

Phytochemical Profiling of Sambucus nigra L. Flower and Leaf Extracts and Their Antimicrobial Potential against Almond Tree Pathogens

 ,
,  ,
,  ,
,  and
and 
Abstract
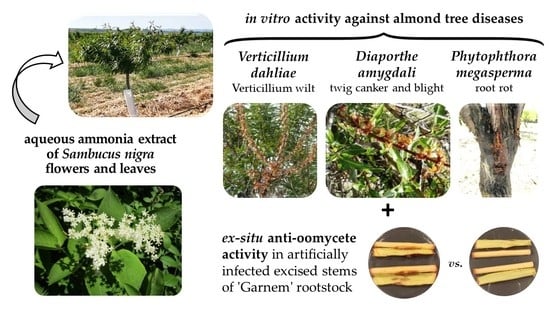
1. Introduction
2. Results
2.1. Vibrational Characterization
2.2. Analysis of the Constituents of the Flower and Leaf Extracts by GC-MS
2.3. Antifungal and Anti-Oomycete Activity Assessment
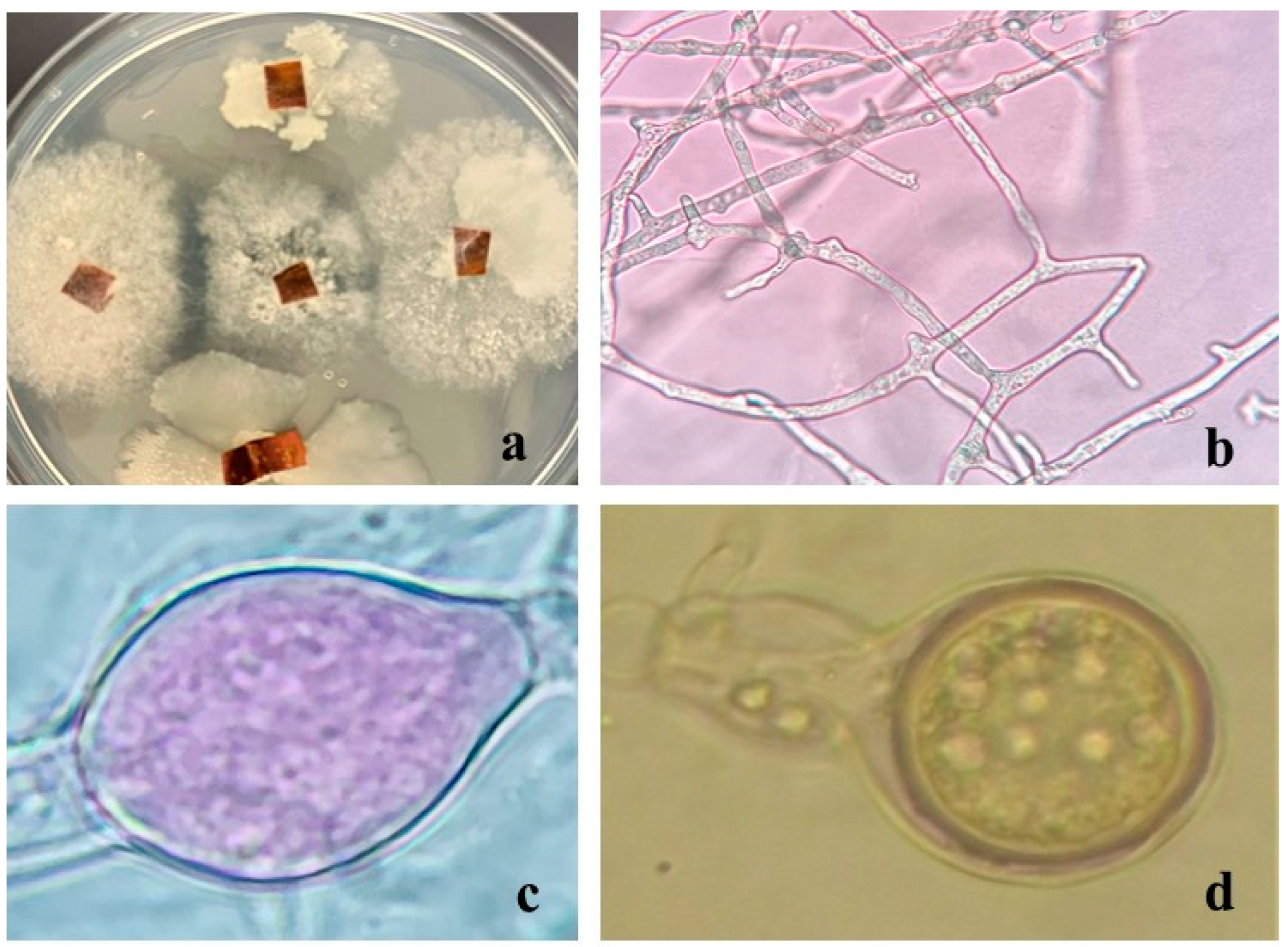
2.3.1. In Vitro Activity
2.3.2. Ex Situ Activity for the Protection of Excised Stems
3. Discussion
3.1. Phytochemical Profile
3.2. Antimicrobial Activity Comparison
3.2.1. Comparison with Other S. nigra Extracts
3.2.2. Comparison of Efficacy vs. Other Natural Compounds
3.2.3. Comparison with Conventional Fungicides
3.2.4. Mode of Action
4. Materials and Methods
4.1. Reagents
4.2. Plant Material and Extraction Procedure
4.3. Phytopathogens Isolates
4.4. Characterization Procedures
4.5. In Vitro Antifungal and Anti-Oomycete Activity
4.6. Ex Situ Protection Assays on Artificially-Inoculated Excised Stems
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boroduske, A.; Jekabsons, K.; Riekstina, U.; Muceniece, R.; Rostoks, N.; Nakurte, I. Wild Sambucus nigra L. from north-east edge of the species range: A valuable germplasm with inhibitory capacity against SARS-CoV2 S-protein RBD and hACE2 binding in vitro. Ind. Crops Prod. 2021, 165, 113438. [Google Scholar] [CrossRef] [PubMed]
- Joulain, D. The composition of the headspace from fragrant flowers: Further results. Flavour Fragr. J. 1987, 2, 149–155. [Google Scholar] [CrossRef]
- Tundis, R.; Ursino, C.; Bonesi, M.; Loizzo, M.R.; Sicari, V.; Pellicanò, T.; Manfredi, I.L.; Figoli, A.; Cassano, A. Flower and leaf extracts of Sambucus nigra L.: Application of membrane processes to obtain fractions with antioxidant and antityrosinase properties. Membranes 2019, 9, 127. [Google Scholar] [CrossRef]
- Przybylska-Balcerek, A.; Szablewski, T.; Szwajkowska-Michałek, L.; Świerk, D.; Cegielska-Radziejewska, R.; Krejpcio, Z.; Suchowilska, E.; Tomczyk, Ł.; Stuper-Szablewska, K. Sambucus nigra extracts–natural antioxidants and antimicrobial compounds. Molecules 2021, 26, 2910. [Google Scholar] [CrossRef] [PubMed]
- Gentscheva, G.; Milkova-Tomova, I.; Nikolova, K.; Buhalova, D.; Andonova, V.; Gugleva, V.; Petkova, N.; Yotkovska, I.; Ivanova, N. Antioxidant activity and chemical characteristics of Sambucus nigra L. blossom from different regions in Bulgaria. Horticulturae 2022, 8, 309. [Google Scholar] [CrossRef]
- Quattrocchi, U. CRC World Dictionary of Medicinal and Poisonous Plants: Common Names, Scientific Names, Eponyms, Synonyms, and Etymology; CRC: Boca Raton, FL, USA, 2012; p. 3960. [Google Scholar]
- Jørgensen, U.; Hansen, M.; Christensen, L.P.; Jensen, K.; Kaack, K. Olfactory and quantitative analysis of aroma compounds in elder flower (Sambucus nigra L.) drink processed from five cultivars. J. Agric. Food. Chem. 2000, 48, 2376–2383. [Google Scholar] [CrossRef]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Łysiak, G.P. Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 2018, 40, 377–390. [Google Scholar] [CrossRef]
- Palomino, O.; García-Aguilar, A.; González, A.; Guillén, C.; Benito, M.; Goya, L. Biological actions and molecular mechanisms of Sambucus nigra L. in neurodegeneration: A cell culture approach. Molecules 2021, 26, 4829. [Google Scholar] [CrossRef]
- Charlebois, D.; Byers, P.L.; Finn, C.E.; Thomas, A.L. Elderberry: Botany, horticulture, potential. In Horticultural Reviews; Janick, J., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; Volume 37, pp. 213–280. [Google Scholar] [CrossRef]
- Álvarez, C.; Barriga, A.; Albericio, F.; Romero, M.; Guzmán, F. Identification of peptides in flowers of Sambucus nigra with antimicrobial activity against aquaculture pathogens. Molecules 2018, 23, 1033. [Google Scholar] [CrossRef]
- Rodino, S.; Negoescu, C.; Butu, M.; Cornea, P.C. Preliminary investigation regarding the antifungal activity of Sambucus nigra extracts. Bull. UASVM Anim. Sci. Biotechnol. 2013, 70, 391–392. [Google Scholar]
- Selitrennikoff, C.P. Antifungal proteins. Appl. Environ. Microbiol. 2001, 67, 2883–2894. [Google Scholar] [CrossRef] [PubMed]
- Browne, G.T. Resistance to Phytophthora species among rootstocks for cultivated Prunus species. HortScience 2017, 52, 1471–1476. [Google Scholar] [CrossRef]
- Deketelaere, S.; Tyvaert, L.; França, S.C.; Höfte, M. Desirable traits of a good biocontrol agent against Verticillium wilt. Front. Microbiol. 2017, 8, 1186. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Subbarao, K.V. Verticillium systematics and evolution: How confusion impedes Verticillium wilt management and how to resolve it. Phytopathology 2014, 104, 564–574. [Google Scholar] [CrossRef]
- Asai, W.K.; Stapleton, J.J. The economic impact of Verticillium wilt on the establishment of a young almond orchard. Acta Hortic. 1994, 47, 333–337. [Google Scholar] [CrossRef]
- Gramaje, D.; Agustí-Brisach, C.; Pérez-Sierra, A.; Moralejo, E.; Olmo, D.; Mostert, L.; Damm, U.; Armengol, J. Fungal trunk pathogens associated with wood decay of almond trees on Mallorca (Spain). Pers. Mol. Phylogeny Evol. Fungi 2012, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thomidis, T.; Michailides, T.J. Studies on Diaporthe eres as a new pathogen of peach trees in Greece. Plant Dis. 2009, 93, 1293–1297. [Google Scholar] [CrossRef]
- Abreo, E.; Martínez, S.; Sessa, L.; Bettucci, L.; Lupo, S. Phomopsis cotoneastri as a pathogen associated with trunk cankers and death of young apple trees cv. Cripps Pink. J. Phytopathol. 2012, 160, 434–436. [Google Scholar] [CrossRef]
- Uddin, W.; Stevenson, K.L.; Pardo-Schultheiss, R.A.; Rehner, S.A. Pathogenic and molecular characterization of three Phomopsis isolates from peach, plum, and Asian pear. Plant Dis. 1998, 82, 732–737. [Google Scholar] [CrossRef]
- León, M.; Berbegal, M.; Rodríguez-Reina, J.M.; Elena, G.; Abad-Campos, P.; Ramón-Albalat, A.; Olmo, D.; Vicent, A.; Luque, J.; Miarnau, X.; et al. Identification and characterization of Diaporthe spp. associated with twig cankers and shoot blight of almonds in Spain. Agronomy 2020, 10, 1062. [Google Scholar] [CrossRef]
- Olaniyan, O.T.; Kunle-Alabi, O.T.; Raji, Y. Protective effects of methanol extract of Plukenetia conophora seeds and 4H-pyran-4-one 2,3-dihydro-3,5-dihydroxy-6-methyl on the reproductive function of male Wistar rats treated with cadmium chloride. JBRA Assist. Reprod. 2018, 22, 289–300. [Google Scholar] [CrossRef]
- Kader, A.; Barrett, D. Classification, composition of fruits, and postharvest maintenance of quality. In Processing Fruits. Science and Technology, 2nd ed.; Barrett, D.M., Somogyi, L., Ramaswamy, H.S., Eds.; CRC Press: Boca Raton, FL, USA, 2004; p. 20. [Google Scholar] [CrossRef]
- Tzakou, O.; Mylonas, P.; Hancianu, M.; Poiata, A. Composition and antimicrobial activity of Malabaila aurea Boiss. essential oil. J. Essent. Oil Res. 2008, 20, 270–271. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Martín-Ramos, P.; Martín-Gil, J.; Santiago-Aliste, A.; Hernández-Navarro, S.; Oliveira, R.; González-García, V. Bark extract of Uncaria tomentosa L. for the control of strawberry phytopathogens. Horticulturae 2022, 8, 672. [Google Scholar] [CrossRef]
- İşcan, G.; Demirci, F.; Kürkçüoǧlu, M.; Kıvanç, M.; Can Başer, K.H. The bioactive essential oil of Heracleum sphondylium L. subsp. ternatum (Velen.) Brummitt. Z. Nat. C 2003, 58, 195–200. [Google Scholar] [CrossRef]
- Ehsani, A.; Rezaeiyan, A.; Hashemi, M.; Aminzare, M.; Jannat, B.; Afshari, A. Antibacterial activity and sensory properties of Heracleum persicum essential oil, nisin, and Lactobacillus acidophilus against Listeria monocytogenes in cheese. Vet. World 2019, 12, 90–96. [Google Scholar] [CrossRef]
- Hamedi, A.; Pasdaran, A.; Pasdaran, A. Antimicrobial activity and analysis of the essential oils of selected endemic edible Apiaceae plants root from Caspian Hyrcanian region (North of Iran). Pharm. Sci. 2019, 25, 138–144. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, Q.; Li, L.; Liu, J.; Shen, L.; Li, H.; Qin, W. Antioxidant activity and chemical compositions of essential oil and ethanol extract of Chuanminshen violaceum. Ind. Crops Prod. 2015, 76, 290–297. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Buzón-Durán, L.; Cuchí-Oterino, J.A.; Martín-Gil, J.; Lorenzo-Vidal, B.; Martín-Ramos, P. Dwarf pomegranate (Punica granatum L. var. nana): Source of 5-HMF and bioactive compounds with applications in the protection of woody crops. Plants 2022, 11, 550. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.D.; Yong, Y.; Iqbal, M. Phytochemical investigation and free radical scavenging activities of essential oil, methanol extract and methanol fractions of Nephrolepis biserrata. Int. J. Pharm. Pharm. Sci. 2014, 6, 269–277. [Google Scholar]
- Sánchez-Hernández, E.; Buzón-Durán, L.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Physicochemical characterization and antimicrobial activity against Erwinia amylovora, Erwinia vitivora, and Diplodia seriata of a light purple Hibiscus syriacus L. cultivar. Plants 2021, 10, 1876. [Google Scholar] [CrossRef]
- Elezabeth, V.; Arumugam, S. GC-MS analysis of ethanol extract of Cyperus rotundus leaves. Int. J. Curr. Biotechnol. 2014, 2, 19–23. [Google Scholar]
- Suman, T.Y.; Elumalai, D.; Kaleena, P.K.; Rajasree, S.R.R. GC–MS analysis of bioactive components and synthesis of silver nanoparticle using Ammannia baccifera aerial extract and its larvicidal activity against malaria and filariasis vectors. Ind. Crops Prod. 2013, 47, 239–245. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, M.; Liu, F.; Zeng, S.; Hu, J. Identification of 2,3-dihydro-3,5-dihydroxy-6-methyl-4H-pyran-4-one as a strong antioxidant in glucose–histidine Maillard reaction products. Food Res. Int. 2013, 51, 397–403. [Google Scholar] [CrossRef]
- Čechovská, L.; Cejpek, K.; Konečný, M.; Velíšek, J. On the role of 2,3-dihydro-3,5-dihydroxy-6-methyl-(4H)-pyran-4-one in antioxidant capacity of prunes. Eur. Food Res. Technol. 2011, 233, 367–376. [Google Scholar] [CrossRef]
- Strittmatter, H.; Hildbrand, S.; Pollak, P. Malonic acid and derivatives. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2007; Volume 22, pp. 157–174. [Google Scholar]
- Ribeiro da Silva, M.A.V.; Cabral, J.I.T.A. Standard molar enthalpies of formation of 1-methyl-2-piperidinemethanol, 1-piperidineethanol, and 2-piperidineethanol. J. Chem. Thermodyn. 2006, 38, 1461–1466. [Google Scholar] [CrossRef]
- Jamilah, J.; Sharifa, A.; Sharifah, N. GC-MS analysis of various extracts from leaf of Plantago major used as traditional medicine. World Appl. Sci. J. 2012, 17, 67–70. [Google Scholar]
- Prabhadevi, V.; Sahaya, S.S.; Johnson, M.; Venkatramani, B.; Janakiraman, N. Phytochemical studies on Allamanda cathartica L. using GC–MS. Asian Pac. J. Trop. Biomed. 2012, 2, S550–S554. [Google Scholar] [CrossRef]
- Jatin, R.R.; Priya, R.S. Determination of bioactive components of Cynodon dactylon by GC-MS analysis & its in vitro antimicrobial activity. Int. J. Pharm. Life Sci. 2016, 7, 4880–4885. [Google Scholar]
- Hameed, R.H.; Mohammed, G.J.; Hameed, I.H. Characterization of antimicrobial metabolites produced by Salvadora persica and analysis of its chemical compounds using GC-MS and FTIR. Indian J. Public Health Res. Dev. 2018, 9, 241. [Google Scholar] [CrossRef]
- Maria, K.K.; Joanna, G.K.; Katerina, A.D.; Leland, G.K. Chemical composition and antibacterial activity against Escherichia coli of extracts of a common household plant. J. Med. Plants Res. 2021, 15, 56–63. [Google Scholar] [CrossRef]
- Villegas, A.; Espinoza, J.; Urzua, A. Piperidine alkaloids from Lobelia polyphylla Hook. & Arn. (Campanulaceae). Bol. Latinoam. Caribe Plantas Med. Aromát. 2014, 13, 205–212. [Google Scholar]
- Nibras, A.-I.; Hasan, R.M. Identification of Artemisinin compound in Artemisia herba alba belong to the Asteracea by HPLC and GC/MS. Al-Kufa Univ. J. Biol. 2019, 11, 20–33. [Google Scholar]
- Haga, Y.; Mizutani, S.; Naya, A.; Kishino, H.; Iwaasa, H.; Ito, M.; Ito, J.; Moriya, M.; Sato, N.; Takenaga, N.; et al. Discovery of novel phenylpyridone derivatives as potent and selective MCH1R antagonists. Biorg. Med. Chem. 2011, 19, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Senthilkumar, A.; Venkatesalu, V. Antibacterial and antifungal efficacy of fatty acid methyl esters from the leaves of Sesuvium portulacastrum L. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 775–780. [Google Scholar]
- Walters, D.; Raynor, L.; Mitchell, A.; Walker, R.; Walker, K. Antifungal activities of four fatty acids against plant pathogenic fungi. Mycopathologia 2004, 157, 87–90. [Google Scholar] [CrossRef]
- Api, A.M.; Belsito, D.; Biserta, S.; Botelho, D.; Bruze, M.; Burton, G.A.; Buschmann, J.; Cancellieri, M.A.; Dagli, M.L.; Date, M.; et al. RIFM fragrance ingredient safety assessment, amyl butyrate, CAS Registry Number 540-18-1. Food Chem. Toxicol. 2020, 141, 111343. [Google Scholar] [CrossRef]
- Ferreira-Santos, P.; Badim, H.; Salvador, Â.C.; Silvestre, A.J.D.; Santos, S.A.O.; Rocha, S.M.; Sousa, A.M.; Pereira, M.O.; Wilson, C.P.; Rocha, C.M.R.; et al. Chemical characterization of Sambucus nigra L. flowers aqueous extract and its biological implications. Biomolecules 2021, 11, 1222. [Google Scholar] [CrossRef] [PubMed]
- Caroline, H.; Graham, M.; David, N.; Linda, M.B.; Colin, E.G.; Paul, J.R.; Anne, L.; John, E.M.; Juluri, R.R. Antibacterial activity of elder (Sambucus nigra L.) flower or berry against hospital pathogens. J. Med. Plants Res. 2010, 4, 1805–1809. [Google Scholar]
- Cioch, M.; Satora, P.; Skotniczny, M.; Semik-Szczurak, D.; Tarko, T. Characterisation of antimicrobial properties of extracts of selected medicinal plants. Pol. J. Microbiol. 2017, 66, 463–472. [Google Scholar] [CrossRef]
- Schoss, K.; Kočevar Glavač, N.; Dolenc Koce, J.; Anžlovar, S. Supercritical CO2 plant extracts show antifungal activities against crop-borne fungi. Molecules 2022, 27, 1132. [Google Scholar] [CrossRef]
- Ramadan, K.M.A.; Ali, M.K.; Georghiou, P.E. Natural fungitoxicants of essential oil from Ageratum houstonianum L. and its application in control the root-rot diseases. J. Biol. Chem. Environ. Sci 2012, 7, 437–453. [Google Scholar]
- Abdelgaleil, S.A.M.; Saad, M.M.G.; Ariefta, N.R.; Shiono, Y. Antimicrobial and phytotoxic activities of secondary metabolites from Haplophyllum tuberculatum and Chrysanthemum coronarium. S. Afr. J. Bot. 2020, 128, 35–41. [Google Scholar] [CrossRef]
- Salamone, A.; Zizzo, G.V.; Scarito, G. The antimicrobial activity of water extracts from Labiatae. Acta Hortic. 2006, 67, 465–470. [Google Scholar] [CrossRef]
- Erdoğan, O.; Celik, A.; Zeybek, A. In vitro antifungal activity of mint, thyme, lavender extracts and essential oils on Verticillium dahliae Kleb. Fresenius Environ. Bull. 2016, 25, 4856–4862. [Google Scholar]
- Varo, A.; Mulero-Aparicio, A.; Adem, M.; Roca, L.F.; Raya-Ortega, M.C.; López-Escudero, F.J.; Trapero, A. Screening water extracts and essential oils from Mediterranean plants against Verticillium dahliae in olive. Crop Prot. 2017, 92, 168–175. [Google Scholar] [CrossRef]
- Rezayan, A.; Ehsani, A. Evaluation of the chemical compounds and antibacterial properties of the aerial parts of Persian Heracleum persicum essence. J. Babol Univ. Med. Sci. 2015, 17, 26–32. [Google Scholar]
- Halder, B.; Maity, H.S.; Banerjee, F.; Kachave, A.B.; Nag, A. Water extract of Tamarindus indica seed ash: An agro-waste green medium for one-pot three-component approach for the synthesis of 4H-pyran derivatives. Polycycl. Aromat. Compd. 2020, 42, 3302–3317. [Google Scholar] [CrossRef]
- Matloubi Moghaddam, F.; Daneshfar, M.; Moghimi, H.; Daneshfar, Z. Ferric metformin drug complex supported on magnetic nanofiber cellulose; An efficient access to 4-H pyrans derivatives and determination of their antimicrobial activity. Synth. Commun. 2022, 52, 974–993. [Google Scholar] [CrossRef]
- Dekamin, M.G.; Eslami, M.; Maleki, A. Potassium phthalimide-N-oxyl: A novel, efficient, and simple organocatalyst for the one-pot three-component synthesis of various 2-amino-4H-chromene derivatives in water. Tetrahedron 2013, 69, 1074–1085. [Google Scholar] [CrossRef]
- Alghamdi, A.I.; Ababutain, I.M. Phytochemical screening and antibacterial activity of Eucalyptus camaldulensis’s leaves and bark extracts. Asian J. Sci. Res. 2019, 12, 202–210. [Google Scholar] [CrossRef]
- Mitra, S.; Anand, U.; Jha, N.K.; Shekhawat, M.S.; Saha, S.C.; Nongdam, P.; Rengasamy, K.R.R.; Proćków, J.; Dey, A. Anticancer applications and pharmacological properties of piperidine and piperine: A comprehensive review on molecular mechanisms and therapeutic perspectives. Front. Pharmacol. 2022, 12, 772418. [Google Scholar] [CrossRef]
- Thawabteh, A.; Juma, S.; Bader, M.; Karaman, D.; Scrano, L.; Bufo, S.; Karaman, R. The biological activity of natural alkaloids against herbivores, cancerous cells and pathogens. Toxins 2019, 11, 656. [Google Scholar] [CrossRef]
- Riahi, L.; Elferchichi, M.; Ghazghazi, H.; Jebali, J.; Ziadi, S.; Aouadhi, C.; Chograni, H.; Zaouali, Y.; Zoghlami, N.; Mliki, A. Phytochemistry, antioxidant and antimicrobial activities of the essential oils of Mentha rotundifolia L. in Tunisia. Ind. Crops Prod. 2013, 49, 883–889. [Google Scholar] [CrossRef]
- Fyhrquist, P.; Virjamo, V.; Hiltunen, E.; Julkunen-Tiitto, R. Epidihydropinidine, the main piperidine alkaloid compound of Norway spruce (Picea abies) shows promising antibacterial and anti-Candida activity. Fitoterapia 2017, 117, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, T.M.A.; Ammare, Y. Synthesis and evaluation of antimicrobial activities of new piperidine derivatives. Biointerface Res. Appl. Chem. 2020, 10, 7177–7186. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2007; p. 804. [Google Scholar]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef]
- Matheron, M.E. Seasonal variation in susceptibility of Juglans hindsii and Paradox rootstocks of English walnut trees to Phytophthora citricola. Phytopathology 1985, 75, 970–972. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Balduque-Gil, J.; Barriuso-Vargas, J.J.; Casanova-Gascón, J.; González-García, V.; Cuchí-Oterino, J.A.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Holm oak (Quercus ilex subsp ballota (Desf.) Samp.) bark aqueous ammonia extract for the control of invasive forest pathogens. Int. J. Mol. Sci. 2022, 23, 11882. [Google Scholar] [CrossRef]
- Si, Y.; Haxim, Y.; Wang, L. Optimum sterilization method for in vitro cultivation of dimorphic seeds of the succulent halophyte Suaeda aralocaspica. Horticulturae 2022, 8, 289. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Er, Y. In vitro and in vivo antimicrobial activity of propolis extracts against various plant pathogens. J. Plant Dis. Prot. 2021, 128, 693–701. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Chen, Q.; Zhou, J.; Xu, J.; Zhao, T.; Huang, B.; Miao, Y.; Liu, D. The role of antifungal activity of ethyl acetate extract from Artemisia argyi on Verticillium dahliae. J. Appl. Microbiol. 2021, 132, 1343–1356. [Google Scholar] [CrossRef]
- Nazari, P.; Bazi, A.; Ayatollahi, S.A.; Dolati, H.; Mahdavi, S.M.; Rafighdoost, L.; Amirmostofian, M. Synthesis and evaluation of the antimicrobial activity of spiro-4h-pyran derivatives on some Gram positive and Gram negative bacteria. Iran. J. Pharm. Res. 2017, 16, 943–952. [Google Scholar]
- El-Sayed, N.N.E.; Zaki, M.E.A.; Al-Hussain, S.A.; Ben Bacha, A.; Berredjem, M.; Masand, V.H.; Almarhoon, Z.M.; Omar, H.S. Synthesis and evaluation of some new 4H-pyran derivatives as antioxidant, antibacterial and anti-HCT-116 cells of CRC, with molecular docking, antiproliferative, apoptotic and ADME investigations. Pharmaceuticals 2022, 15, 891. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flower | Flower Extract | Leaf | Leaf Extract | Assignment |
|---|---|---|---|---|
| 3369 | O–H stretching of free hydroxyl groups | |||
| 3288 | 3231 | 3239 | 3239 | bonded O–H stretching (cellulose, hemicellulose, lignin) |
| 2921 | 2923 | 2924 | 2930 | –CH2 asymmetric stretching of alkyls (cutine, wax, pectin) |
| 2851 | 2852 | 2850 | –CH2 sym. stretch (cutine and wax)/CH2–(C6) bend (cellulose) | |
| 1733 | 1733 | 1733 | C=O stretching of alkyl ester | |
| 1685 | 1685 | 1686 | α,β-unsaturated carbonyls | |
| 1633 | 1636 | skeletal aromatic C=C ring stretching and C=O stretching | ||
| 1605 | 1602 | 1588 | 1582 | aromatic C=C stretching |
| 1519 | 1516 | 1522 | aromatic skeletal | |
| 1455 | 1462 | CH2, CH3 bending/C=C stretching, furan ring (furfural)/O–CH3 str. | ||
| 1388 1376 | 1387 1375 | 1389 | 1389 | CH3 antisymmetric bending aliphatic C–H stretching in methyl and phenol OH |
| 1312 | 1315 | 1314 | C–H vibration of the methyl group | |
| 1254 | 1254 | 1250 | 1263 | C–O–C symmetric stretching/C–O stretching |
| 1143 | 1164 | 1163 | 1154 | C–O–C asym. stretching in celluloses; C–O stretching; C–C in-plane |
1100 | 1121 1100 | in plane =C–H bending/C=C stretching/C−O−C stretching in the pyranose ring skeletal (cellulose) | ||
| 1047 | 1045 | 1046 | O–H out plane bending/C–O stretching | |
| 1015 | 1026 | 1010 | C–H bending | |
| 919 | 918 | 918 | 914 | β-glycosidic linkages (glucose units of cellulose chains) |
| 872 | 873 | C–H bending |
| Treatment | Effective Concentration (µg·mL−1) | D. amygdali | P. megasperma | V. dahliae |
|---|---|---|---|---|
| S. nigra flower extract | EC50 | 720.5 | 193.9 | 516.1 |
| EC90 | 981.7 | 241.2 | 984.1 | |
| S. nigra leaf extract | EC50 | 860.3 | 269.3 | 539.2 |
| EC90 | 1322.4 | 354.6 | 975.2 | |
| DDMP | EC50 | 152.7 | 63.6 | 159.9 |
| EC90 | 482.4 | 211.1 | 314.1 | |
| Octyl isobutyrate | EC50 | 164.4 | 87.0 | 142.4 |
| EC90 | 647.5 | 177.2 | 471.2 | |
| Piperidinemethanol | EC50 | 233.4 | 76.4 | 185.8 |
| EC90 | 635.5 | 225.6 | 447.0 |
| Commercial Fungicide | Pathogen | Radial Growth of Mycelium (mm) | Inhibition (%) | ||||
|---|---|---|---|---|---|---|---|
| Rd/10 | Rd * | Rd × 10 | Rd/10 | Rd * | Rd × 10 | ||
| Azoxystrobin | D. amygdali | 33.2 | 14.2 | 0 | 55.7 | 81.1 | 100 |
| P. megasperma | 48.1 | 34.3 | 14.5 | 35.9 | 54.3 | 80.6 | |
| V. dahliae | 26 | 24 | 0 | 65.3 | 68 | 100 | |
| Mancozeb | D. amygdali | 13.4 | 0 | 0 | 82.2 | 100 | 100 |
| P. megasperma | 61.3 | 31.3 | 0 | 18.3 | 58.3 | 100 | |
| V. dahliae | 0 | 0 | 0 | 100 | 100 | 100 | |
| Fosetyl-Al | D. amygdali | 72 | 9.9 | 0 | 4 | 86.9 | 100 |
| P. megasperma | 75 | 16.1 | 0 | 0 | 78.6 | 100 | |
| V. dahliae | 36 | 0 | 0 | 52 | 100 | 100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Hernández, E.; Balduque-Gil, J.; González-García, V.; Barriuso-Vargas, J.J.; Casanova-Gascón, J.; Martín-Gil, J.; Martín-Ramos, P. Phytochemical Profiling of Sambucus nigra L. Flower and Leaf Extracts and Their Antimicrobial Potential against Almond Tree Pathogens. Int. J. Mol. Sci. 2023, 24, 1154. https://doi.org/10.3390/ijms24021154
Sánchez-Hernández E, Balduque-Gil J, González-García V, Barriuso-Vargas JJ, Casanova-Gascón J, Martín-Gil J, Martín-Ramos P. Phytochemical Profiling of Sambucus nigra L. Flower and Leaf Extracts and Their Antimicrobial Potential against Almond Tree Pathogens. International Journal of Molecular Sciences. 2023; 24(2):1154. https://doi.org/10.3390/ijms24021154
Chicago/Turabian StyleSánchez-Hernández, Eva, Joaquín Balduque-Gil, Vicente González-García, Juan J. Barriuso-Vargas, José Casanova-Gascón, Jesús Martín-Gil, and Pablo Martín-Ramos. 2023. "Phytochemical Profiling of Sambucus nigra L. Flower and Leaf Extracts and Their Antimicrobial Potential against Almond Tree Pathogens" International Journal of Molecular Sciences 24, no. 2: 1154. https://doi.org/10.3390/ijms24021154
APA StyleSánchez-Hernández, E., Balduque-Gil, J., González-García, V., Barriuso-Vargas, J. J., Casanova-Gascón, J., Martín-Gil, J., & Martín-Ramos, P. (2023). Phytochemical Profiling of Sambucus nigra L. Flower and Leaf Extracts and Their Antimicrobial Potential against Almond Tree Pathogens. International Journal of Molecular Sciences, 24(2), 1154. https://doi.org/10.3390/ijms24021154