
Genome-Wide Identification and Characterization of the GRF Gene Family in Melastoma dodecandrum


 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Identification and Physicochemical Property Analysis of GRF
2.2. Conserved Domain and Gene Structure Analysis of GRF
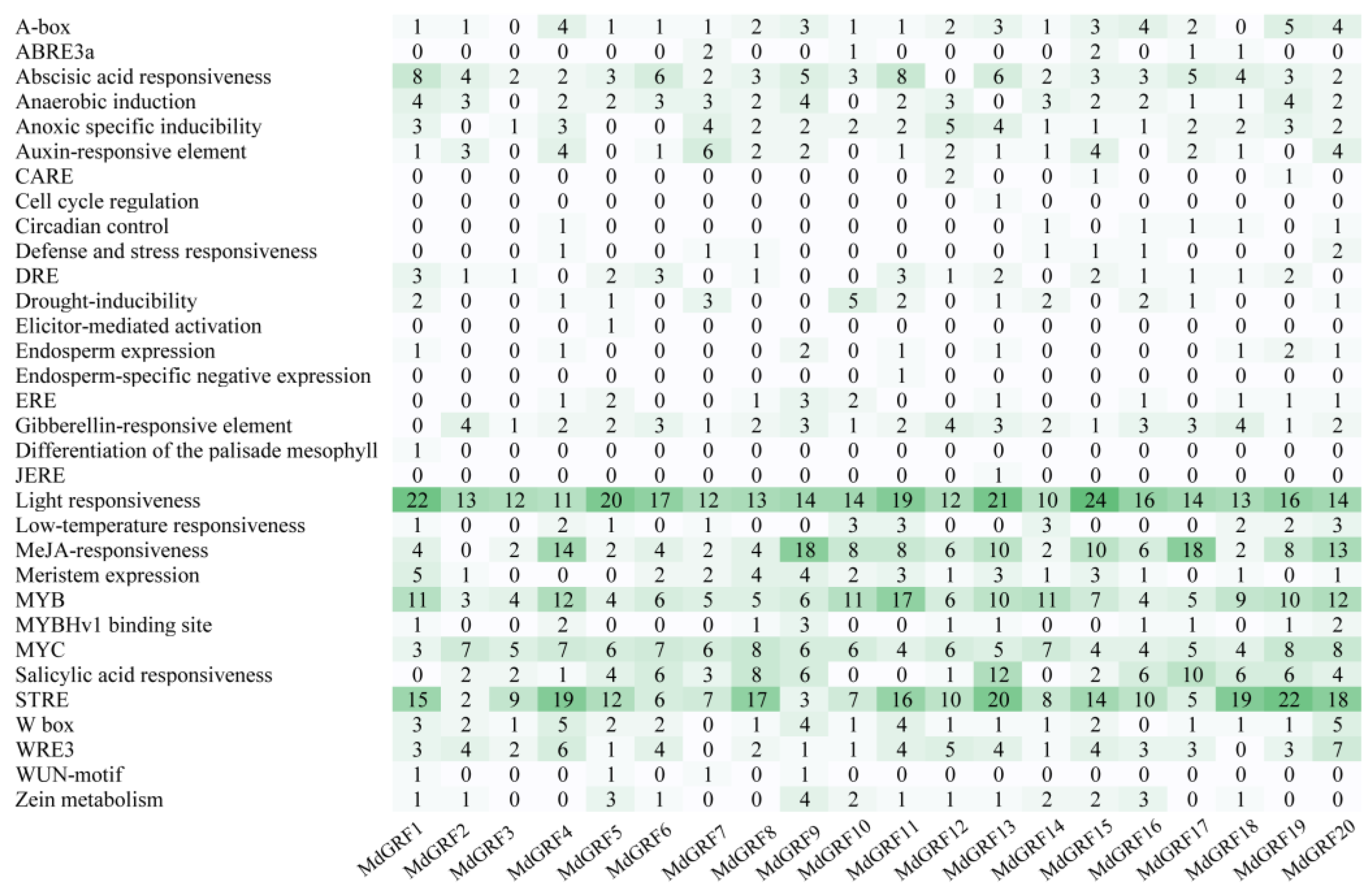
2.3. Cis-acting element prediction of GRF gene family
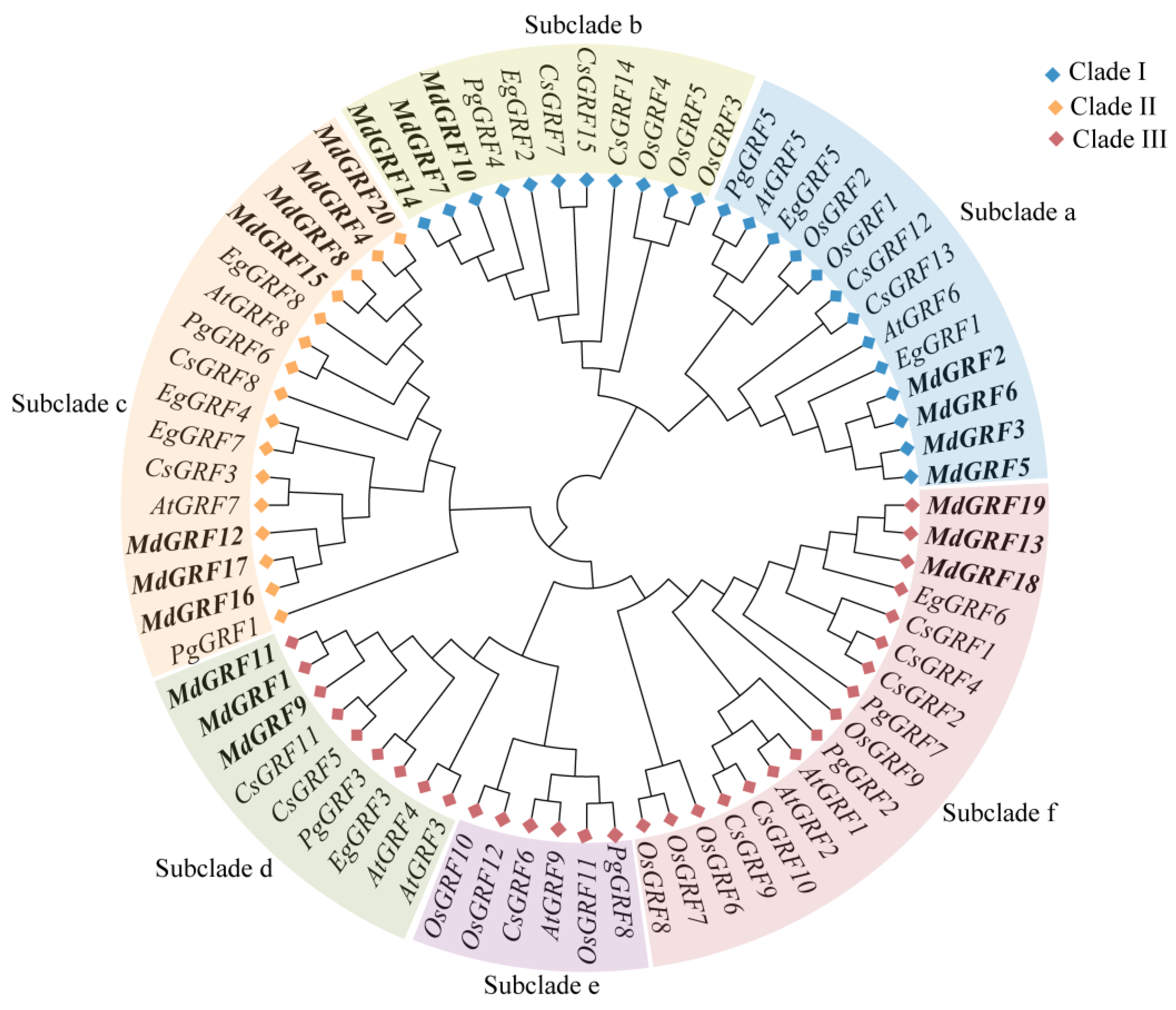
2.4. Phylogeny and Evolution Analysis of GRF Gene Family
2.5. Gene Chromosome Mapping and Collinearity Analysis
2.6. Expression Pattern Analysis of GRF Gene Family
2.7. Subcellular Localization Analysis of MdGRF19
3. Discussion
4. Materials and Methods
4.1. Data Sources
4.2. Identification and Physicochemical Property Analysis of GRF Gene Family in M. dodecandrum
4.3. Conserved Domain and Gene Structure Analysis of GRF
4.4. Phylogeny Analysis of GRF Gene Family
4.5. Chromosome Mapping, Collinearity Analysis
4.6. Cis-Acting Element Prediction of GRF Gene Family
4.7. Transcriptome Data and RT-qPCR Analysis of GRF Gene Family
4.8. Subcellular Localization Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van der Knaap, E.; Kim, J.H.; Kende, H. A novel gibberellin-in-duced gene from rice and its potential regulatory role in stem growth. Plant Physiol. 2000, 122, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-F.; Li, B.; Jia, G.-Q.; Zhang, T.-F.; Dai, J.-R.; Li, J.-S.; Wang, S.-C. Isolation and characterization of genes encoding GRF transcription factors and GIF transcriptional coactivators in maize (Zea mays L.). Plant Sci. 2008, 175, 809–817. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, Y.; Lin, Y.; Zhou, Z.; Yang, J.; Ye, N. Genome-wide identification and expression analysis of GRF gene family in Camellia sinensis. Acta Bot. Boreal. Occident. Sin. 2019, 39, 0413–0421. [Google Scholar]
- Kim, J.H.; Choi, D.; Kende, H. The AtGRF family of putative transcription factors is involved in leaf and cotyledon growth in Arabidopsis. Plant J. 2003, 36, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Qiu, N.; Ding, Q.; Li, J.; Zhang, Y.; Li, H.; Gao, J. Genome-wide identification and analysis of the growth-regulating factor family in chinese cabbage (Brassica Rapa L. Ssp. Pekinensis). BMC Genom. 2014, 15, 807. [Google Scholar] [CrossRef]
- Ma, J.-Q.; Jian, H.-J.; Yang, B.; Lu, K.; Zhang, A.-X.; Liu, P.; Li, J.-N. Genome-wide analysis and expression profiling of the GRF gene family in oilseed rape (Brassica Napus L.). Gene 2017, 620, 36–45. [Google Scholar] [CrossRef]
- Khatun, K.; Robin, A.H.K.; Park, J.-I.; Nath, U.K.; Kil Kim, C.; Lim, K.-B.; Nou, I.S.; Chung, M.-Y. Molecular characterization and expression profiling of tomato GRF transcription factor family genes in response to abiotic stresses and phytohormones. Int. J. Mol. Sci. 2017, 18, 1056. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Z.; Jin, J.; Xie, X.; Zhang, H.; Chen, Q.; Luo, Z.; Yang, J. Genome-wide identification and analysis of the growth-regulating factor family in tobacco (Nicotiana Tabacum). Gene 2017, 639, 117–127. [Google Scholar] [CrossRef]
- Shang, S.; Wu, C.; Huang, C.; Tie, W.; Yan, Y.; Ding, Z.; Xia, Z.; Wang, W.; Peng, M.; Tian, L.; et al. Genome-wide analysis of the GRF family reveals their involvement in abiotic stress response in Cassava. Genes 2018, 9, 110. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, J.; Song, C.; Zhang, L.; Gao, C.; Zhang, D.; An, N.; Mao, J.; Han, M. Genome-wide identification and expression analysis of GRF genes regulating apple tree architecture. Tree Genet. Genomes 2018, 14, 54. [Google Scholar] [CrossRef]
- Baloglu, M.C. Genome-wide in silico identification and comparison of growth regulating factor (GRF) genes in Cucurbitaceae family. Plant Omics. 2014, 7, 260–270. [Google Scholar] [CrossRef]
- Filiz, E.; Koç, İ.; TombuloğLu, H. Genome-wide identification and analysis of growth regulating factor genes in Brachypodium distachyon: In silico approaches. Turkish J. Biol. 2014, 38, 296–306. [Google Scholar] [CrossRef]
- Rukmangada, M.S.; Sumathy, R.; Sivaprasad, V.; Naik, V.G. Genome-wide identification and characterization of growth-regulating factors in mulberry (Morus spp.). Trees 2018, 32, 1695–1705. [Google Scholar] [CrossRef]
- Zhang, Z.; Quan, S.; Niu, J.; Guo, C.; Kang, C.; Liu, J.; Yuan, X. Comprehensive Identification and Analyses of the GRF Gene Family in the Whole-Genome of Four Juglandaceae Species. Int. J. Mol. Sci. 2022, 23, 12663. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Z.; Huang, J.; Zhou, X.-Q.; Hao, Y.; Chen, J.-L.; Zhou, Y.-Z.; Ahmad, S.; Lan, S.; Liu, Z.-J.; Peng, D.-H. Comprehensive Analysis for GRF Transcription Factors in sacred lotus (Nelumbo nucifera). Int. J. Mol. Sci. 2022, 23, 6673. [Google Scholar] [CrossRef]
- Horiguchi, G.; Kim, G.T.; Tsukaya, H. The transcription factor AtGRF5 and the transcription coactivator AN3 regulate cell proliferation in leaf primordia of Arabidopsis thaliana. Plant J. 2005, 43, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.E.; Ercoli, M.F.; Debernardi, J.M.; Breakfield, N.W.; Mecchia, M.A.; Sabatini, M.; Cools, T.; De Veylder, L.; Benfey, P.N.; Palatnik, J.F. MicroRNA miR396 regulates the switch between stem cells and transit-amplifying cells in Arabidopsis roots. Plant Cell 2015, 27, 3354–3366. [Google Scholar] [CrossRef]
- Kemal, A.M.; Muavviz, A.; Hüseyin, U.; Emre, S.; Örenay, B.S.; Çiğdem, Y. Order-wide in silico comparative analysis and identification of growth-regulating factor proteins in Malpighiales. Turk. J. Biol. 2016, 40, 26–42. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lee, B.H.; Jung, J.-H.; Park, S.K.; Song, J.T.; Kim, J.H. Growth-regulating factor and GRF-in-teracting factor specify meristematic cells of gynoecia and anthers. Plant Physiol. 2018, 176, 717–729. [Google Scholar] [CrossRef]
- Li, S.; Gao, F.; Xie, K.; Zeng, X.; Cao, Y.; Zeng, J.; He, Z.; Ren, Y.; Li, W.; Deng, Q.; et al. The OsmiR396c-OsGRF4-OsGIF1 regulatory module determines grain size and yield in rice. Plant Biotechnol. J. 2016, 14, 2134–2146. [Google Scholar] [CrossRef]
- Che, R.H.; Tong, H.N.; Shi, B.H.; Liu, Y.Q.; Fang, S.R.; Liu, D.P.; Xiao, Y.H.; Hu, B.; Liu, L.C.; Wang, H.R.; et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants 2005, 2, 15195. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Li, K.; Ning, L.; He, J.; Ma, X.; Li, Z.; Zhang, X.; Yin, D. Genome-wide analysis of the growth-regulating factor family in peanut (Arachis hypogaea L.). Int. J. Mol. Sci. 2019, 2, 4120. [Google Scholar] [CrossRef]
- Rodriguez, R.E.; Ercoli, M.F.; Debernardi, J.M.; Palatnik, J.F. Growth-regulating factors, a transcription factor family regulating more than just plant growth. In Plant Transcription Factors; Gonzalez, D.H., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 269–280. [Google Scholar] [CrossRef]
- Kim, J.H.; Kende, H. A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morphology in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 13374–13379. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Kim, J.H.; Kende, H. Whole genome analysis of the OsGRF gene family encoding plant-specific putative transcription activators in rice (Oryza Sativa L.). Plant Cell Physiol. 2004, 45, 897–904. [Google Scholar] [CrossRef] [PubMed]
- He, G.F.; Xia, Y.P. A fine wild ground cover plant for flower viewing Melastoma dodecandrum. North. Hor. 2005, 3, 39. [Google Scholar]
- Yu, Z.C.; Lin, X.X.; Su, J.Q.; Lin, Q.J.; Chen, Z.D. Research progress of Melastoma dodecandrum. J. Anhui Agric. Sci. 2011, 39, 16557–16559. [Google Scholar]
- Wang, J.F.; Zhang, P.H.; Wang, Y.T.; Wang, L.H.; Li, Q.; Lin, R.C.; Zhang, J.J. Research progress on Melastoma dodecandrum. Chin. Tradit. Herb. Drugs 2018, 49, 1211–1219. [Google Scholar] [CrossRef]
- Tian, N.; Liu, F.; Wu, J.W.; Liu, J.P.; Li, D.; Fu, S.; Huang, Y.J.; Cheng, C.Z. Genome-wide identification and expression analysis of GRF gene family in banana. J. Fruit Sci. 2020, 2, 1821–1835. [Google Scholar] [CrossRef]
- Cao, D.; Wang, J.; Ju, Z.; Liu, Q.; Li, S.; Tian, H.; Fu, D.; Zhu, H.; Luo, Y.; Zhu, B. Regulations on growth and development in tomato cotyledon, flower and fruit via destruction of miR396 with short tandem target mimic. Plant Sci. 2016, 247, 1–12. [Google Scholar] [CrossRef]
- Liu, X.; Guo, L.-X.; Jin, L.-F.; Liu, Y.-Z.; Liu, T.; Fan, Y.-H.; Peng, S.-A. Identification and transcript profiles of citrus growth-regulating factor genes involved in the regulation of leaf and fruit development. Mol. Biol. Rep. 2016, 43, 1059–1067. [Google Scholar] [CrossRef]
- Büyük, I.; Aras, S. Genome-wide in silico identification, characterization and transcriptional analysis of the family of growth-regulating factors in common bean (Phaseolus vulgaris L.) subjected to polyethylene glycol-induced drought stress. Arch. Biol. Sci. 2017, 69, 5–14. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon–intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.F.; Liu, S.Q. Genome-wide identification and phylogenetic analysis of the GRF gene family in cucumbers. Genet Mol. Biol. 2011, 34, 624–633. [Google Scholar] [CrossRef]
- Chen, F.; Yang, Y.; Luo, X.; Zhou, W.; Dai, Y.; Zheng, C.; Liu, W.; Yang, W.; Shu, K. Genome-wide identification of GRF transcription factors in soybean and expression analysis of GmGRF family under shade stress. BMC Plant Biol. 2019, 19, 269. [Google Scholar] [CrossRef] [PubMed]
- Zan, T.; Zhang, L.; Xie, T.T.; Li, L.Q. Genome-wide identification and analysis of the growth-regulating factor (GRF) gene family and GRF-interacting factor family in Triticum aestivum L. Biochem. Genet. 2020, 58, 705–724. [Google Scholar] [CrossRef]
- Kim, J.H.; Tsukaya, H. Regulation of plant growth and development by the growth-regulating factor and GRF-interacting factor duo. J. Exp. Bot. 2015, 66, 6093–6107. [Google Scholar] [CrossRef] [PubMed]
- Liebsch, D.; Palatnik, J.F. MicroRNA miR396, GRF transcription factors and GIF co-regulators: A conserved plant growth regulatory module with potential for breeding and biotechnology. Curr. Opin. Plant Biol. 2020, 53, 31–42. [Google Scholar] [CrossRef]
- Kim, J.H. Biological roles and an evolutionary sketch of the GRF-GIF transcriptional complex in plants. BMB Rep. 2019, 52, 227–238. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roe-ber, B. Growth-regulating factors (GRFs): A small transcription factor family with important functions in plant biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef]
- Hao, Y.; Zhou, Y.-Z.; Chen, B.; Chen, G.-Z.; Wen, Z.-Y.; Zhang, D.; Sun, W.-H.; Liu, D.-K.; Huang, J.; Chen, J.-L.; et al. The Melastoma dodecandrum genome and the evolution of Myrtales. J. Genet. Genom. 2022, 49, 120–131. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S. Hisat: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol. 2019, 20, 278. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Fatima, F.; Haider, M.S.; Shazadee, H.; Liu, Z.; Zheng, T.; Fang, J. Genome-wide identification and expression profiling of the polygalacturonase (PG) and pectin methylesterase (PME) genes in grapevine (Vitis vinifera L.). Int. J. Mol. Sci. 2019, 20, 3180. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | Number of Amino Acids | Molecular Weight (Average) | PI | Instability Index | Grand Average of Hydropathicity (GRAVY) | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| MdGRF1 | DR000066 | 441 | 48,554.34 | 7.34 | 54.19 | −0.693 | Nucleus |
| MdGRF2 | DR000875 | 405 | 45,832.01 | 9.19 | 61.19 | −0.76 | Nucleus |
| MdGRF3 | DR001546 | 386 | 43,184.82 | 8.69 | 57.13 | −0.816 | Nucleus |
| MdGRF4 | DR022817 | 399 | 44,121.32 | 9.3 | 57.08 | −0.809 | Nucleus |
| MdGRF5 | DR009489 | 292 | 31,752.63 | 7.2 | 44.85 | −0.475 | Nucleus |
| MdGRF6 | DR005587 | 592 | 63,117.05 | 8.12 | 49.33 | −0.518 | Nucleus |
| MdGRF7 | DR017530 | 387 | 42,203.03 | 6.85 | 61.5 | −0.674 | Nucleus |
| MdGRF8 | DR016167 | 277 | 29,999.53 | 7.72 | 44.32 | −0.569 | Nucleus |
| MdGRF9 | DR028300 | 477 | 51,389.04 | 6.62 | 56.85 | −0.623 | Nucleus |
| MdGRF10 | DR013209 | 567 | 60,022.76 | 6.44 | 67.12 | −0.796 | Nucleus |
| MdGRF11 | DR022630 | 432 | 46,814.3 | 8.02 | 45.27 | −0.542 | Nucleus |
| MdGRF12 | DR017665 | 298 | 33,057.79 | 8.19 | 53.3 | −0.652 | Nucleus |
| MdGRF13 | DR030091 | 439 | 48,810.12 | 8.79 | 69.76 | −0.859 | Nucleus |
| MdGRF14 | DR012552 | 400 | 44,738.51 | 8.97 | 60.93 | −0.839 | Nucleus |
| MdGRF15 | DR013042 | 303 | 33,526.3 | 8.89 | 53.52 | −0.706 | Nucleus |
| MdGRF16 | DR006760 | 443 | 49,686.1 | 6.99 | 74.3 | −0.773 | Nucleus |
| MdGRF17 | DR016005 | 359 | 39,646.59 | 8.67 | 54.35 | −0.585 | Nucleus |
| MdGRF18 | DR022043 | 345 | 36,736.86 | 8.8 | 49.51 | −0.481 | Nucleus |
| MdGRF19 | DR034227 | 637 | 68,659.67 | 8.02 | 54.68 | −0.486 | Nucleus |
| MdGRF20 | DR035468 | 551 | 59,247.32 | 5.94 | 59.39 | −0.662 | Nucleus |
| Sequence 1 | Sequence 2 | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| MdGRF1 | MdGRF11 | 0.173493307 | 0.398825333 | 0.43501075 |
| MdGRF2 | MdGRF5 | 0.132485395 | 0.850783938 | 0.155721552 |
| MdGRF2 | MdGRF6 | 0.063855643 | 0.280841154 | 0.227372814 |
| MdGRF3 | MdGRF5 | 0.077035229 | 0.21431482 | 0.359448912 |
| MdGRF3 | MdGRF6 | 0.151447669 | 0.797959229 | 0.189793743 |
| MdGRF6 | MdGRF5 | 0.141888261 | 0.907851387 | 0.156290185 |
| MdGRF8 | MdGRF4 | 0.215952273 | 0.982105616 | 0.219887015 |
| MdGRF10 | MdGRF7 | 0.139852066 | 0.560101487 | 0.249690582 |
| MdGRF11 | MdGRF9 | 0.350866443 | 0.976254354 | 0.359400643 |
| MdGRF14 | MdGRF10 | 0.143791642 | 0.5827495 | 0.246746916 |
| MdGRF14 | MdGRF7 | 0.053995183 | 0.141992753 | 0.380267179 |
| MdGRF15 | MdGRF4 | 0.227932514 | 0.860771403 | 0.264800287 |
| MdGRF15 | MdGRF8 | 0.047974242 | 0.208451325 | 0.230146014 |
| MdGRF16 | MdGRF12 | 0.258405456 | 0.941249704 | 0.274534435 |
| MdGRF16 | MdGRF17 | 0.1223847 | 0.272933451 | 0.44840491 |
| MdGRF17 | MdGRF12 | 0.325439183 | 0.969576227 | 0.335650951 |
| MdGRF18 | MdGRF13 | 0.192896922 | 0.819618927 | 0.235349521 |
| MdGRF19 | MdGRF13 | 0.061481126 | 0.201014717 | 0.305853855 |
| MdGRF19 | MdGRF18 | 0.181514036 | 0.76674553 | 0.236733087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Chen, G.-Z.; Ahmad, S.; Hao, Y.; Chen, J.-L.; Zhou, Y.-Z.; Lan, S.-R.; Liu, Z.-J.; Peng, D.-H. Genome-Wide Identification and Characterization of the GRF Gene Family in Melastoma dodecandrum. Int. J. Mol. Sci. 2023, 24, 1261. https://doi.org/10.3390/ijms24021261
Huang J, Chen G-Z, Ahmad S, Hao Y, Chen J-L, Zhou Y-Z, Lan S-R, Liu Z-J, Peng D-H. Genome-Wide Identification and Characterization of the GRF Gene Family in Melastoma dodecandrum. International Journal of Molecular Sciences. 2023; 24(2):1261. https://doi.org/10.3390/ijms24021261
Chicago/Turabian StyleHuang, Jie, Gui-Zhen Chen, Sagheer Ahmad, Yang Hao, Jin-Liao Chen, Yu-Zhen Zhou, Si-Ren Lan, Zhong-Jian Liu, and Dong-Hui Peng. 2023. "Genome-Wide Identification and Characterization of the GRF Gene Family in Melastoma dodecandrum" International Journal of Molecular Sciences 24, no. 2: 1261. https://doi.org/10.3390/ijms24021261
APA StyleHuang, J., Chen, G.-Z., Ahmad, S., Hao, Y., Chen, J.-L., Zhou, Y.-Z., Lan, S.-R., Liu, Z.-J., & Peng, D.-H. (2023). Genome-Wide Identification and Characterization of the GRF Gene Family in Melastoma dodecandrum. International Journal of Molecular Sciences, 24(2), 1261. https://doi.org/10.3390/ijms24021261