
Cytotoxic Potential of Novel Quinoline Derivative: 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline against Different Cancer Cell Lines via Activation and Deactivation of the Expression of Some Proteins

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
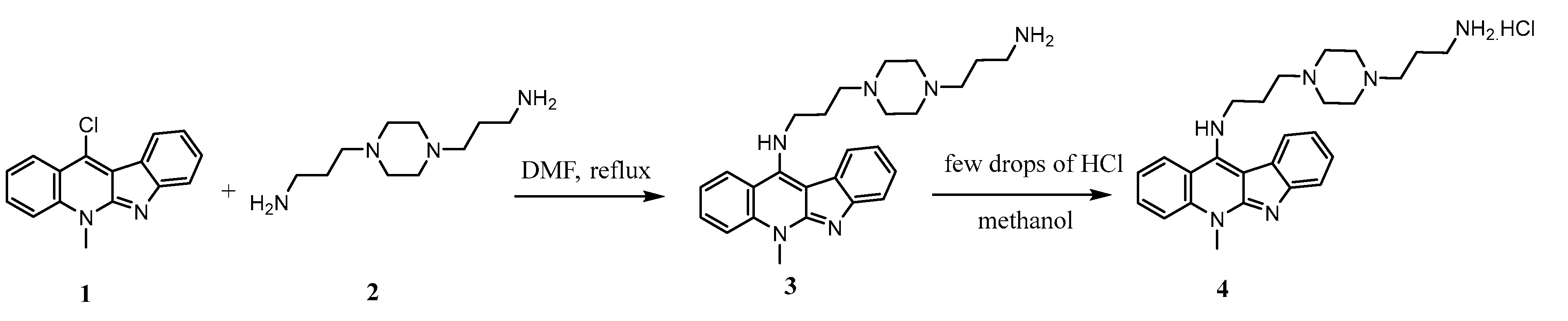
2.1. Synthesis of 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline 4 (BAPPN) and Its Structural Characterization by 1H NMR and 13C NMR
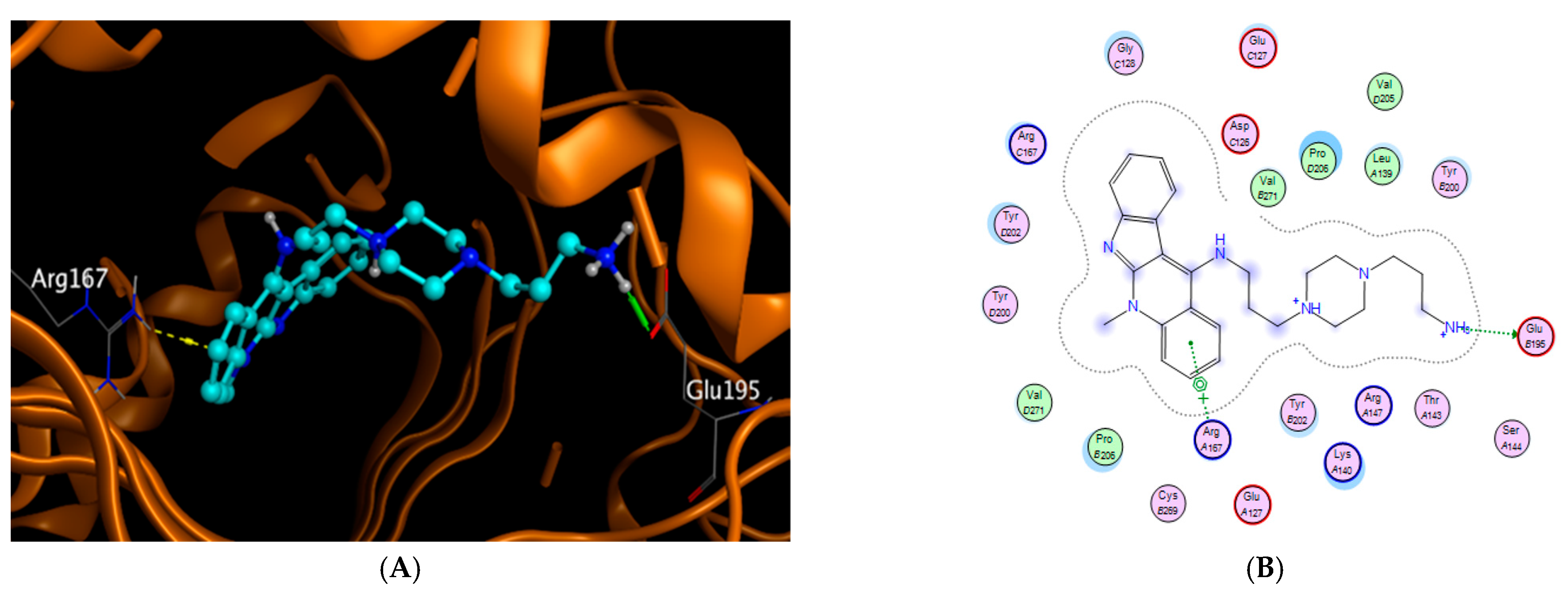
2.2. Modeling Studies
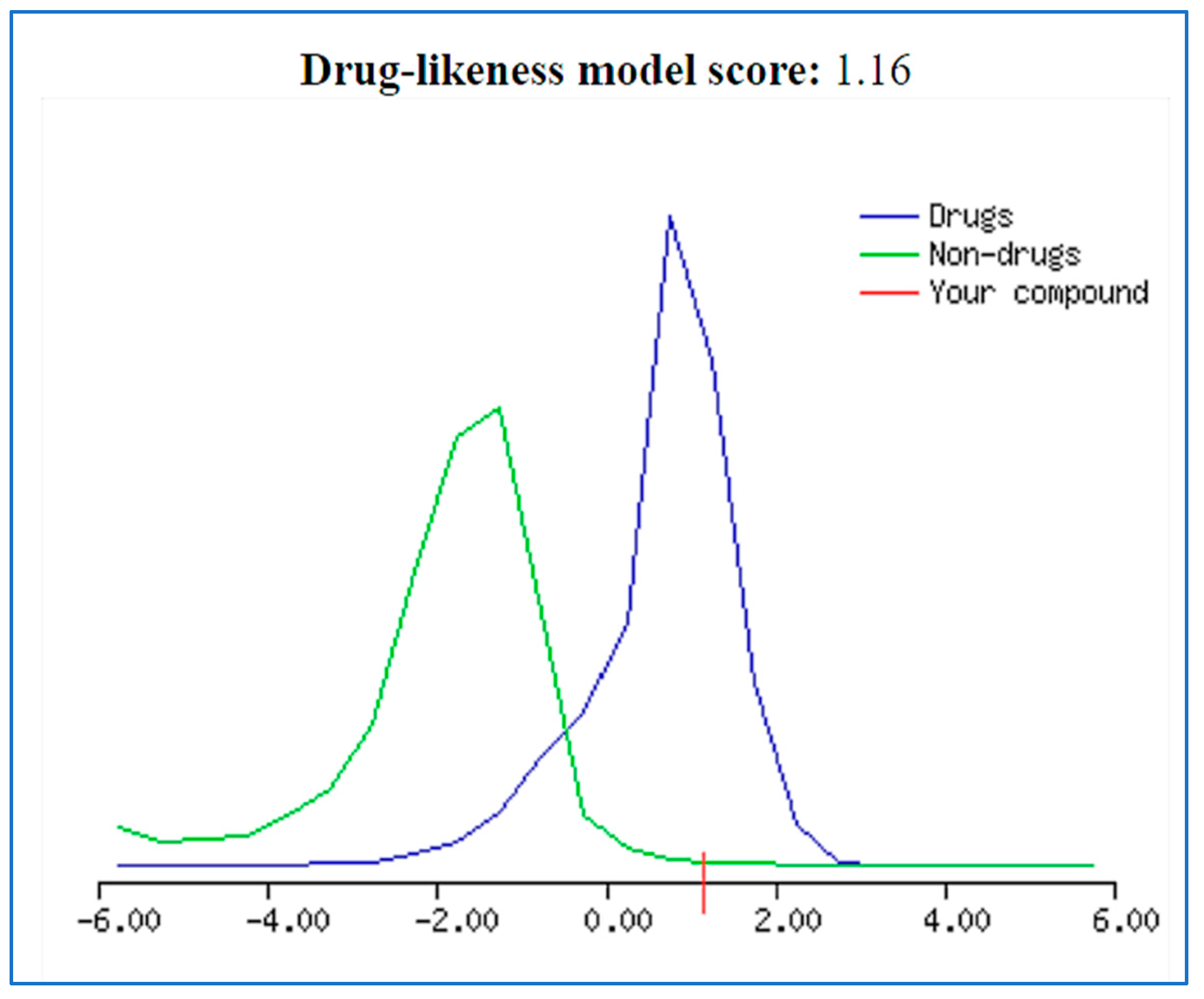
2.3. Bioavailability and ADME in Silico Studies
2.4. Drug Likeness Probability
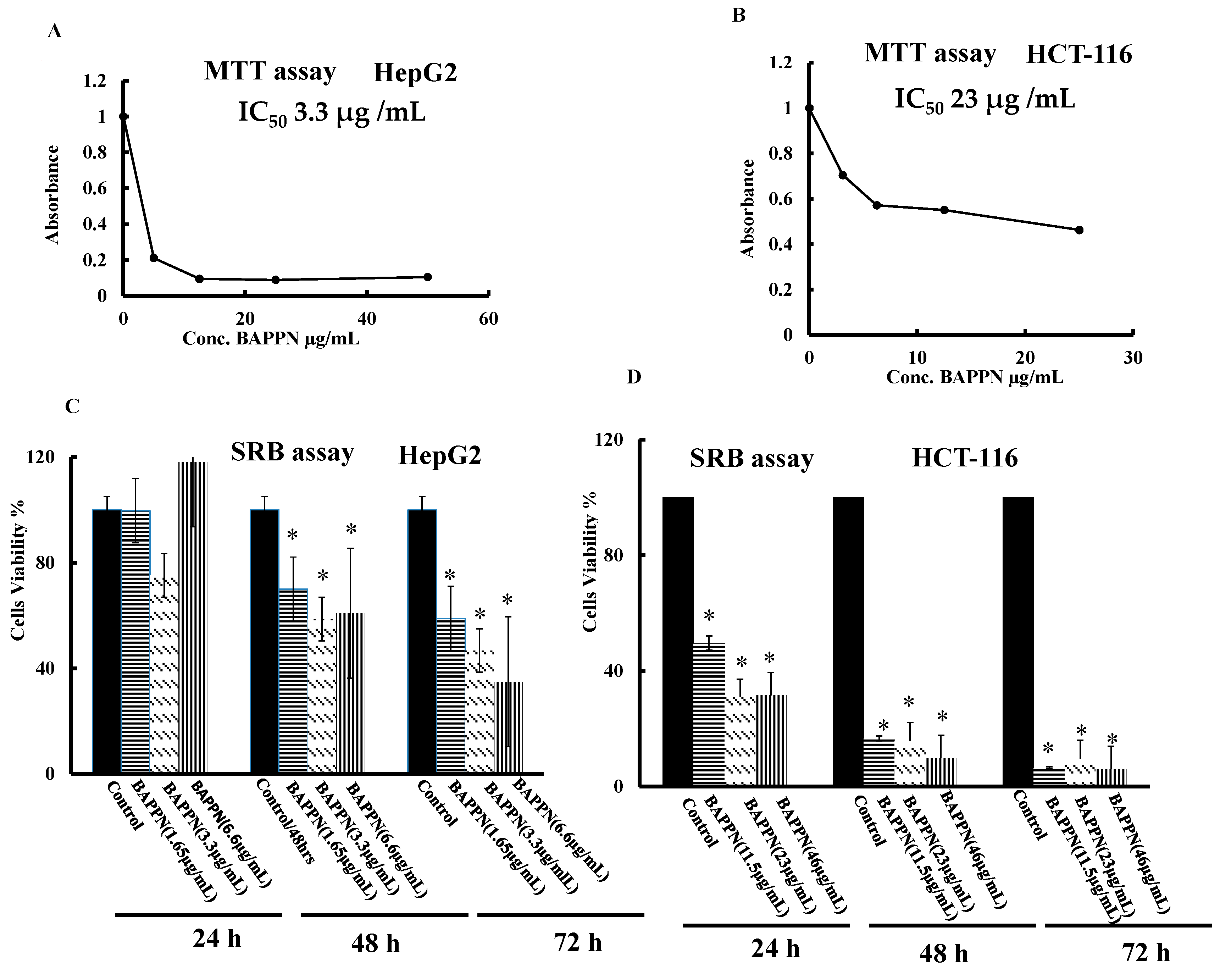
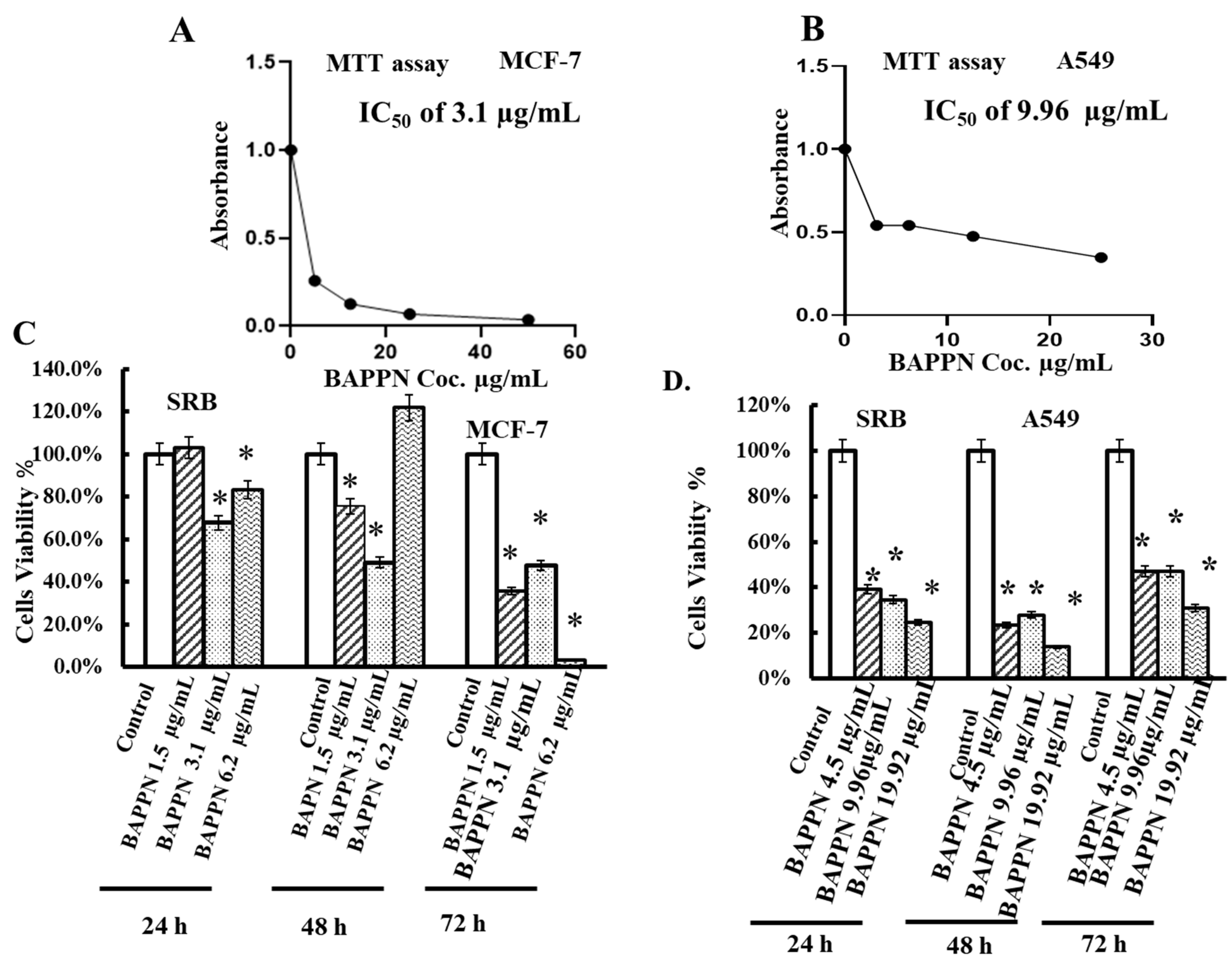
2.5. 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline Induced Cytotoxicity in Cancer Cell Lines
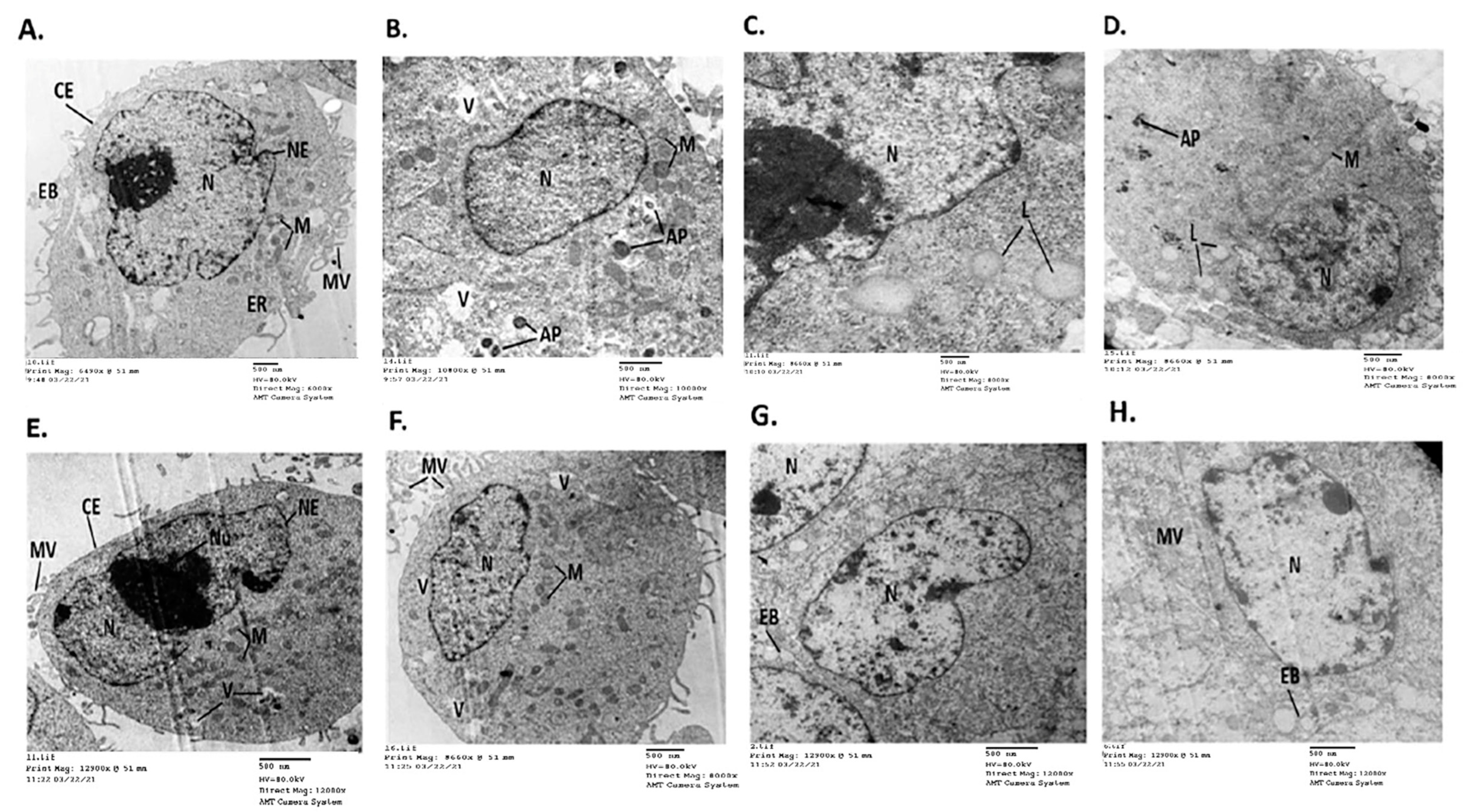
2.6. 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline Induced Detrimental Changes in the Ultrastructure of Cancer Cell Lines
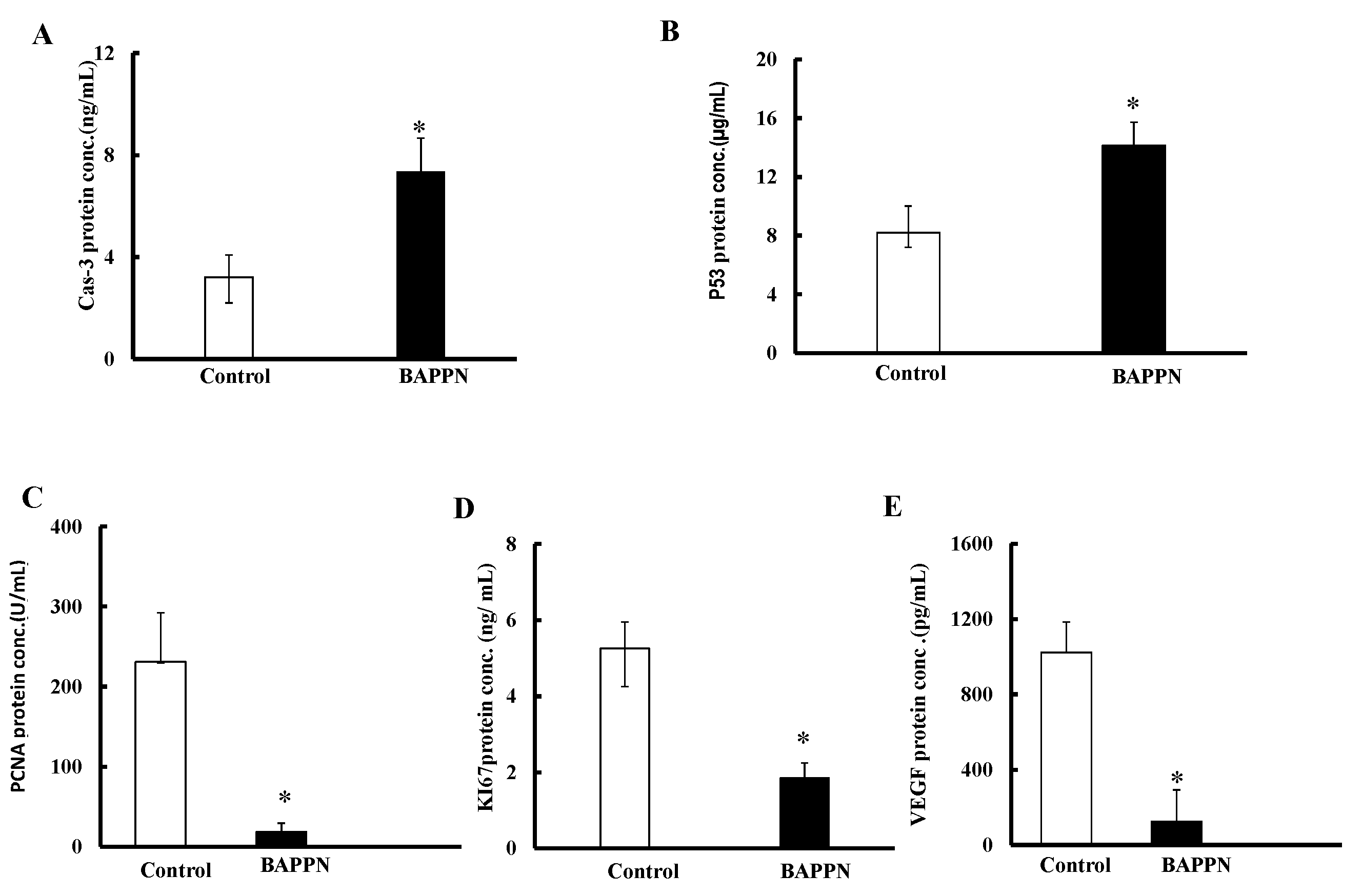
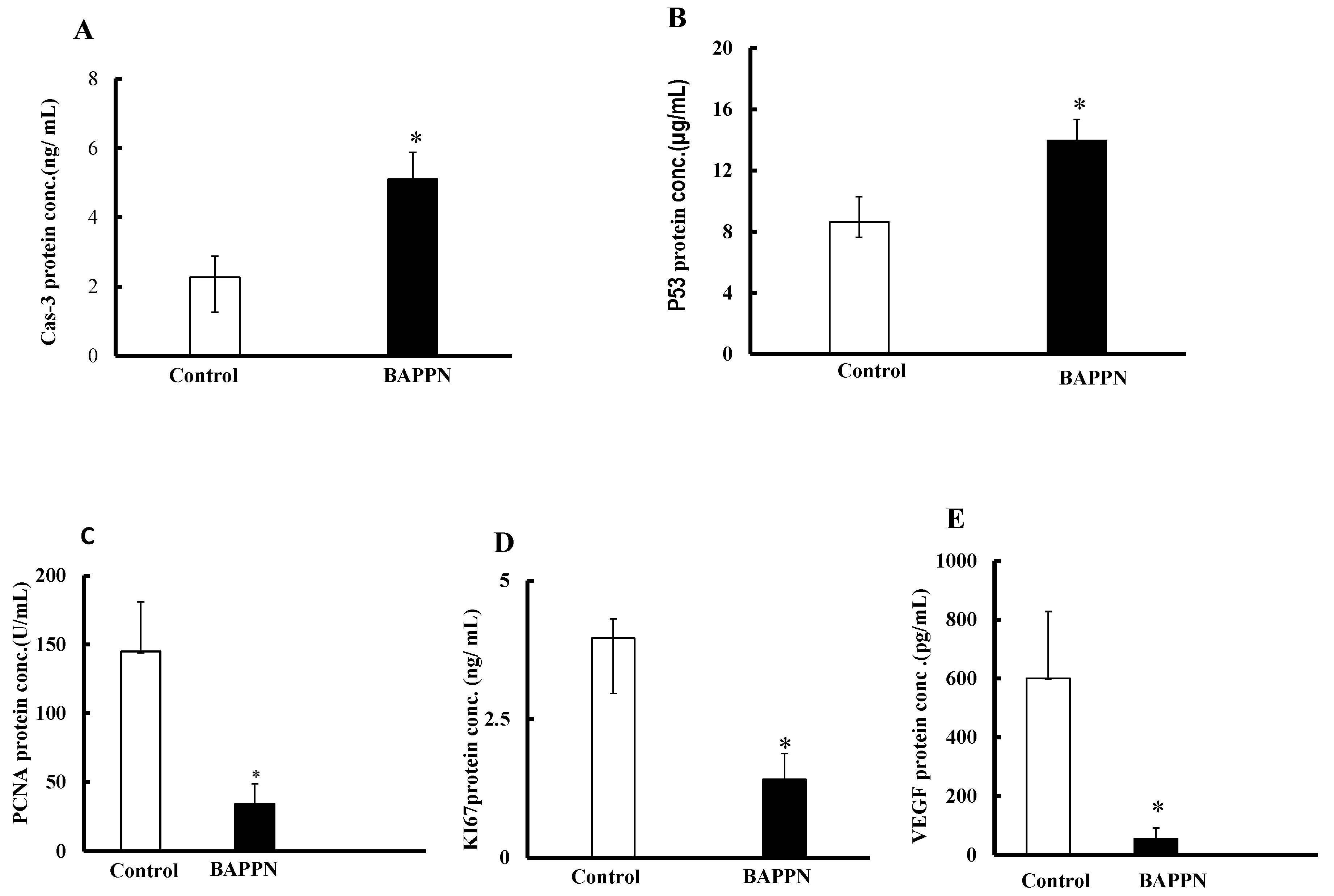
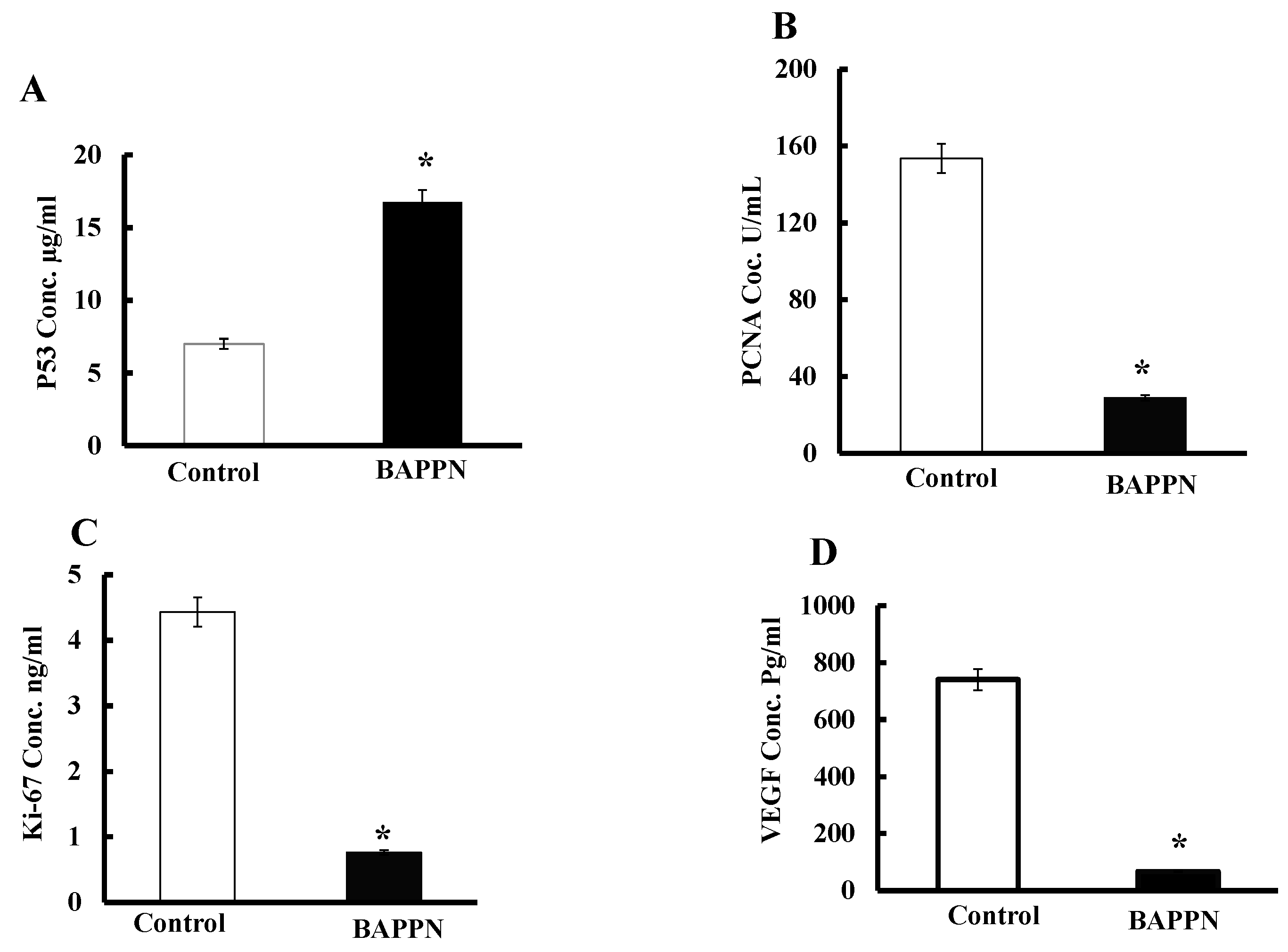
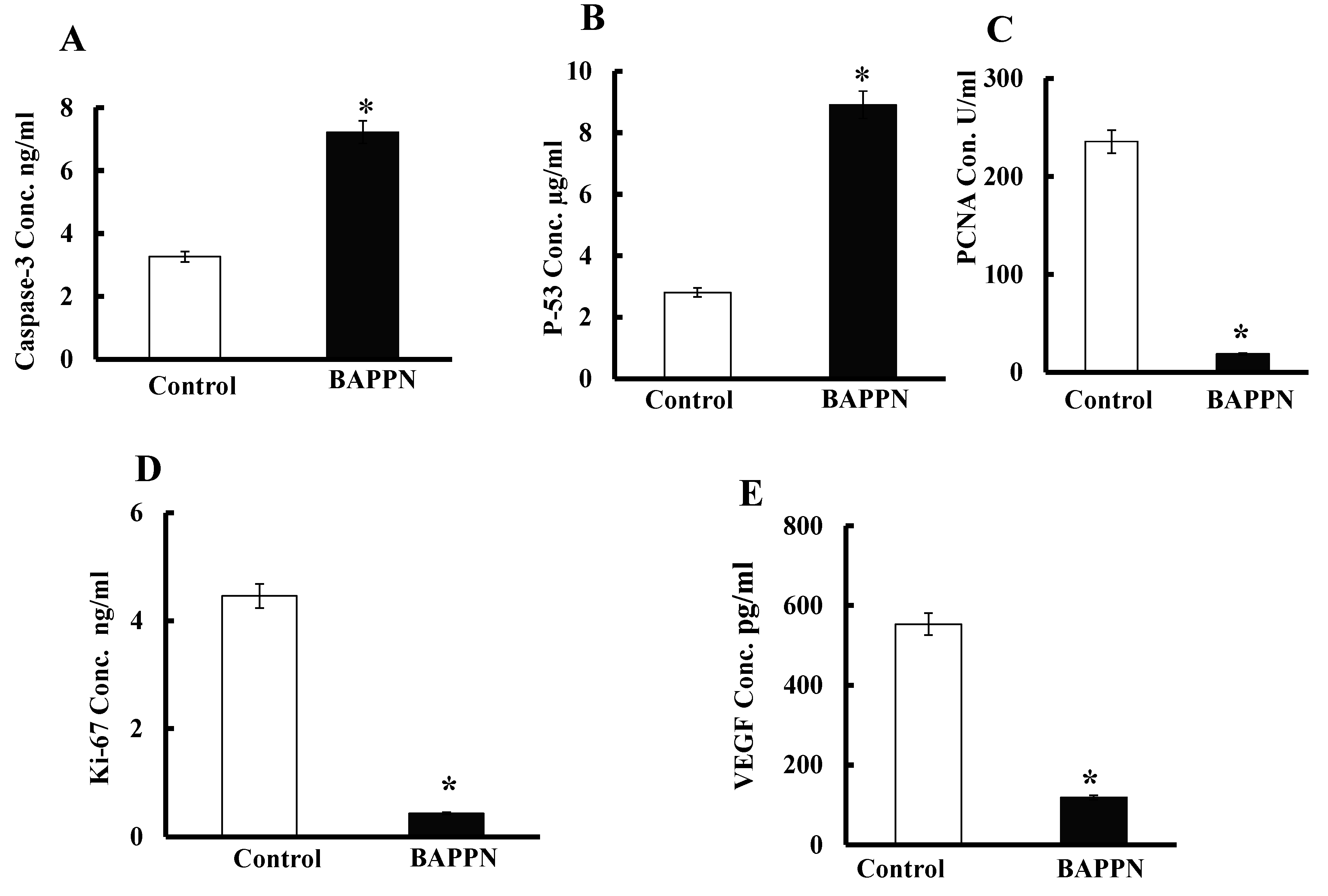
2.7. Effect of 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline on the Expression of Apoptotic and Proliferative Proteins in Cancer Cells
3. Discussion
4. Material and Methods
4.1. General
4.2. Preparation of N-(3-(4-(3-Aminopropyl)piperazin-1-Yl)propyl)-5-methyl-5H-indolo[2,3-b]quinolin-11-amine hydrochloride) (BAPPN)
4.3. Modeling Studies
4.4. Pharmacokinetics in Silico-Screening
4.5. Cell Culture
4.6. Determination of Medium Inhibitory Concentration (IC50) by MTT Assay
4.7. Sulforhodamine B Assay (SRB Assay)
4.8. Measurement of the Expression of Cellular Proteins of PCNA, Ki67, P53, VEGF, and Caspase-3 by ELISA Assay
4.9. Transmission Electron Microscope
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaudhry, G.E.; Md Akim, A.; Sung, Y.Y.; Sifzizul, T.M.T. Cancer and apoptosis: The apoptotic activity of plant and marine natural products and their potential as targeted cancer therapeutics. Front. Pharmacol. 2022, 10, 842376. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Petrick, J.L.; McGlynn, K.A. The changing epidemiology of primary liver cancer. Curr. Epidemiol. Rep. 2019, 6, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Chidambaranathan-Reghupaty, S.; Fisher, P.B.; Sarkar, D. Hepatocellular carcinoma (HCC): Epidemiology, etiology and molecular classification. Adv. Cancer Res. 2021, 149, 1–61. [Google Scholar]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Gastroenterol. Rev. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Tariq, K.; Ghias, K. Colorectal cancer carcinogenesis: A review of mechanisms. Cancer Biol. Med. 2016, 13, 120. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer—Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies—An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Arnold, M.; Rutherford, M.J.; Bardot, A.; Ferlay, J.; Andersson, T.M.; Myklebust, T.Å.; Tervonen, H.; Thursfield, V.; Ransom, D.; Shack, L.; et al. Progress in cancer survival, mortality, and incidence in seven high-income countries 1995–2014 (ICBP SURVMARK-2): A population-based study. Lancet Oncol. 2019, 20, 1493–1505. [Google Scholar] [CrossRef]
- Debela, D.T.; Muzazu, S.G.; Heraro, K.D.; Ndalama, M.T.; Mesele, B.W.; Haile, D.C.; Kitui, S.K.; Manyazewal, T. New approaches and procedures for cancer treatment: Current perspectives. SAGE Open Med. 2021, 9, 20503121211034366. [Google Scholar] [CrossRef]
- Muthukutty, P.; Woo, H.Y.; Ragothaman, M.; Yoo, S.Y. Recent Advances in Cancer Immunotherapy Delivery Modalities. Pharmaceutics 2023, 15, 504. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.-T.; Nguyen, T.T.; Tien, N.L.B.; Tran, D.-K.; Jeong, J.-H.; Anh, P.G.; Thanh, V.V.; Truong, D.T.; Dinh, T.C. Recent Progress of Stem Cell Therapy in Cancer Treatment: Molecular Mechanisms and Potential Applications. Cells 2020, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-W.; Wu, Y.-S.; Xu, T.-M.; Cui, M.-H. CAR-T Cells in the Treatment of Ovarian Cancer: A Promising Cell Therapy. Biomolecules 2023, 13, 465. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, G.; Zheng, L.; Long, H.; Liu, Y. A new era of gene and cell therapy for cancer: A narrative review. Ann. Transl. Med. 2023, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Min, H.Y.; Lee, H.Y. Molecular targeted therapy for anticancer treatment. Exp. Mol. Med. 2022, 54, 1670–1694. [Google Scholar] [CrossRef]
- Al-Zoubi, M.S.; Al-Zoubi, R.M. Nanomedicine tactics in cancer treatment: Challenge and hope. Crit. Rev. Oncol. Hematol. 2022, 174, 103677. [Google Scholar] [CrossRef]
- Fulton, M.D.; Najahi-Missaoui, W. Liposomes in Cancer Therapy: How Did We Start and Where Are We Now. Int. J. Mol. Sci. 2023, 24, 6615. [Google Scholar] [CrossRef]
- Iacopetta, D.; Ceramella, J.; Baldino, N.; Sinicropi, M.S.; Catalano, A. Targeting Breast Cancer: An Overlook on Current Strategies. Int. J. Mol. Sci. 2023, 24, 3643. [Google Scholar] [CrossRef]
- Waarts, M.R.; Stonestrom, A.J.; Park, Y.C.; Levine, R.L. Targeting mutations in cancer. J. Clin. Invest. 2022, 132, e154943. [Google Scholar] [CrossRef]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Larghi, E.L.; Bracca, A.B.; Arroyo Aguilar, A.A.; Heredia, D.A.; Pergomet, J.L.; Simonetti, S.O.; Kaufman, T.S. Neocryptolepine: A Promising Indoloisoquinoline Alkaloid with Interesting Biological Activity. Evaluation of the Drug and its Most Relevant Analogs. Curr. Top. Med. Chem. 2015, 15, 1683–1707. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wicht, K.J.; Wang, L.; Lu, W.J.; Misumi, R.; Wang, M.Q.; El Gokha, A.A.; Kaiser, M.; El Sayed, I.E.T.; Egan, T.J.; et al. Synthesis and in vitro testing of antimalarial activity of non-natural-type neocryptolepines: Structure–activity relationship study of 2,11- and 9,11-disubstituted 6-methylindolo[2,3-b]quinolines. Chem. Pharm. Bull. 2013, 6, 1282–1290. [Google Scholar] [CrossRef]
- Akkachairin, B.; Rodphon, W.; Reamtong, O.; Mungthin, M.; Tummatorn, J.; Thongsornkleeb, C.; Ruchirawat, S. Synthesis of neocryptolepines and carbocycle-fused quinolines and evaluation of their anticancer and antiplasmodial activities. Bioorg. Chem. 2020, 98, 103732. [Google Scholar] [CrossRef] [PubMed]
- Amin, B.H.; Ahmed, H.Y.; Abd El-Aziz, M.M. In vitro anticancer activity of fungal secondary metabolites of stemphylium lycopersici. N. Egypt. J. Microbiol. 2018, 50, 80–97. [Google Scholar]
- Pal, H.C.; Katiyar, S.K. Cryptolepine, a Plant Alkaloid, Inhibits the Growth of Non-Melanoma Skin Cancer Cells through Inhibition of Topoisomerase and Induction of DNA Damage. Molecules 2016, 21, 1758. [Google Scholar] [CrossRef] [PubMed]
- Sebeka, A.A.H.; Osman, A.M.A.; El Sayed, I.E.T.; El Bahanasawy, M.; Tantawy, M.A. Synthesis and antiproliferative activity of novel neocryptolepine-hydrazides hybrids. J. Appl. Pharm. Sci. 2017, 7, 9. [Google Scholar]
- Wang, N.; Świtalska, M.; Wang, L.; Shaban, E.; Hossain, M.I.; El Sayed, I.E.T.; Wietrzyk, J.; Inokuchi, T. Structural Modifications of Nature-Inspired Indoloquinolines: A Mini Review of Their Potential Antiproliferative Activity. Molecules 2019, 24, 2121. [Google Scholar] [CrossRef]
- Ma, Y.; Tian, Y.; Zhou, Z.; Chen, S.; Du, K.; Zhang, H.; Jiang, X.; Lu, J.; Niu, Y.; Tu, L.; et al. Design, Synthesis and Biological Evaluation of Neocryptolepine Derivatives as Potential Anti-Gastric Cancer Agents. Int. J. Mol. Sci. 2022, 23, 11924. [Google Scholar] [CrossRef]
- Boddupally, P.V.; Hahn, S.; Beman, C.; De, B.; Brooks, T.A.; Gokhale, V.; Hurley, L.H. Anticancer activity and cellular repression of c-MYC by the G-quadruplex-stabilizing 11-piperazinylquindoline is not dependent on direct targeting of the G-quadruplex in the c-MYC promoter. J. Med. Chem. 2012, 55, 6076–6086. [Google Scholar] [CrossRef]
- Rodphon, W.; Laohapaisan, P.; Supantanapong, N.; Reamtong, O.; Ngiwsara, L.; Lirdprapamongkol, K.; Thongsornkleeb, C.; Khunnawutmanotham, N.; Tummatorn, J.; Svasti, J.; et al. Synthesis of Isocryptolepine-Triazole Adducts and Evaluation of Their Cytotoxic Activity. ChemMedChem 2021, 16, 3750–3762. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.; Li, Z.; Huag, Q.; Li, F.; Li, C.Y. Caspase-3 regulates the migration, invasion and metastasis of colon cancer cells. Int. J. Cancer 2018, 143, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Dassonneville, L.; Lansiaux, A.; Wattelet, A.; Wattez, N.; Mahieu, C.V.; Miert, S.; Pieters, L.; Bailly, C. Cytotoxicity and cell cycle effects of the plant alkaloids cryptolepine and neocryptolepine: Relation to drug-induced apoptosis. Eur. J. Pharmacol. 2000, 409, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Engeland, K. Cell cycle arrest through indirect transcriptional repression by p53: I have a DREAM. Cell Death Differ. 2018, 25, 114–132. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, T.; Nakagawara, A. Role of p53 in Cell Death and Human Cancers. Cancers 2011, 3, 994–1013. [Google Scholar] [CrossRef]
- Bologna-Molina, R.; Mosqueda-Taylor, A.; Molina-Frechero, N.; Mori-Estevez, A.D.; Sánchez-Acuña, G. Comparison of the value of PCNA and Ki-67 as markers of cell proliferation in ameloblastic tumors. Med. Oral Patol. Oral Cir. Bucal. 2013, 18, e174–e179. [Google Scholar] [CrossRef]
- Gerdes, J.; Schwab, U.; Lemke, H.; Stein, H. Production of a mouse monoclonal antibody reactive with a human nuclear antigen associated with cell proliferation. Int. J. Cancer 1983, 31, 13–20. [Google Scholar] [CrossRef]
- Inwald, E.C.; Klinkhammer-Schalke, M.; Hofstädter, F.; Zeman, F.; Koller, M.; Gerstenhauer, M.; Ortmann, O. Ki-67 is a prognostic parameter in breast cancer patients: Results of a large population-based cohort of a cancer registry. Breast Cancer Res. Treat. 2013, 139, 539–552. [Google Scholar] [CrossRef]
- Aspriţoiu, V.M.; Stoica, I.; Bleotu, C.; Diaconu, C.C. Epigenetic Regulation of Angiogenesis in Development and Tumors Progression: Potential Implications for Cancer Treatment. Front. Cell Dev. Biol. 2021, 9, 689962. [Google Scholar] [CrossRef]
- Yang, S.H.; Chien, C.M.; Su, J.C.; Chen, Y.L.; Chang, L.S.; Lin, S.R. Novel Indoloquinoline Derivative, IQDMA, Inhibits STAT5 Signaling Associated with Apoptosis in K562 Cells. J. Biochem. Mol. Toxicol. 2008, 22, 396–404. [Google Scholar] [CrossRef]
- Lu, W.-J.; Switalska, M.; Wang, L.; Yonezawa, M.; El-Sayed, I.E.-T.; Wietrzyk, J.; Inokuchi, T. In vitro antiproliferative activity of 11-aminoalkylamino-substituted 5 H-indolo [2, 3-b] quinolines; improving activity of neocryptolepines by installation of estersubstituent. Med. Chem. Res. 2013, 22, 4492–4504. [Google Scholar] [CrossRef]
- El Sayed, I.E.T.; Ullah, S.; Al-Hartomy, O.A.; Hasanein, A.M.; Ahmed, A.A.S.; Kahilo, K.A.; El-Naggar, M.E. Synthesis, Nanoformulations, and In Vitro Anticancer Activity of N-Substituted Side Chain Neocryptolepine Scaffolds. Molecules 2022, 27, 1024. [Google Scholar] [CrossRef] [PubMed]
- Chemical Computing Group. Molecular Operating Environment (MOE); Chemical Computing Group Inc.: Montreal, QC, Canada, 2014. [Google Scholar]
- Yao, L.; Swartz, P.; Hamilton, P.T.; Clark, A.C. Remodeling hydrogen bond interactions results in relaxed specificity of Caspase-3. Biosci. Rep. 2021, 41, BSR20203495. [Google Scholar] [CrossRef]
- Nagy, E.T.; Ahmed, A.A.S.; Elmongy, E.I.; El-Gendy, S.M.; Elmadbouh, I.; El Sayed, I.E.T.; Abd Eldaim, M.A.; El-Gokha, A.A. Design and cytotoxic evaluation via apoptotic and antiproliferative activity for novel 11(4-aminophenylamino)neocryptolepine on hepatocellular and colorectal cancer cells. Apoptosis 2023, 28, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Elmongy, E.I.; Henidi, H.A. In Silico Evaluation of a Promising Key Intermediate Thieno [2,3-d] Pyrimidine Derivative with Expected JAK2 Kinase Inhibitory Activity. Molbank 2022, 2022, M1352. [Google Scholar] [CrossRef]
- Ahmad Mir, S.; Paramita Mohanta, P.; Kumar Meher, R.; Baitharu, I.; Kumar Raval, M.; Kumar Behera, A.; Nayak, B. Structural insights into conformational stability and binding of thiazolo-[2,3-b] quinazolinone derivatives with EGFR-TKD and in-vitro study. Saudi J. Biol. Sci. 2022, 29, 103478. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. iLOGP: A Simple, Robust, and Efficient Description of n-Octanol/Water Partition Coefficient for Drug Design Using the GB/SA Approach. J. Chem. Inf. Model. 2014, 54, 3284–3301. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Marks, D.C.; Belov, L.; Davey, M.W.; Davey, R.A.; Kidman, A.D. The MTT cell viability assay for cytotoxicity testing in multidrug-resistant human leukemic cells. Leuk. Res. 1992, 16, 1165–1173. [Google Scholar] [CrossRef]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef]
- Berlin, A.; Castro-Mesta, J.F.; Rodriguez-Romo, L.; Hernandez-Barajas, D.; González-Guerrero, J.F.; Rodríguez-Fernández, I.A.; GonzálezConchas, G.; Verdines-Perez, A.; Vera-Badillo, F.E. Prognostic role of Ki-67 score in localized prostate cancer: A systematic review and meta-analysis. Urol. Oncol. 2017, 35, 499. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Montenarh, M.; Wagner, P. Regulation of CAK kinase activity by p53. Oncogene 1998, 17, 2733–2741. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Janicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Millauer, B.; Shawver, L.K.; Plate, K.H.; Risau, W.; Ullrich, A. Glioblastoma growth inhibited in vivo by a dominant negative Flk-1 mutant. Nature 1994, 367, 576–579. [Google Scholar] [CrossRef]
- Amin, B.H.; Amer, A.; Azzam, M.; Abd El-Sattar, N.E.A.; Mahmoud, D.; Al-Ashaal, S.; Al-Khalaf, B.A.; Hozzein, W.N. Antimicrobial and anticancer activities of Periplaneta americana tissue lysate: An in vitro study. J. King Saud. Univ. Sci. 2022, 34, 102095. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Protein | PDB Code | Binding Energy | RMSD | Amino Acids | Type of Bonding |
|---|---|---|---|---|---|---|
| BAPPN | Caspase-3 | 7JL7 | −7.836 | 1.483 | GLU195 | H-donor ionic |
| ARG 167 | Pi-cation |
| M.wt | Rotatable Bonds | HBA | HBD | TPSA | IlogP | GI Absorption | BBB Permeant | Lipinski Violations | Bioavailability Score |
|---|---|---|---|---|---|---|---|---|---|
| 430.5 | 8 | 4 | 2 | 62.3 | 3.67 | High | Yes | 0 | 0.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd Elrahman, S.F.; Ahmed, A.A.S.; Abd Elsatar, D.; Elkady, S.; Elgendy, A.; Alnakeeb, F.; Elmongy, E.I.; Henidi, H.A.; El-Gendy, S.M.; El Sayed, I.E.T.; et al. Cytotoxic Potential of Novel Quinoline Derivative: 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline against Different Cancer Cell Lines via Activation and Deactivation of the Expression of Some Proteins. Int. J. Mol. Sci. 2023, 24, 14336. https://doi.org/10.3390/ijms241814336
Abd Elrahman SF, Ahmed AAS, Abd Elsatar D, Elkady S, Elgendy A, Alnakeeb F, Elmongy EI, Henidi HA, El-Gendy SM, El Sayed IET, et al. Cytotoxic Potential of Novel Quinoline Derivative: 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline against Different Cancer Cell Lines via Activation and Deactivation of the Expression of Some Proteins. International Journal of Molecular Sciences. 2023; 24(18):14336. https://doi.org/10.3390/ijms241814336
Chicago/Turabian StyleAbd Elrahman, Sara Fathy, Abdullah A. S. Ahmed, Doaa Abd Elsatar, Salma Elkady, Amira Elgendy, Fatma Alnakeeb, Elshaymaa I. Elmongy, Hanan A. Henidi, Saad M. El-Gendy, Ibrahim El Tantawy El Sayed, and et al. 2023. "Cytotoxic Potential of Novel Quinoline Derivative: 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline against Different Cancer Cell Lines via Activation and Deactivation of the Expression of Some Proteins" International Journal of Molecular Sciences 24, no. 18: 14336. https://doi.org/10.3390/ijms241814336
APA StyleAbd Elrahman, S. F., Ahmed, A. A. S., Abd Elsatar, D., Elkady, S., Elgendy, A., Alnakeeb, F., Elmongy, E. I., Henidi, H. A., El-Gendy, S. M., El Sayed, I. E. T., El-Gokha, A. A., & Abd Eldaim, M. A. (2023). Cytotoxic Potential of Novel Quinoline Derivative: 11-(1,4-Bisaminopropylpiperazinyl)5-methyl-5H-indolo[2,3-b]quinoline against Different Cancer Cell Lines via Activation and Deactivation of the Expression of Some Proteins. International Journal of Molecular Sciences, 24(18), 14336. https://doi.org/10.3390/ijms241814336