
Isolation and Characterization of Thermus thermophilus Strain ET-1: An Extremely Thermophilic Bacterium with Extracellular Thermostable Proteolytic Activity Isolated from El Tatio Geothermal Field, Antofagasta, Chile


Abstract
1. Introduction
2. Results
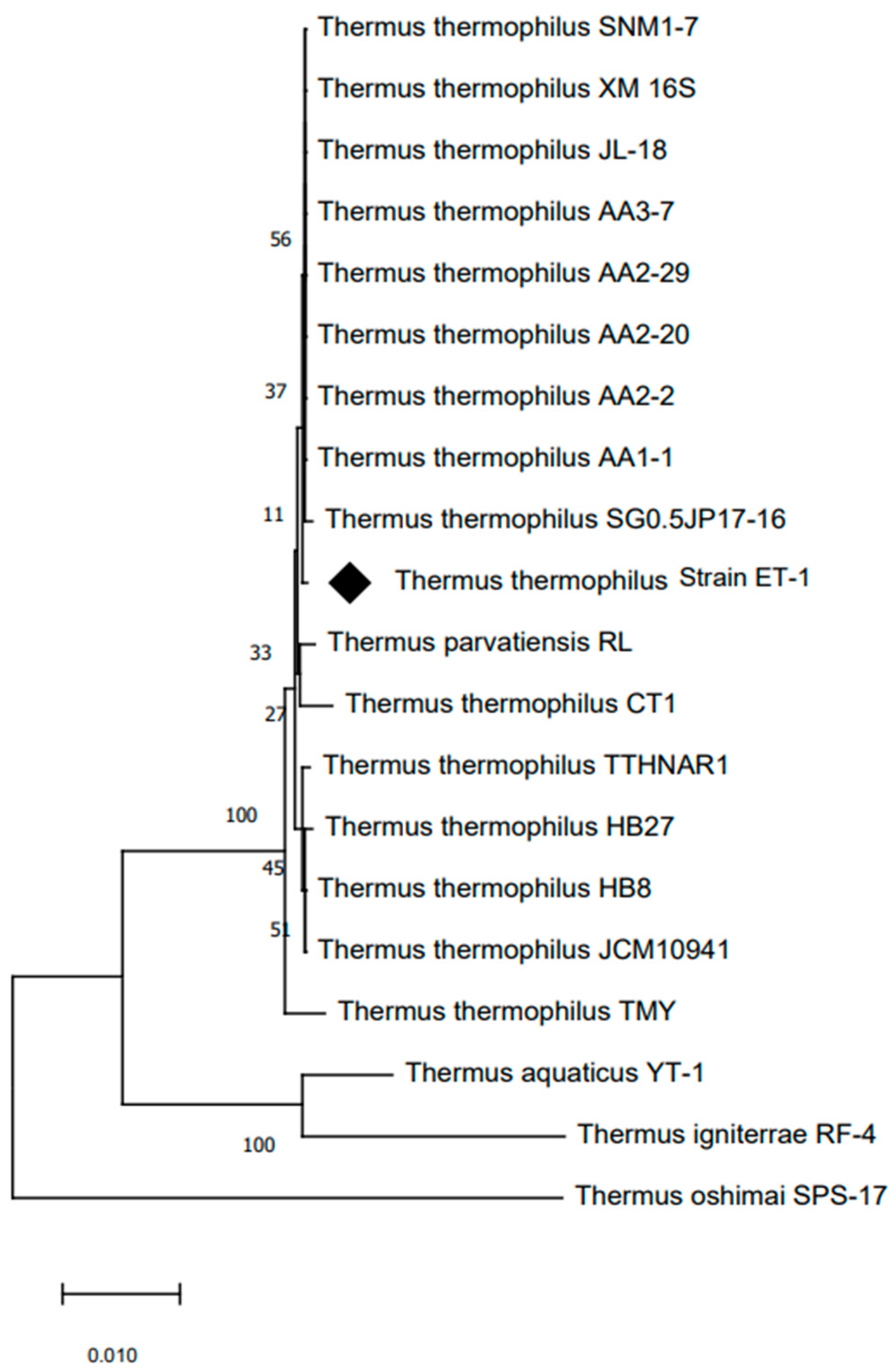
2.1. Isolation, Characterization, and Identification of CT 801 Strain
2.2. Effect of Temperature and pH on Growth
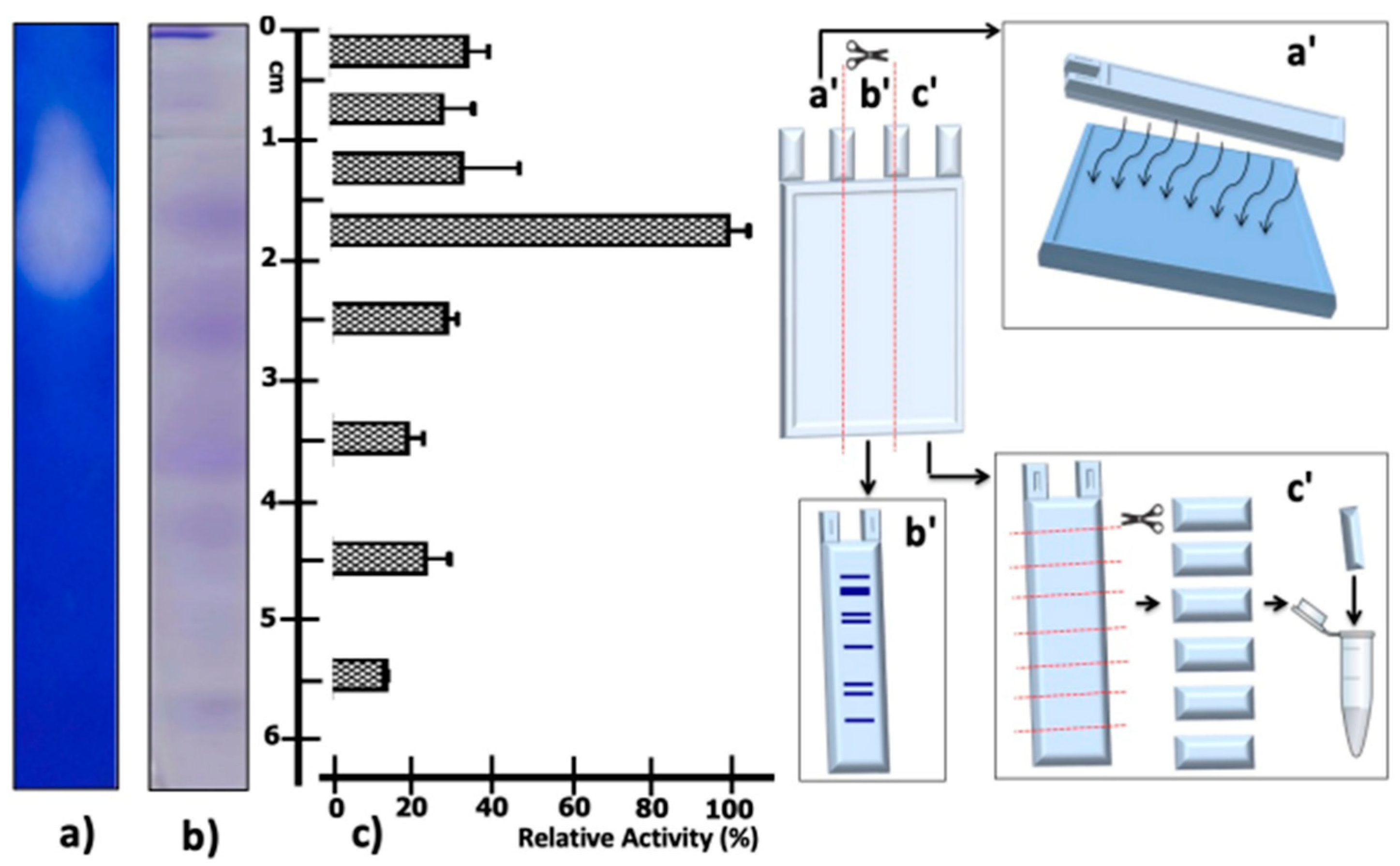
2.3. Determination of Proteolytic Activity in Native PAGE
2.4. Effect of Temperature and pH on Protease Activity and Stability
2.5. Effect of Cations on Protease Activity and Stability
2.6. Effect of Different Inhibitors on Proteolytic Activity
2.7. Effect of Denaturing Agents on Proteolytic Activity
2.8. Effect of Organic Solvent on Residual Activity
2.9. Effect of Commercially Used Detergents on Residual Activity
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Isolation and Culture
4.3. Effect of Temperature and pH on Bacterial Growth
4.4. Phylogenetic Identification
4.5. Partial Concentration and Dialysis of Proteolytic Activity
4.6. Proteolytic Activity Assays
4.7. Native PAGE
4.8. Effect of Temperature and pH on Protease Activity and Stability
4.9. Stability of Protease Activity in the Presence of Cation, Inhibitor, and Other Chemical Reagents
4.10. Effect of Denaturing Agents on the Proteolytic Activity
4.11. Effect of Organic Solvent on Protease Stability
4.12. Stability and Activity in the Presence of Commercial Detergents
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Dwivedi, V.; Kumari, K.; Gupta, S.K.; Kumari, R.; Tripathi, C.; Lata, P.; Niharika, N.; Kumar Singh, A.; Kumar, R.; Nigam, A.; et al. Thermus parvatiensis RLT sp. nov., Isolated from a Hot Water Spring, Located Atop the Himalayan Ranges at Manikaran, India. Indian J. Microbiol. 2015, 55, 357–365. [Google Scholar] [CrossRef]
- Kristjansson, J.K.; Hreggvidsson, G.O.; Alfredsson, G.A. Isolation of Halotolerant Thermus spp. from Submarine Hot Springs in Iceland. Appl. Environ. Microbiol. 1986, 52, 1313–1316. [Google Scholar] [CrossRef]
- Leone, S.; Molinaro, A.; Lindner, B.; Romano, I.; Nicolaus, B.; Parrilli, M.; Lanzetta, R.; Holst, O. The structures of glycolipids isolated from the highly thermophilic bacterium Thermus thermophilus Samu-SA1. Glycobiology 2006, 16, 766–775. [Google Scholar] [CrossRef]
- Sharp, R.J.; Williams, R.A. Properties of Thermus ruber Strains Isolated from Icelandic Hot Springs and DNA:DNA Homology of Thermus ruber and Thermus aquaticus. Appl. Environ. Microbiol. 1988, 54, 2049–2053. [Google Scholar] [CrossRef]
- Marteinsson, V.T. Isolation and characterization of Thermus thermophilus Gy1211 from a deep-sea hydrothermal vent. Extremophiles 1999, 3, 247–251. [Google Scholar] [CrossRef]
- Brock, T.D.; Freeze, H. Thermus aquaticus gen. n. and sp. n., a nonsporulating Extreme Thermophile. J. Bacteriol. 1969, 98, 289–297. [Google Scholar] [CrossRef]
- Navas, L.E.; Berretta, M.F.; Ortiz, E.M.; Benintende, G.B.; Amadio, A.F.; Zandomeni, R.O. Draft genome sequence of Thermus sp. isolate 2.9, obtained from a hot water spring located in Salta, Argentina. Genome Announc. 2015, 3, e01414-14. [Google Scholar] [CrossRef]
- Pantazaki, A.; Pritsa, A.; Kyriakidis, D. Biotechnologically relevant enzymes from Thermus thermophilus. Appl. Microbiol. Biotechnol. 2002, 58, 1–12. [Google Scholar] [CrossRef]
- Mullis, K.; Faloona, F.; Saiki, R.; Sharf, F.; Horn, G.; Erlich, H. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Biotechnol. Ser. 1992, 17, 263–273. [Google Scholar] [CrossRef]
- Cava, F.; Hidalgo, A.; Berenguer, J. Thermus thermophilus as biological model. Extremophiles 2009, 13, 213–231. [Google Scholar] [CrossRef]
- Beaujean, A.; Ducrocq-Assaf, C.; Sangwan, R.; Lilius, G.; Bülow, L.; Sangwan-Norreel, B. Engineering Direct Fructose Production in Processed Potato Tubers by Expressing a Bifunctional Alpha-Amylase/Glucose Isomerase Gene Complex. Biotechnol. Bioeng. 2000, 70, 9–16. [Google Scholar] [CrossRef]
- Chang, C.; Song, H.K.; Park, B.C.; Lee, D.S.; Suh, S.W. Thermostable xylose isomerase from Thermus caldophilus: Biochemical characterization, crystallization and preliminary X-ray analysis. Acta Crystallogr. Sect. D Biol. Crystallogr. 1999, 55, 294–296. [Google Scholar] [CrossRef]
- Fridjonsson, O.; Watzlawick, H.; Mattes, R. The structure of the α-galactosidase gene loci in Thermus brockianus ITI360 and Thermus thermophilus TH125. Extremophiles 2000, 4, 23–33. [Google Scholar] [CrossRef]
- Luo, J.; Bergstrom, D.E.; Barany, F. Improving the fidelity of Thermus thermophilus DNA ligase. Nucleic Acids Res. 1996, 24, 3071–3078. [Google Scholar] [CrossRef][Green Version]
- Myers, T.W.; Gelfand, D.H. Reverse transcription and DNA amplification by a Thermus thermophilus DNA polymerase. Biochemistry 1991, 30, 7661–7666. [Google Scholar] [CrossRef]
- Matsuzawa, H.; Hamaoki, M.; Ohta, T. Production of thermophilic extracellular proteases (aqualysins I and II) by Thermus aquaticus YT-1, an extreme thermophile. Agric. Biol. Chem. 1983, 47, 25–28. [Google Scholar] [CrossRef]
- Cowan, D.A.; Daniel, R.M. Purification and some properties of an extracellular protease (caldolysin) from an extreme thermophile. Biochim. Biophys. Acta BBA Protein Struct. Mol. Enzymol. 1982, 705, 293–305. [Google Scholar] [CrossRef]
- Peek, K.; Daniel, R.M.; Monk, C.; Parker, L.; Coolbear, T. Purification and characterization of a thermostable proteinase isolated from Thermus sp. strain Rt41A. Eur. J. Biochem. 1992, 207, 1035–1044. [Google Scholar] [CrossRef]
- Jones, C.W.; Morgan, H.W.; Daniel, R.M. Aspects of protease production by Thermus strain Ok6 and other New Zealand isolates. Microbiology 1988, 134, 191–198. [Google Scholar] [CrossRef][Green Version]
- Xie, G.; Shao, Z.; Zong, L.; Li, X.; Cong, D.; Huo, R. Heterologous expression and characterization of a novel subtilisin-like protease from a thermophilic Thermus thermophilus HB8. Int. J. Biol. Macromol. 2019, 138, 528–535. [Google Scholar] [CrossRef]
- Antranikian, G.; Vorgias, C.E.; Bertoldo, C. Extreme environments as a resource for microorganisms and novel biocatalysts. Adv. Biochem. Eng. Biotechnol. 2005, 96, 219–262. [Google Scholar]
- Lahsen, A.; Trujillo, P. The geothermal field of El Tatio, Chile. In Proceedings of the Second United Nations Symposium on the Development and Use of Geothermal Resources, San Francisco, CA, USA, 20–29 May 1975; Volume 1, pp. 170–177. [Google Scholar]
- Phoenix, V.R.; Bennett, P.C.; Engel, A.S.; Tyler, S.W.; Ferris, F.G. Chilean high-altitude hot-spring sinters: A model system for UV screening mechanisms by early Precambrian cyanobacteria. Geobiology 2006, 4, 15–28. [Google Scholar] [CrossRef]
- Munoz-Saez, C.; Namiki, A.; Manga, M. Geyser eruption intervals and interactions: Examples from El Tatio, Atacama, Chile. J. Geophys. Res. 2015, 120, 7490–7507. [Google Scholar] [CrossRef]
- Glennon, J.A.; Pfaff, R.M. The extraordinary thermal activity of El Tatio geyser field, Antofagasta Region, Chile. GOSA Trans. 2003, 8, 31–78. [Google Scholar]
- Barbieri, R.; Cavalazzi, B. How do modern extreme hydrothermal environments inform the identification of Martian habitability? The case of the El Tatio Geyser Field. Challenges 2014, 5, 430–443. [Google Scholar] [CrossRef]
- Nicolau, C.; Reich, M.; Lynne, B. Physico-chemical and environmental controls on siliceous sinter formation at the high-altitude El Tatio geothermal field, Chile. J. Volcanol. Geotherm. Res. 2014, 282, 60–76. [Google Scholar] [CrossRef]
- Fernandez-Turiel, J.L.; Garcia-Valles, M.; Gimeno-Torrente, D.; Saavedra-Alonso, J.; Martinez-Manent, S. The hot spring and geyser sinters of El Tatio, Northern Chile. Sediment. Geol. 2005, 180, 125–147. [Google Scholar] [CrossRef]
- Dunckel, A.E.; Cardenas, M.B.; Sawyer, A.H.; Bennett, P.C. High-resolution in-situ thermal imaging of microbial mats at El Tatio Geyser, Chile shows coupling between community color and temperature. Geophys. Res. Lett. 2009, 36, L23403. [Google Scholar] [CrossRef]
- Engel, A.S.; Johnson, L.R.; Porter, M.L. Arsenite oxidase gene diversity among Chloroflexi and Proteobacteria from El Tatio Geyser Field, Chile. FEMS Microbiol. Ecol. 2013, 83, 745–756. [Google Scholar] [CrossRef]
- Zabel, H.P.; König, H.; Winter, J. Isolation and characterization of a new coccoid methanogen, Methanogenium tatii spec. nov. from a solfataric field on Mount Tatio. Arch. Microbiol. 1984, 137, 308–315. [Google Scholar] [CrossRef]
- Zellner, G.; Boone, D.R.; Keswani, J.; Whitman, W.B.; Woese, C.R.; Hagelstein, A.; Tindall, B.; Stackebrandt, E. Reclassification of Methanogenium tationis and Methanogenium liminatans as Methanofollis tationis gen. nov., comb. nov. and Methanofollis liminatans comb. nov. and description of a new strain of Methanofollis liminatans. Int. J. Syst. Evol. Microbiol. 1999, 49, 247–255. [Google Scholar] [CrossRef]
- Bragger, J.M.; Daniel, R.M.; Coolbear, T.; Morgan, H.W. Very stable enzymes from extremely thermophilic Archaebacteria and Eubacteria. Appl. Microbiol. Biotechnol. 1989, 31, 556–561. [Google Scholar] [CrossRef]
- Zamost, B.L.; Nielsen, H.K.; Starnes, R.L. Thermostable enzymes for industrial applications. J. Ind. Microbiol. 1991, 8, 71–81. [Google Scholar] [CrossRef]
- Anbu, P.; Gopinath, S.C.; Chaulagain, B.P.; Lakshmipriya, T. Microbial enzymes and their applications in industries and medicine 2016. BioMed Res. Int. 2017, 2017, 2195808. [Google Scholar] [CrossRef]
- Kaper, T.; Van der Maarel, M.J.E.C.; Euverink, G.J.W.; Dijkhuizen, L. Exploring and exploiting starch-modifying amylomaltases from thermophiles. Biochem. Soc. Trans. 2004, 32, 279–282. [Google Scholar] [CrossRef]
- Svetlitchnyi, V.A.; Svetlichnaya, T.P.; Falkenhan, D.A.; Swinnen, S.; Knopp, D.; Läufer, A. Direct conversion of cellulose to L-lactic acid by a novel thermophilic Caldicellulosiruptor strain. Biotechnol. Biofuels Bioprod. 2022, 15, 44. [Google Scholar] [CrossRef]
- Khan, M.; Sathya, T.A. Extremozymes from metagenome: Potential applications in food processing. Crit. Rev. Food Sci. Nutr. 2018, 58, 2017–2025. [Google Scholar] [CrossRef]
- Daniel, R.M.; Cowan, D.A. Biomolecular stability and life at high temperatures. Cell. Mol. Life Sci. 2000, 57, 250–264. [Google Scholar] [CrossRef]
- Tobler, D.J.; Benning, L.G. Bacterial diversity in five Icelandic geothermal waters: Temperature and sinter growth rate effects. Extremophiles 2011, 15, 473–485. [Google Scholar] [CrossRef]
- Priyadharshini, R.; Brindha, T.; Uthandi, S. Thermophilic microbes producing industrially important enzymes from the Manikaran geothermal springs of Himachal Pradesh (India) and their application in biomass saccharification. Biomass Convers. Biorefinery 2023, 1–12. [Google Scholar] [CrossRef]
- Mohammad, B.T.; Al Daghistani, H.I.; Jaouani, A.; Abdel-Latif, S.; Kennes, C. Isolation and characterization of thermophilic bacteria from Jordanian hot springs: Bacillus licheniformis and Thermomonas hydrothermalis isolates as potential producers of thermostable enzymes. Int. J. Microbiol. 2017, 2017, 6943952. [Google Scholar] [CrossRef]
- Oztas Gulmus, E.; Gormez, A. Identification and characterization of novel thermophilic bacteria from hot springs, Erzurum, Turkey. Curr. Microbiol. 2020, 77, 979–987. [Google Scholar] [CrossRef]
- Hollocher, T.C.; Kristjánsson, J.K. Thermophilic denitrifying bacteria: A survey of hot springs in Southwestern Iceland. FEMS Microbiol. Ecol. 1992, 10, 113–119. [Google Scholar] [CrossRef]
- Johnson, D.B.; Okibe, N.; Roberto, F.F. Novel thermo-acidophilic bacteria isolated from geothermal sites in Yellowstone National Park: Physiological and phylogenetic characteristics. Arch. Microbiol. 2003, 180, 60–68. [Google Scholar] [CrossRef]
- Bendia, A.G.; Araujo, G.G.; Pulschen, A.A.; Contro, B.; Duarte, R.T.; Rodrigues, F.; Galante, D.; Pellizari, V.H. Surviving in hot and cold: Psychrophiles and thermophiles from Deception Island volcano, Antarctica. Extremophiles 2018, 22, 917–929. [Google Scholar] [CrossRef]
- Oshima, T.; Imahori, K. Description of Thermus thermophilus (Yoshida and Oshima) comb. nov., a nonsporulating thermophilic bacterium from a Japanese thermal spa. Int. J. Syst. Evol. Microbiol. 1974, 24, 102–112. [Google Scholar] [CrossRef]
- Murugapiran, S.K.; Huntemann, M.; Wei, C.L.; Han, J.; Detter, J.C.; Han, C.; Han, C.; Erkkila, T.C.; Teshima, H.; Chen, H.; et al. Thermus oshimai JL-2 and T. thermophilus JL-18 genome analysis illuminates pathways for carbon, nitrogen, and sulfur cycling. Stand. Genom. Sci. 2013, 7, 449–468. [Google Scholar] [CrossRef]
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef]
- Sato, S.; Hutchinson, C.A., 3rd; Harris, J.I. A thermostable sequence-specific endonuclease from Thermus aquaticus. Proc. Natl. Acad. Sci. USA 1977, 74, 542–546. [Google Scholar] [CrossRef]
- Chien, A.; Edgar, D.B.; Trela, J.M. Deoxyribonucleic acid polymerase from the extreme thermophile Thermus aquaticus. J. Bacteriol. 1976, 127, 1550–1557. [Google Scholar] [CrossRef]
- Vieille, C.; Zeikus, J.G. Thermozymes: Identifying molecular determinants of protein structural and functional stability. Trends Biotechnol. 1996, 14, 183–190. [Google Scholar] [CrossRef]
- Matsuzawa, H.; Tokugawa, K.; Hamaoki, M.; Mizoguchi, M.; Taguchi, H.; Terada, I.; Kwon, S.; Ohta, T. Purification and characterization of aqualysin I (a thermophilic alkaline serine protease) produced by Thermus aquaticus YT-1. Eur. J. Biochem. 1988, 171, 441–447. [Google Scholar] [CrossRef]
- Carballeira, N.; Nazabal, M.; Brito, J.; Garcia, O. Purification of a thermostable DNA polymerase from Thermus thermophilus HB8, useful in the polymerase chain reaction. Biotechniques 1990, 9, 276–281. [Google Scholar]
- Domínguez, A.; Deive, F.J.; Pastrana, L.; Rúa, M.L.; Longo, M.A.; Sanroman, M.A. Thermostable lipolytic enzymes production in batch and continuous cultures of Thermus thermophilus HB27. Bioprocess. Biosyst. Eng. 2010, 33, 347–354. [Google Scholar] [CrossRef]
- Asahara, Y.; Atsuta, K.; Motohashi, K.; Taguchi, H.; Yohda, M.; Yoshida, M. FtsH recognizes proteins with unfolded structure and hydrolyzes the carboxyl side of hydrophobic residues. J. Biochem. 2000, 127, 931–937. [Google Scholar] [CrossRef]
- Watanabe, S.; Muramatsu, T.; Ao, H.; Hirayama, Y.; Takahashi, K.; Tanokura, M.; Kuchino, Y. Molecular cloning of the Lon protease gene from Thermus thermophilus HB8 and characterization of its gene product. Eur. J. Biochem. 1999, 266, 811–819. [Google Scholar] [CrossRef]
- Meinnel, T.; Lazennec, C.; Villoing, S.; Blanquet, S. Structure-function relationships within the peptide deformylase family. Evidence for a conserved architecture of the active site involving three conserved motifs and a metal ion. J. Mol. Biol. 1997, 267, 749–761. [Google Scholar] [CrossRef]
- Ohtani, N.; Tomita, M.; Itaya, M. An extreme thermophile, Thermus thermophilus, is a polyploid bacterium. J. Bacteriol. 2010, 192, 5499–5505. [Google Scholar] [CrossRef]
- Lasa, I.; Berenguer, J. Thermophilic enzymes and their biotechnological potential. Microbiologia 1993, 9, 77–89. [Google Scholar]
- Steinhardt, J. Properties of hemoglobin and pepsin in solutions of urea and other amides. J. Biol. Chem. 1938, 123, 543–575. [Google Scholar] [CrossRef]
- Rajagopalan, K.V.; Fridovich, I.; Handler, P. Competitive inhibition of enzyme activity by urea. J. Biol. Chem. 1961, 236, 1059–1065. [Google Scholar] [CrossRef]
- Souza, P.M.D.; Bittencourt, M.L.D.A.; Caprara, C.C.; Freitas, M.D.; Almeida, R.P.C.D.; Silveira, D.; Fonseca, Y.M.; Magalhães, P.O. A biotechnology perspective of fungal proteases. Braz. J. Microbiol. 2015, 46, 337–346. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Families of serine peptidases. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1994; pp. 19–61. [Google Scholar]
- Kim, J.Y.; Choi, Y.L.; Cho, Y.S.; Kim, C.H.; Lee, Y.C. Independently expressed N-terminal pro-domain of aqualysin I precursor complements the folding of its mature domain to active form in Escherichia coli. J. Basic Microbiol. 2002, 42, 181–189. [Google Scholar] [CrossRef]
- Takagi, H.; Suzumura, A.; Hasuura, Y.; Hoshino, T.; Nakamori, S. Efficient selection for thermostable protease in Thermus thermophilus. Biosci. Biotechnol. Biochem. 2000, 64, 899–902. [Google Scholar] [CrossRef]
- Foophow, T.; Tanaka, S.I.; Koga, Y.; Takano, K.; Kanaya, S. Subtilisin-like serine protease from hyperthermophilic archaeon Thermococcus kodakaraensis with N-and C-terminal propeptides. Protein Eng. Des. Sel. 2010, 23, 347–355. [Google Scholar] [CrossRef]
- Zhu, X.; Ohta, Y.; Jordan, F.; Inouye, M. Pro-sequence of subtilisin can guide the refolding of denatured subtilisin in an intermolecular process. Nature 1989, 339, 483–484. [Google Scholar] [CrossRef]
- Joo, H.S.; Chang, C.S. Production of protease from a new alkalophilic Bacillus sp. I-312 grown on soybean meal: Optimization and some properties. Process Biochem. 2005, 40, 1263–1270. [Google Scholar] [CrossRef]
- Baweja, M.; Tiwari, R.; Singh, P.K.; Nain, L.; Shukla, P. An alkaline protease from Bacillus pumilus MP 27: Functional analysis of its binding model toward its applications as detergent additive. Front. Microbiol. 2016, 7, 1195. [Google Scholar] [CrossRef]
- Ramkumar, A.; Sivakumar, N.; Gujarathi, A.M.; Victor, R. Production of thermotolerant, detergent stable alkaline protease using the gut waste of Sardinella longiceps as a substrate: Optimization and characterization. Sci. Rep. 2018, 8, 12442. [Google Scholar] [CrossRef]
- Dias, D.R.; Vilela, D.M.; Silvestre, M.P.C.; Schwan, R.F. Alkaline protease from Bacillus sp. isolated from coffee bean grown on cheese whey. World J. Microbiol. Biotechnol. 2008, 24, 2027–2034. [Google Scholar] [CrossRef]
- Gupta, R.; Beg, Q.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32. [Google Scholar]
- Joo, H.S.; Kumar, C.G.; Park, G.C.; Paik, S.R.; Chang, C.S. Oxidant and SDS-stable alkaline protease from Bacillus clausii I-52: Production and some properties. J. Appl. Microbiol. 2003, 95, 267–272. [Google Scholar] [CrossRef]
- Sharma, K.M.; Kumar, R.; Panwar, S.; Kumar, A. Microbial alkaline proteases: Optimization of production parameters and their properties. J. Genet. Eng. Biotechnol. 2017, 15, 115–126. [Google Scholar] [CrossRef]
- Tiwari, O.N.; Devi, T.B.; Devi, K.S.; Oinam, G.; Indrama, T.; Ojit, K.; Avijeet, O.; Ningshen, L. Isolation and optimization of alkaline protease producing Bacteria from undisturbed soil of NE-region of India falling under Indo-Burma biodiversity hotspots. J. Appl. Biol. Biotechnol. 2015, 3, 25–31. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Bajaj, B.K.; Sharma, P. An alkali-thermotolerant extracellular protease from a newly isolated Streptomyces sp. DP2. New Biotechnol. 2011, 28, 725–732. [Google Scholar] [CrossRef]
- Hayashi, K.; Fukushima, D.; Mogi, K. Isolation of alkaline proteinase from Aspergillus sojae in homogeneous form. Agric. Biol. Chem. 1967, 31, 1237–1241. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cation (10 mM) | Relative Activity (%) |
|---|---|
| Control | 100 |
| Ca+2 | 63 ± 4.2 |
| K+1 | 57 ± 0.5 |
| Mg+2 | 61 ± 3.2 |
| Zn+2 | 58 ± 1.9 |
| Mn+2 | 80 ± 10.8 |
| Fe+2 | 75 ± 5.5 |
| Cu+2 | 69 ± 6.6 |
| Co+2 | 68 ± 4.6 |
| Inhibitor | Concentration | Relative Activity (%) |
|---|---|---|
| Control | - | 100 |
| EDTA | 10 mM | 85 ± 3.7 |
| EGTA | 10 mM | 93 ± 1.8 |
| PMSF | 5 mM | 34 ± 1.6 |
| Aprotinin | 100 µg/mL | 82 ± 3.8 |
| Benzamidine-HCl | 50 mM | 83 ± 5.0 |
| Leupeptin | 50 µM | 76 ± 5.0 |
| Reagent | Concentration | Relative Activity (%) |
|---|---|---|
| Control | - | 100 |
| Urea | 7 M | 75 ± 1.0 |
| HCl-Guanidine | 6 M | 103 ± 4.2 |
| 2-Mercaptoethanol | 10 mM | 90 ± 5.3 |
| DTT * | 10 mM | 88 ± 5.5 |
| H2O2 | 1 mM | 90 ± 2.3 |
| Triton X-100 | 1 mM | 100 ± 4.4 |
| Tween 80 | 1 mM | 100 ± 3.0 |
| SDS * | 1 mM | 84 ± 1.0 |
| Organic Solvent (0.5 mM) | Residual Activity (%) |
|---|---|
| Control | 100 |
| Methanol | 74 ± 6.8 |
| Ethanol | 70 ± 9.2 |
| 2-propanol | 57 ± 3.2 |
| Isoamyl alcohol | 79 ± 3.7 |
| Hexane | 96 ± 1.8 |
| Chloroform | 100 ± 11.2 |
| Acetone | 93 ± 0.3 |
| DMSO * | 98 ± 4.7 |
| Xylene | 102 ± 4.2 |
| Detergents 1% (v/v) | Residual Activity (%) |
|---|---|
| Control | 100 |
| D1 (OMO) | 88 ± 2.4 |
| D2 (ACE) | 93 ± 3.0 |
| D3 (QUIX) | 94 ± 3.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, B.; Solís-Cornejo, F.; Araya, R.; Zamorano, P. Isolation and Characterization of Thermus thermophilus Strain ET-1: An Extremely Thermophilic Bacterium with Extracellular Thermostable Proteolytic Activity Isolated from El Tatio Geothermal Field, Antofagasta, Chile. Int. J. Mol. Sci. 2023, 24, 14512. https://doi.org/10.3390/ijms241914512
Valenzuela B, Solís-Cornejo F, Araya R, Zamorano P. Isolation and Characterization of Thermus thermophilus Strain ET-1: An Extremely Thermophilic Bacterium with Extracellular Thermostable Proteolytic Activity Isolated from El Tatio Geothermal Field, Antofagasta, Chile. International Journal of Molecular Sciences. 2023; 24(19):14512. https://doi.org/10.3390/ijms241914512
Chicago/Turabian StyleValenzuela, Bernardita, Francisco Solís-Cornejo, Rubén Araya, and Pedro Zamorano. 2023. "Isolation and Characterization of Thermus thermophilus Strain ET-1: An Extremely Thermophilic Bacterium with Extracellular Thermostable Proteolytic Activity Isolated from El Tatio Geothermal Field, Antofagasta, Chile" International Journal of Molecular Sciences 24, no. 19: 14512. https://doi.org/10.3390/ijms241914512
APA StyleValenzuela, B., Solís-Cornejo, F., Araya, R., & Zamorano, P. (2023). Isolation and Characterization of Thermus thermophilus Strain ET-1: An Extremely Thermophilic Bacterium with Extracellular Thermostable Proteolytic Activity Isolated from El Tatio Geothermal Field, Antofagasta, Chile. International Journal of Molecular Sciences, 24(19), 14512. https://doi.org/10.3390/ijms241914512