
miR-30a-3p Regulates Autophagy in the Involution of Mice Mammary Glands

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
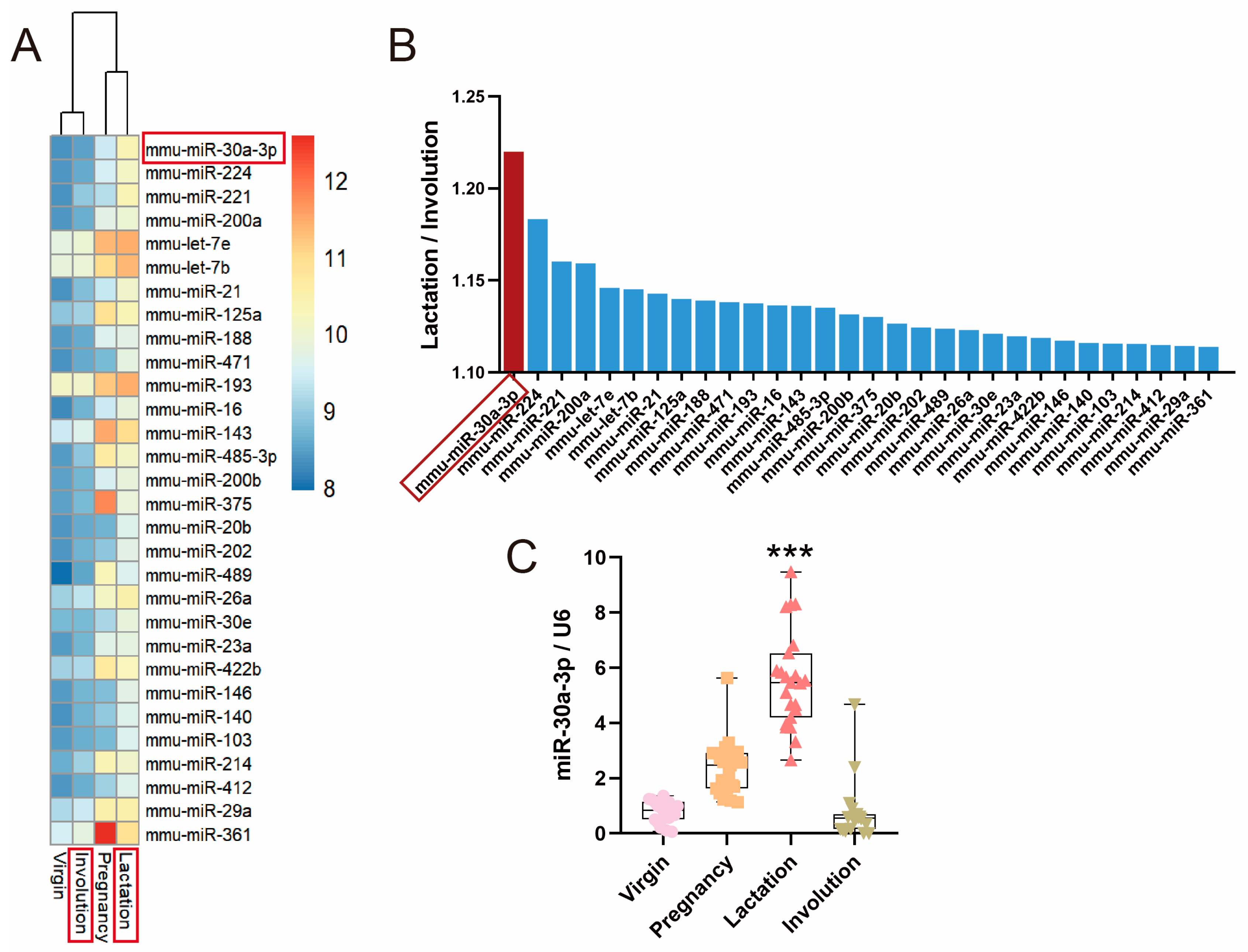
2.1. miR-30a-3p Was Downregulated during Involution Compared with Lactation in Mice Mammary Glands
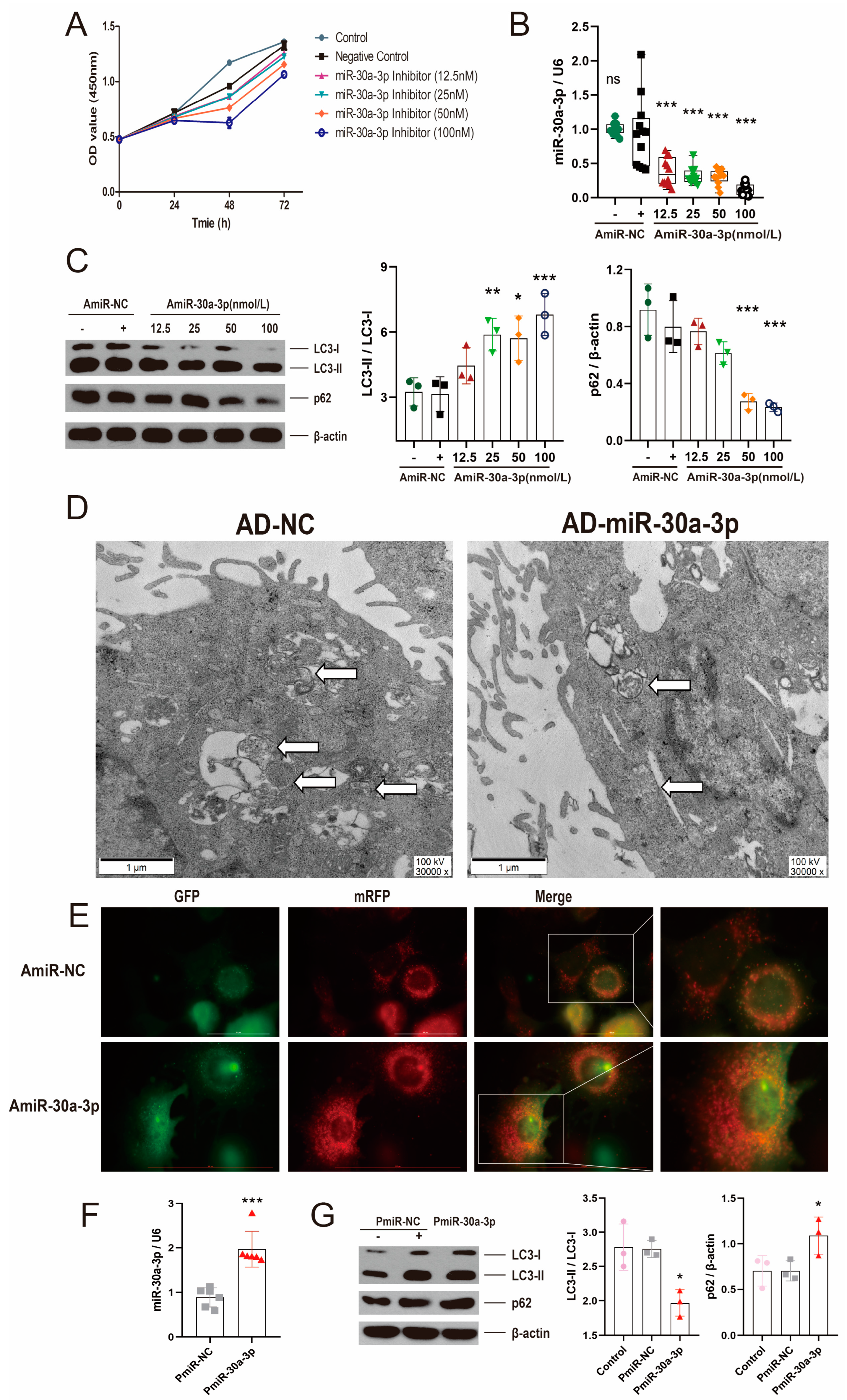
2.2. miR-30a-3p Negatively Regulated Autophagy in MMECs
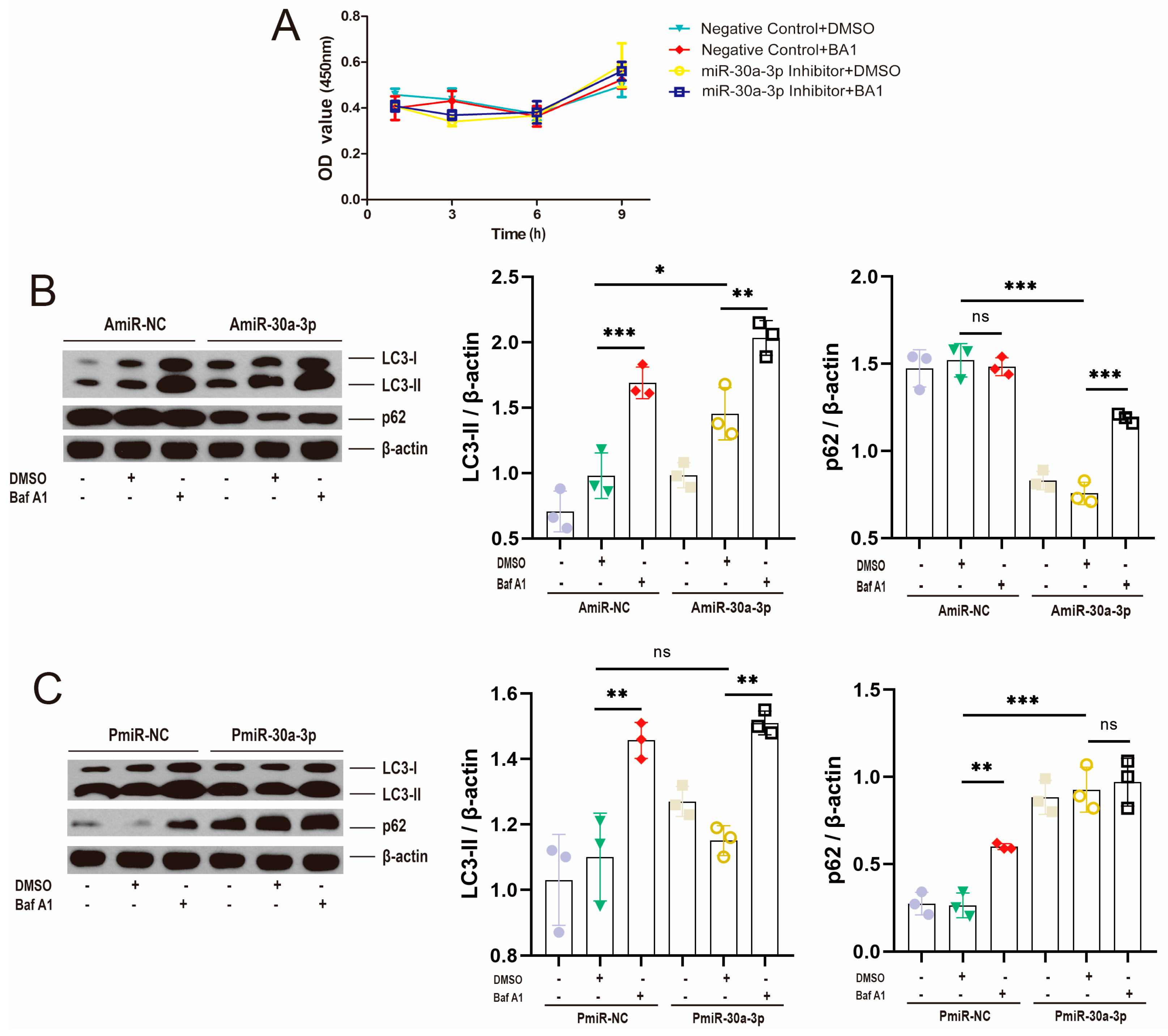
2.3. miR-30a-3p-Regulated Autophagy Verified by Bafilomycin A1 in MMECs
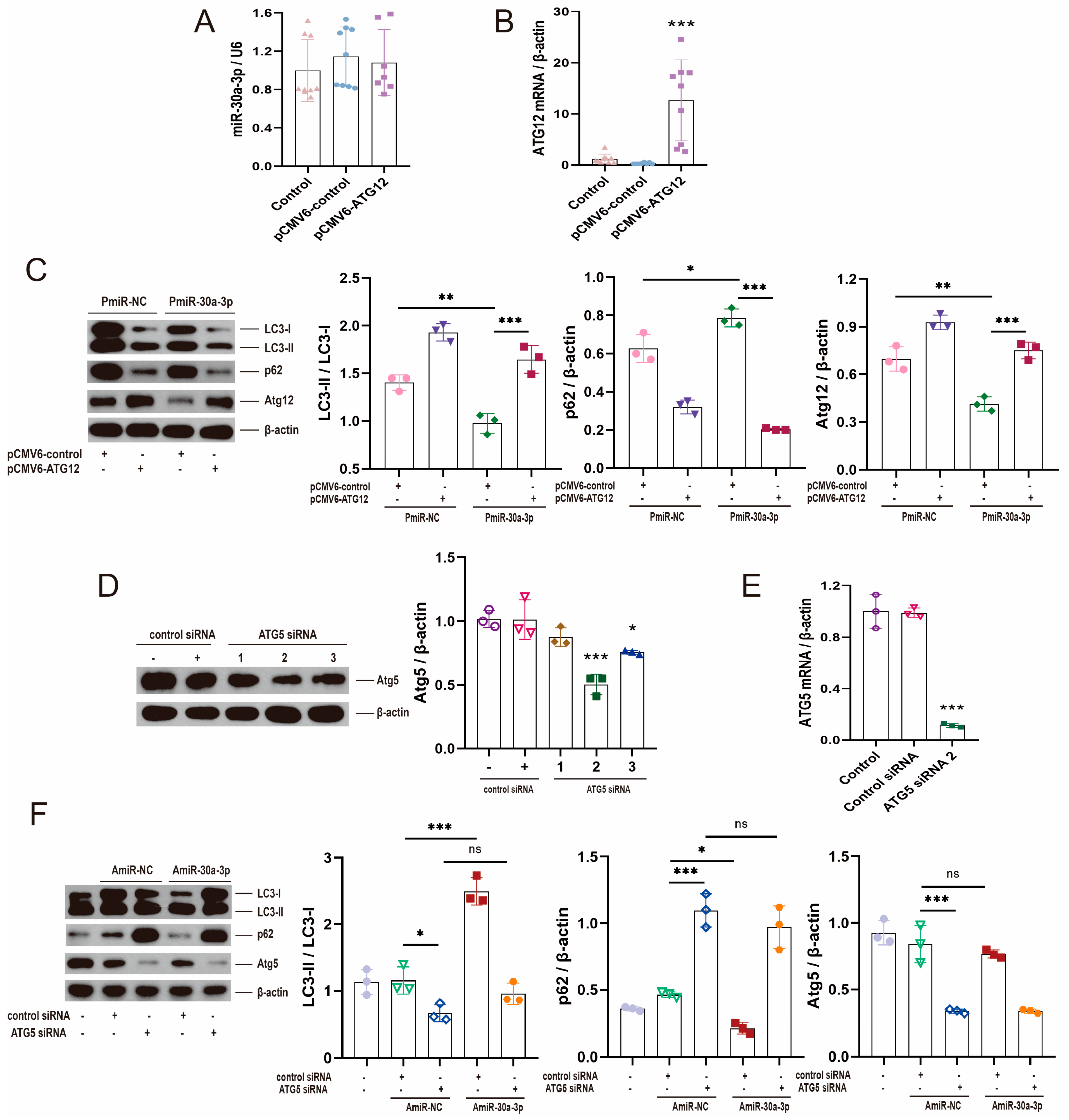
2.4. miR-30a-3p Might Inhibit Autophagy in MMECs by Atg12
2.5. Restoring ATG12 and Knocking down ATG5 Were Used to Validate That miR-30a-3p Inhibited Autophagy in MMECs by Atg12
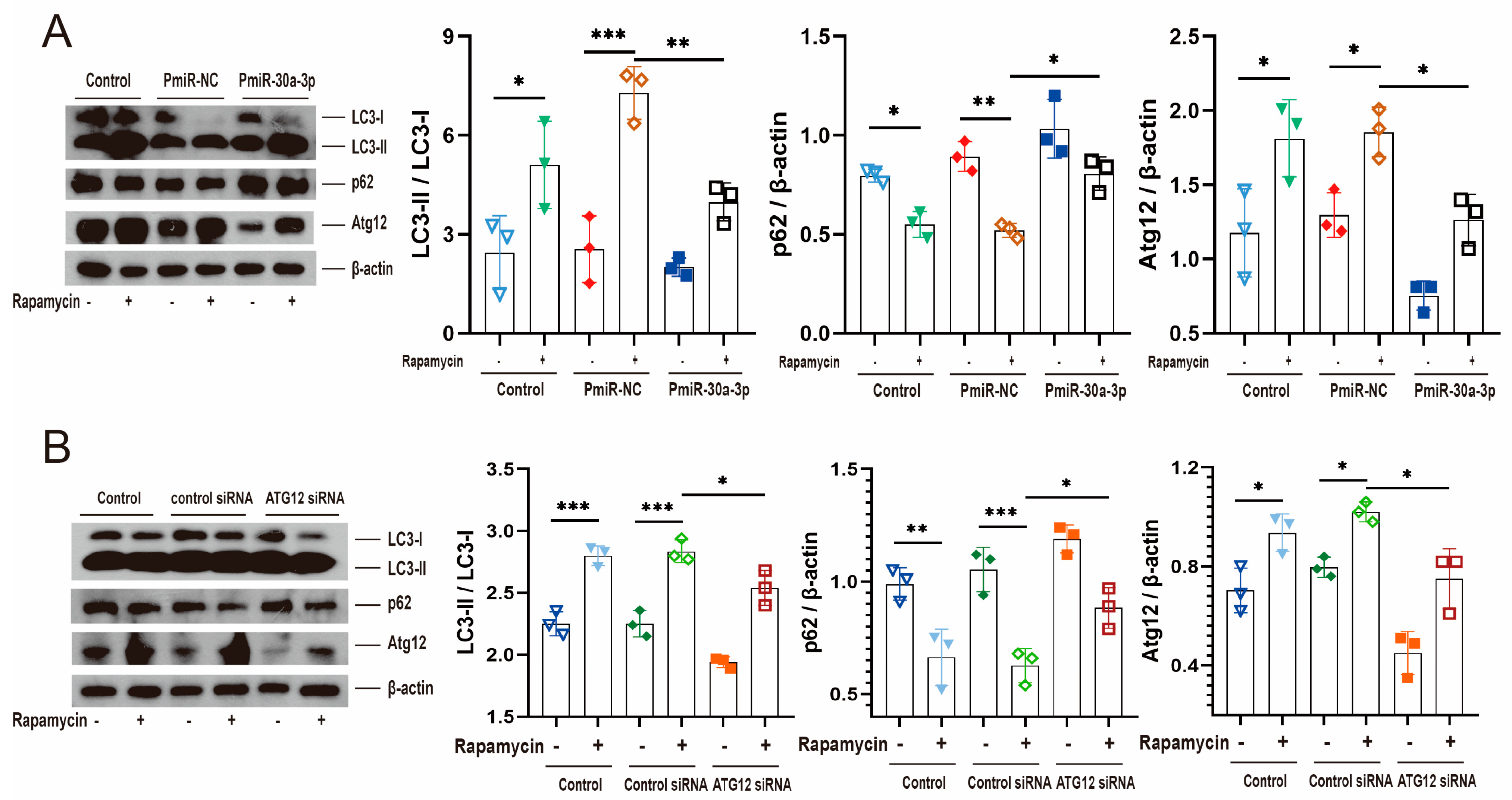
2.6. miR-30a-3p’s Regulation of Autophagy by ATG12 Was Verified through Starvation and Rapamycin In Vitro
2.7. Overexpression of miR-30a-3p Inhibited Autophagy and Delayed Mice Mammary Involution
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cell Culture
4.3. Quantitative Real-Time PCR (RT-PCR) Analyses
4.4. Cell Viability Assay
4.5. Western Blot Analysis
4.6. Vectors
4.7. mRFP-GFP-LC3 Assay
4.8. Immunofluorescence, Hematoxylin, and Eosin Staining
4.9. TEM
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Oftedal, O.T. The origin of lactation as a water source for parchment-shelled eggs. J. Mammary Gland Biol. Neoplasia 2002, 7, 253–266. [Google Scholar] [CrossRef]
- Slepicka, P.F.; Somasundara, A.V.H.; dos Santos, C.O. The molecular basis of mammary gland development and epthelial differentiation. Semin. Cell Dev. Biol. 2021, 114, 93–112. [Google Scholar] [CrossRef] [PubMed]
- Jindal, S.; Gao, D.; Bell, P.; Albrektsen, G.; Edgerton, S.M.; Ambrosone, C.B.; Thor, A.D.; Borges, V.F.; Schedin, P. Postpartum breast involution reveals regression of secretory lobules mediated by tissue-remodeling. Breast Cancer Res. 2014, 16, R31. [Google Scholar] [CrossRef]
- Sharp, J.A.; Lefevre, C.; Brennan, A.J.; Nicholas, K.R. The fur seal-a model lactation phenotype to explore molecular factors involved in the initiation of apoptosis at involution. J. Mammary Gland Biol. Neoplasia 2007, 12, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; Seo, Y.A.; Sharma, S.; Soybel, D.I.; Kelleher, S.L. ZnT2 is a critical mediator of lysosomal-mediated cell death during early mammary gland involution. Sci. Rep. 2015, 5, 8033. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.L.; Or, Y.Z.; Ong, Z.; Chung, H.H.; Hayashi, H.; Shrestha, S.; Chiba, S.; Lin, F.; Lin, V.C.L. Estrogen exacerbates mammary involution through neutrophil-dependent and -independent mechanism. eLife 2020, 9, e57274. [Google Scholar] [CrossRef]
- Teplova, I.; Lozy, F.; Price, S.; Singh, S.; Barnard, N.; Cardiff, R.D.; Birge, R.B.; Karantza, V. ATG proteins mediate efferocytosis and suppress inflammation in mammary involution. Autophagy 2013, 9, 459–475. [Google Scholar] [CrossRef]
- Hu, B.; Song, W.; Tang, Y.; Shi, M.; Li, H.; Yu, D. Induction of chemerin on autophagy and apoptosis in dairy cow mammary epithelial cells. Animals 2019, 9, 848. [Google Scholar] [CrossRef]
- Wärri, A.; Cook, K.L.; Hu, R.; Jin, L.; Zwart, A.; Soto-Pantoja, D.R.; Liu, J.; Finkel, T.; Clarke, R. Autophagy and unfolded protein response (UPR) regulate mammary gland involution by restraining apoptosis-driven irreversible changes. Cell Death Discov. 2018, 4, 40. [Google Scholar] [CrossRef]
- Fabris, T.F.; Laporta, J.; Skibiel, A.L.; Dado-Senn, B.; Wohlgemuth, S.E.; Dahl, G.E. Effect of heat stress during the early and late dry period on mammary gland development of Holstein dairy cattle. J. Dairy Sci. 2020, 103, 8576–8586. [Google Scholar] [CrossRef]
- Pensa, S.; Lloyd-Lewis, B.; Sargeant, T.J.; Resemann, H.K.; Kahn, C.R.; Watson, C.J. Signal transducer and activator of transcription 3 and the phosphatidylinositol 3-kinase regulatory subunits p55α and p50α regulate autophagy in vivo. FEBS J. 2014, 281, 4557–4567. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, Q. Identification of Differentially expressed microRNAs during the development of Chinese murine mammary gland. J. Genet. Genom. 2007, 34, 966–973. [Google Scholar] [CrossRef]
- Yang, B.; Jiao, B.; Ge, W.; Zhang, X.; Wang, S.; Zhao, H.; Wang, X. Transcriptome sequencing to detect the potential role of long non-coding RNAs in bovine mammary gland during the dry and lactation period. BMC Genom. 2018, 19, 605. [Google Scholar] [CrossRef]
- Lyu, M.; Li, F.; Wang, X.; Xu, K.; Sun, S. miR-145 modulates fatty acid metabolism by targeting FOXO1 to affect SERBP1 activity in bovine mammary epithelial cells. J. Agric. Food Chem. 2023, 71, 7440–7450. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Li, Y.; Wang, C.; Li, Q. Let-7g-5p regulates mouse mammary cells differentiation and function by targeting PRKCA. J. Cell. Physiol. 2019, 234, 10101–10110. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhang, L.; Cui, Y.; Li, H.; Xie, X.; Li, Y.; Wang, C. miR-142-3p regulates milk synthesis and structure of murine mammary glands via PRLR-Mediated multiple signaling pathways. J. Agric. Food Chem. 2019, 67, 9532–9542. [Google Scholar] [CrossRef]
- Yamamoto, A.; Tagawa, Y.; Yoshimori, T.; Moriyama, Y.; Masaki, R.; Tashiro, Y. Bafilomycin A1 prevents maturation of autophagic vacuoles by inhibiting fusion between autophagosomes and lysosomes in rat hepatoma cell line, H-4-II-E cells. Cell Struct. Funct. 1998, 23, 33–42. [Google Scholar] [CrossRef]
- Li, M.; Liu, X.; Robinson, G.; Bar-Peled, U.; Wagner, K.U.; Young, W.S.; Hennighausen, L.; Furth, P.A. Mammary-derived signals activate programmed cell death during the first stage of mammary gland involution. Proc. Natl. Acad. Sci. USA 1997, 94, 3425–3430. [Google Scholar] [CrossRef]
- Akhtar, N.; Li, W.; Mironov, A.; Streuli, C.H. Rac1 controls both the secretory function of the mammary gland and its remodeling for successive gestations. Dev. Cell 2016, 38, 522–535. [Google Scholar] [CrossRef]
- Jaswal, S.; Jena, M.K.; Anand, V.; Jaswal, A.; Kancharla, S.; Kolli, P.; Mandadapu, G.; Kumar, S.; Mohanty, A.K. Critcal review on physiological and molecular features during bovine mammary gland development: Recent advances. Cells 2022, 11, 3325. [Google Scholar] [CrossRef]
- Lyons, T.R.; O’Brien, J.; Borges, V.F.; Conklin, M.W.; Keely, P.J.; Eliceiri, K.W.; Marusyk, A.; Tan, A.C.; Schedin, P. Postpartum mammary gland involution drives progression of ductal carcinoma in situ through collagen and COX-2. Nat. Med. 2011, 17, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Crump, L.S.; Kines, K.T.; Richer, J.K.; Lyons, T.R. Breast cancers co-opt normal mechanisms of tolerance to promote immune evasion and metastasis. Am. J. Physiol. Cell Physiol. 2022, 323, C1475–C1495. [Google Scholar] [CrossRef]
- Mironov, A.; Fisher, M.; Narayanan, P.; Elsayed, R.; Karabulutoglu, M.; Akhtar, N. Rac1 controls cell turnover and reversibility of the involution process in postpartum mammary glands. PLoS Biol. 2023, 21, e3001583. [Google Scholar] [CrossRef] [PubMed]
- Ewan, K.B.R.; Oketch-Rabah, H.A.; Ravani, S.A.; Shyamala, G.; Moses, H.L.; Barcellos-Hoff, M.H. Proliferation of etrogen receptor-α-positive mammary epithelial cells is restrained by transforming growth factor-β1 in adult mice. Am. J. Pathol. 2005, 167, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Iwasaka, T.; Umemura, S.; Kakimoto, K.; Koizumi, H.; Osamura, Y.R. Expression of prolactin mRNA in rat mammary gland during pregnancy and lactation. J. Histochem. Cytochem. 2000, 48, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Vetharaniam, I.; Dobson, J.M.; Prewitz, M.; Oden, K.; Murney, R.; Swanson, K.M.; McDonald, R.; Henderson, H.V.; Stelwagen, K. Cell survival signaling in the bovine mammary gland during the transition from lactation to involution. J. Dairy Sci. 2016, 99, 7523–7543. [Google Scholar] [CrossRef]
- Singh, S.; Choudhary, S.; Anand, V.; Jaswal, S.; Verma, A.K.; Kumar, S.; Kaushik, J.K.; Mohanty, A.K. New insights into the catalytic inactivity of mammary gland protein-40, a chitinase-like protein expressed during mammary gland involution. Mol. Biol. Rep. 2019, 46, 2243–2257. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in development and disease: Background, mechanisms, and therapeutic approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef]
- Tanigawa, K.; Misono, S.; Mizuno, K.; Asai, S.; Suetsugu, T.; Uchida, A.; Kawano, M.; Inoue, H.; Seki, N. MicroRNA signature of small-cell lung cancer after treatment failure: Impact on oncogenic targets by miR-30a-3p control. Mol. Oncol. 2023, 17, 328–343. [Google Scholar] [CrossRef]
- Wang, H.; Kanmangne, D.; Li, R.; Qian, Z.; Xia, X.; Wang, X.; Wang, T. miR-30a-3p suppresses the proliferation and migration of lung adenocarcinoma cells by downregulating CNPY2. Oncol. Rep. 2020, 43, 646–654. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wu, X.; Huang, J.; Chen, W.; Liu, D.; Zhang, J.; Huang, Y.; Xue, W. miR-30a-3p inhibits renal cancer cell invasion and metastasis through targeting ATG12. Transl. Androl. Urol. 2020, 9, 646–653. [Google Scholar] [CrossRef]
- Feng, L.; Jing, W.; Jin, S.; Wang, B. Circ_0088194 regulates proliferation, migration, apoptosis, and inflammation by miR-30a-3p/ADAM10 axis in rheumatoid arthritis fibroblastic synovial cells. Inflammation 2023, 46, 161–174. [Google Scholar] [CrossRef]
- Hwang, T.I.S.; Chen, P.C.; Tsai, T.F.; Lin, J.F.; Chou, K.Y.; Ho, C.Y.; Chen, H.E.; Chang, A.C. Hsa-miR-30a-3p overcomes the acquired protective autophagy of bladder cancer in chemotherapy and suppresses tumor growth and muscle invasion. Cell Death Dis. 2022, 13, 390. [Google Scholar] [CrossRef]
- Mizushima, N. The ATG conjugation systems in autophagy. Curr. Opin. Cell Biol. 2020, 63, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Walczak, M.; Martens, S. Dissecting the role of the Atg12-Atg5-Atg16 complex during autophagosome formation. Atophagy 2013, 9, 424–425. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Chen, S.; Du, F.; Li, S.; Zhao, L.; Wang, X. Nutrient starvation elicits an acute autophagic response mediated by Ulk1 dephosphorylation and its subsequent dissociation from AMPK. Proc. Natl. Acad. Sci. USA 2011, 108, 4788–4793. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.A.; Biggers, C.D.; Li, P.A. Rapamycin treatment increases hippocampal cell viability in an mTOR-independent manner during exposure to hypoxia mimetic, cobalt chloride. BMC Neurosci. 2018, 19, 82. [Google Scholar] [CrossRef]
- Hannan, F.M.; Elajnaf, T.; Vandenberg, L.N.; Kennedy, S.H.; Thakker, R.V. Hormonal regulation of mammary gland development and lactation. Nat. Rev. Endocrinol. 2023, 19, 46–61. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, L.; Guo, S.; Zhao, Z.; Chen, Y.; Wang, C.; Li, Q.; Li, Y. miR-30a-3p Regulates Autophagy in the Involution of Mice Mammary Glands. Int. J. Mol. Sci. 2023, 24, 14352. https://doi.org/10.3390/ijms241814352
Tian L, Guo S, Zhao Z, Chen Y, Wang C, Li Q, Li Y. miR-30a-3p Regulates Autophagy in the Involution of Mice Mammary Glands. International Journal of Molecular Sciences. 2023; 24(18):14352. https://doi.org/10.3390/ijms241814352
Chicago/Turabian StyleTian, Lei, Shancheng Guo, Zhiye Zhao, Yuxu Chen, Chunmei Wang, Qingzhang Li, and Ye Li. 2023. "miR-30a-3p Regulates Autophagy in the Involution of Mice Mammary Glands" International Journal of Molecular Sciences 24, no. 18: 14352. https://doi.org/10.3390/ijms241814352
APA StyleTian, L., Guo, S., Zhao, Z., Chen, Y., Wang, C., Li, Q., & Li, Y. (2023). miR-30a-3p Regulates Autophagy in the Involution of Mice Mammary Glands. International Journal of Molecular Sciences, 24(18), 14352. https://doi.org/10.3390/ijms241814352