynes, J.; Dunyach-Remy, C. Effect of Antibiotic Exposure on Staphylococcus epidermidis Responsible for Catheter-Related Bacteremia. Int. J. Mol. Sci. 2023, 24, 1547. https://doi.org/10.3390/ijms24021547
Pouget C, Chatre C, Lavigne J-P, Pantel A, Reynes J, Dunyach-Remy C. Effect of Antibiotic Exposure on Staphylococcus epidermidis Responsible for Catheter-Related Bacteremia. International Journal of Molecular Sciences. 2023; 24(2):1547. https://doi.org/10.3390/ijms24021547
Chicago/Turabian StylePouget, Cassandra, Clotilde Chatre, Jean-Philippe Lavigne, Alix Pantel, Jacques Reynes, and Catherine Dunyach-Remy. 2023. "Effect of Antibiotic Exposure on Staphylococcus epidermidis Responsible for Catheter-Related Bacteremia" International Journal of Molecular Sciences 24, no. 2: 1547. https://doi.org/10.3390/ijms24021547
APA StylePouget, C., Chatre, C., Lavigne, J.-P., Pantel, A., Reynes, J., & Dunyach-Remy, C. (2023). Effect of Antibiotic Exposure on Staphylococcus epidermidis Responsible for Catheter-Related Bacteremia. International Journal of Molecular Sciences, 24(2), 1547. https://doi.org/10.3390/ijms24021547