
Dynamic Interplay in Tumor Ecosystems: Communication between Hepatoma Cells and Fibroblasts

 , , , , ,
, , , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
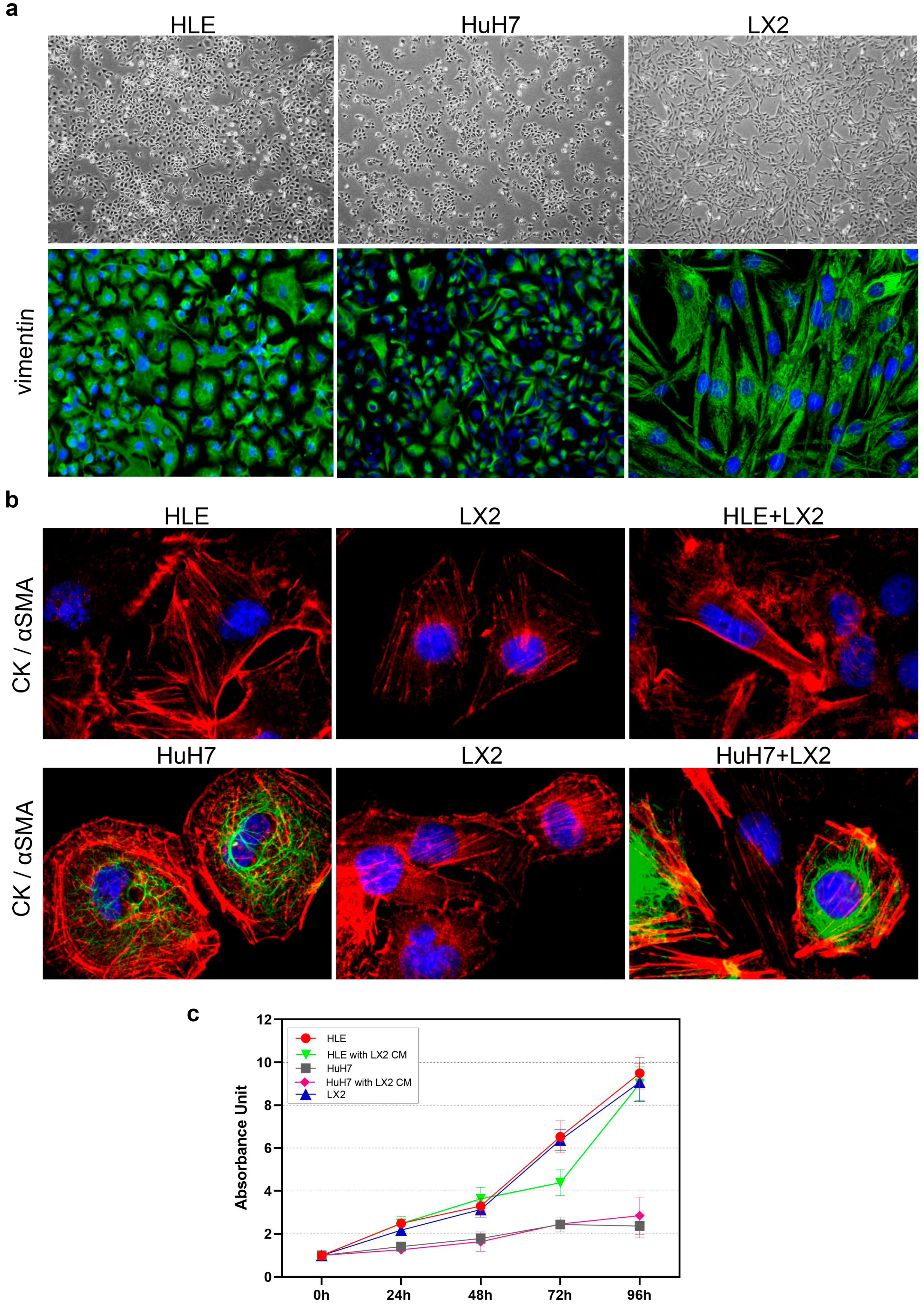
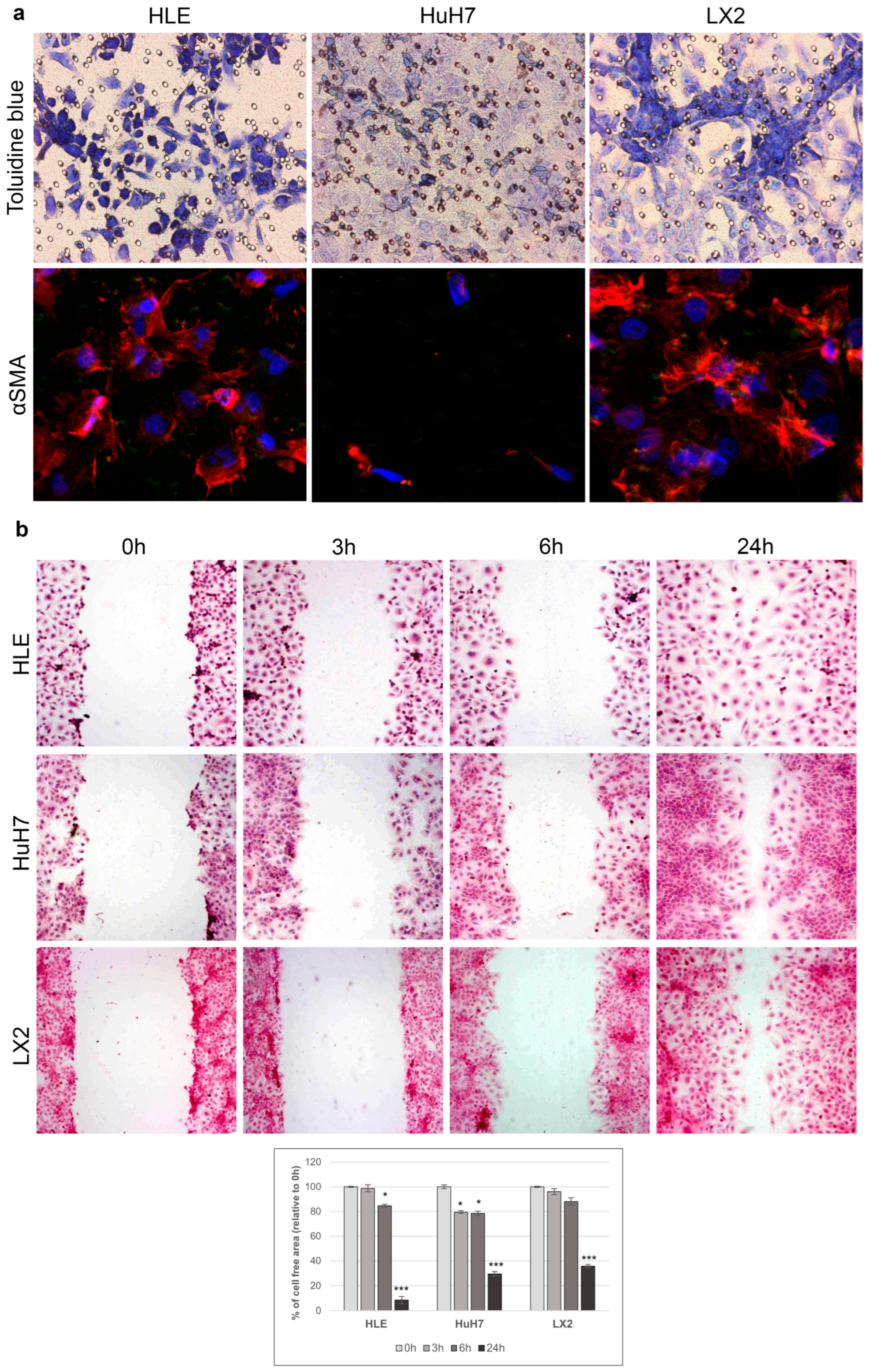
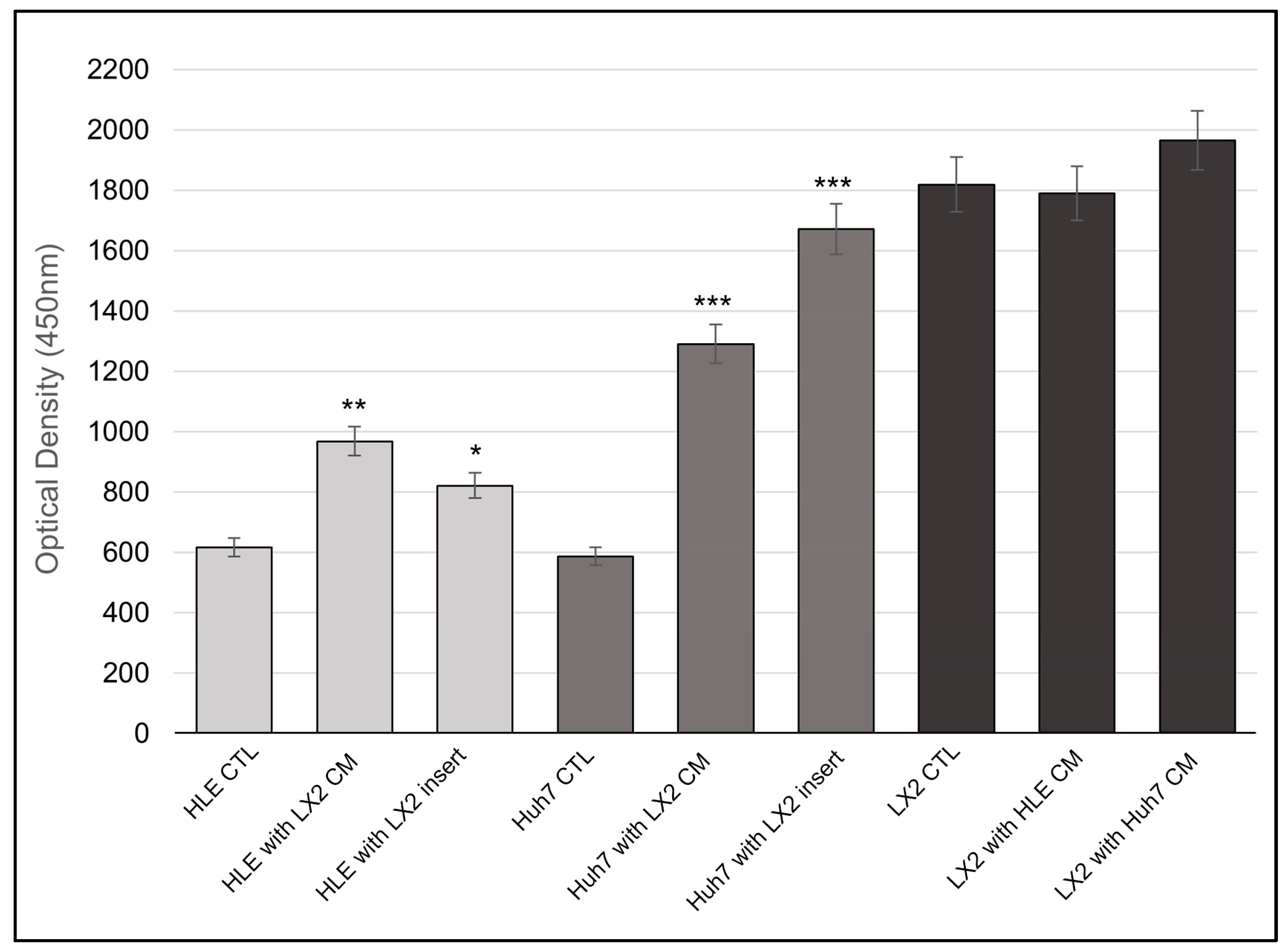
2.1. Morphology, Proliferation, and Invasion of HLE and HuH7 Hepatoma Cell Lines and LX2 Liver Fibroblast Cells
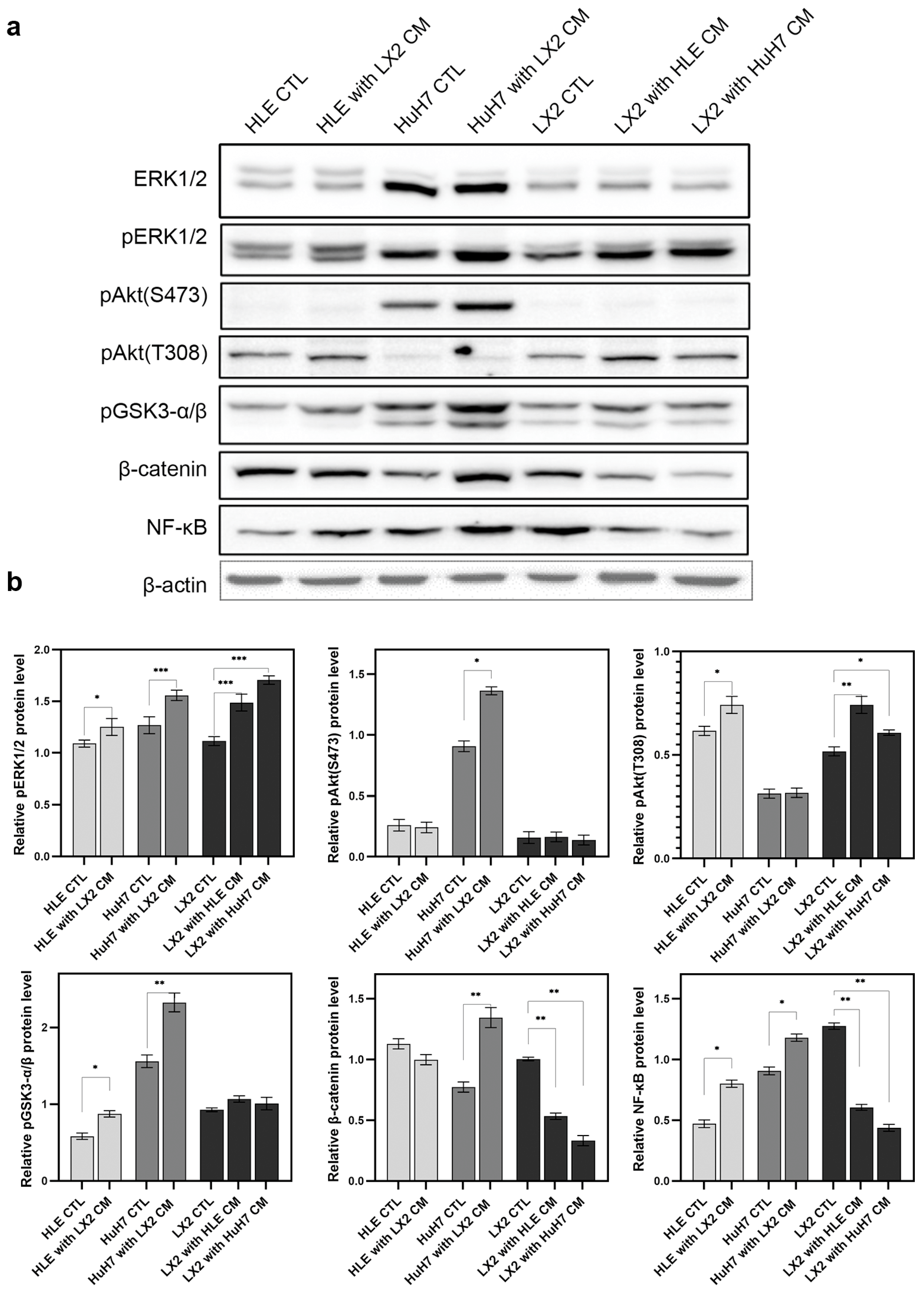
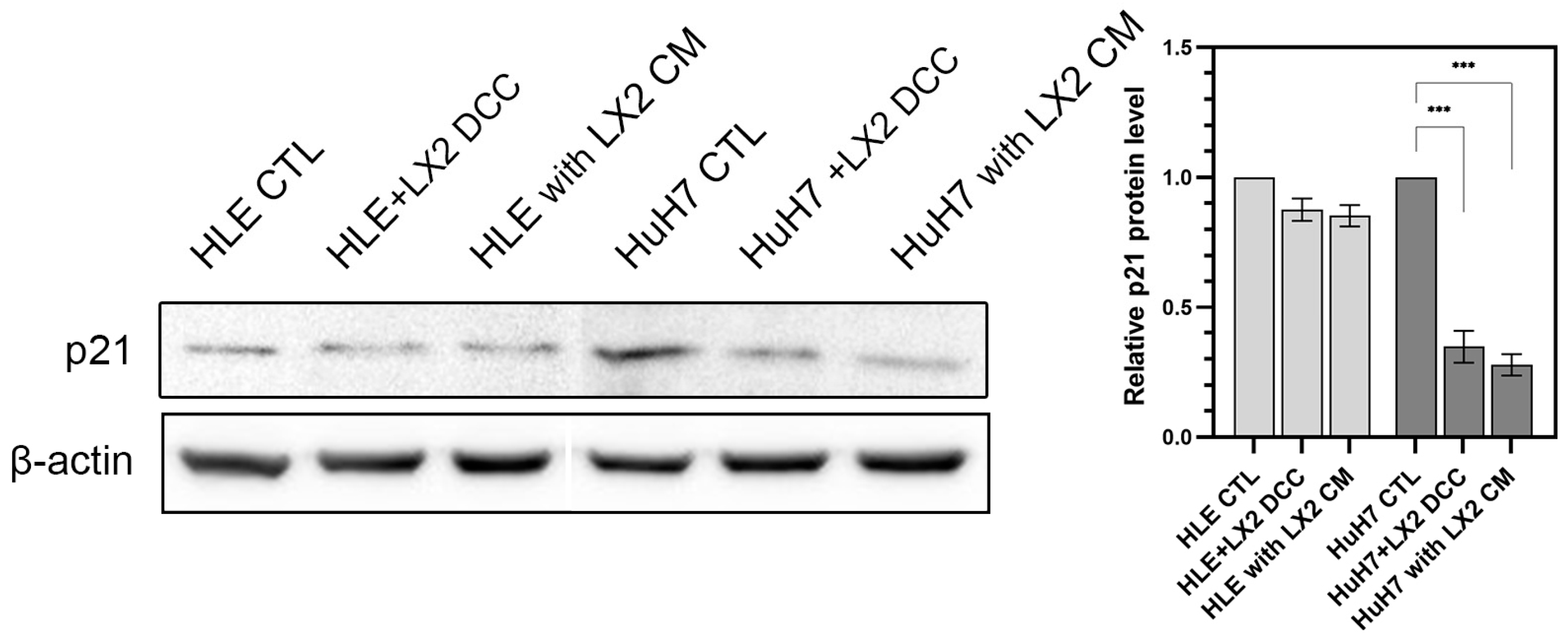
2.2. Crosstalk between Hepatoma Cells and LX2 via Soluble Mediators Mutually Modulates Signal Transduction and Downregulates Cyclin-Dependent Kinase Inhibitor p21
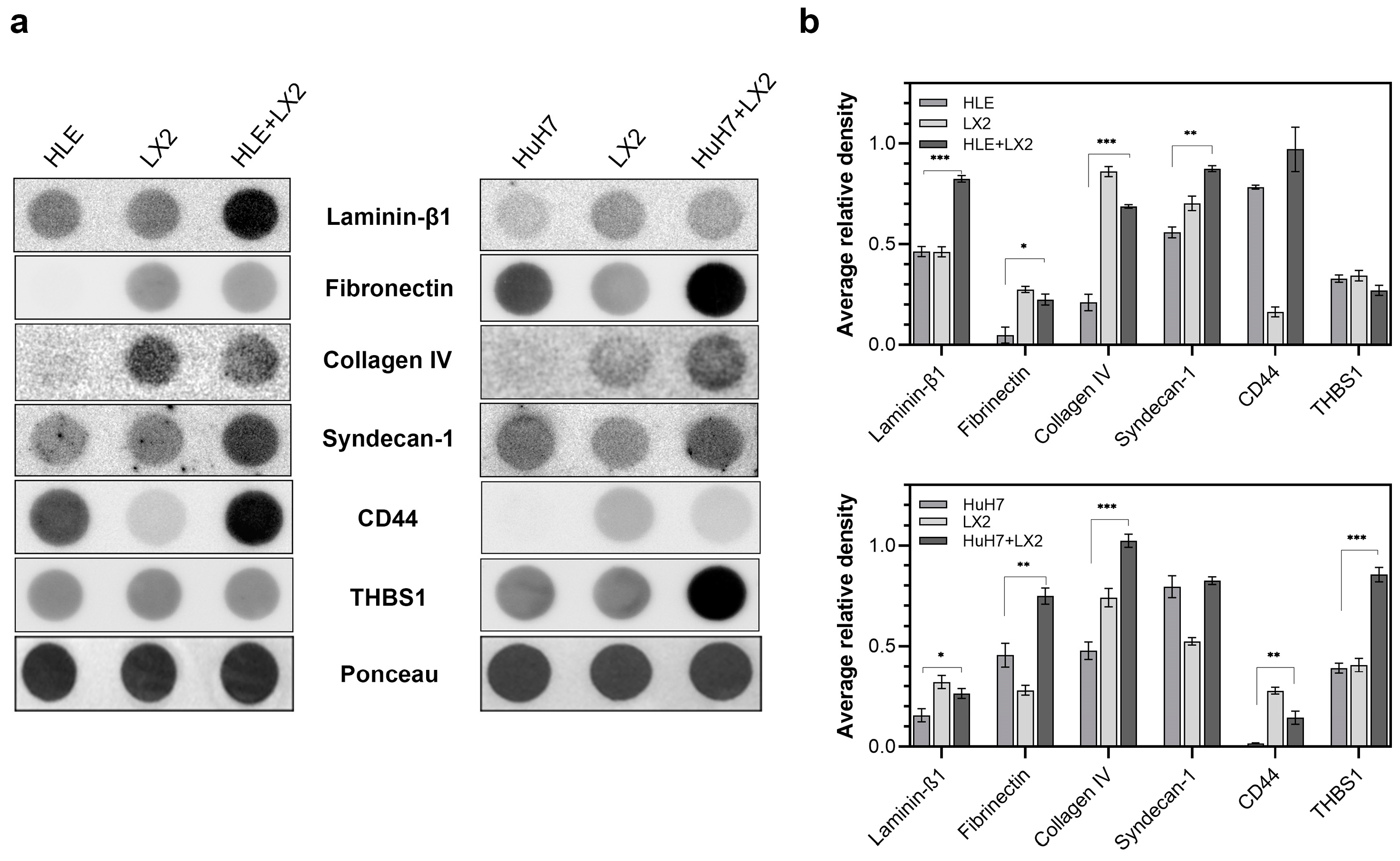
2.3. Contact Co-Culture of Hepatomas and Fibroblasts Alters the Expression of ECM Proteins
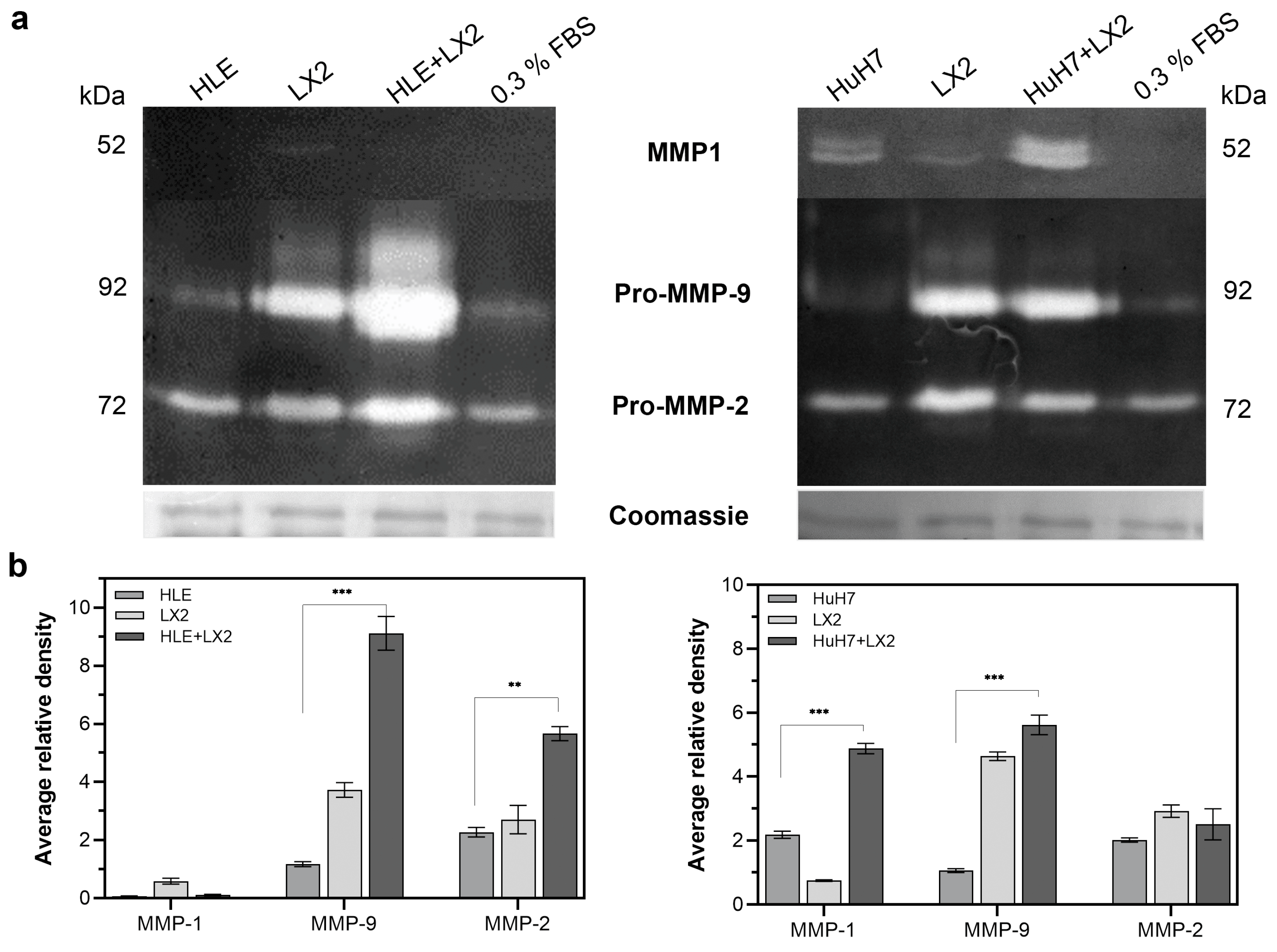
2.4. Expression of Matrix Metalloproteases (MMPs) Is Enhanced by Co-Culturing Hepatoma Cells with LX2 Fibroblasts
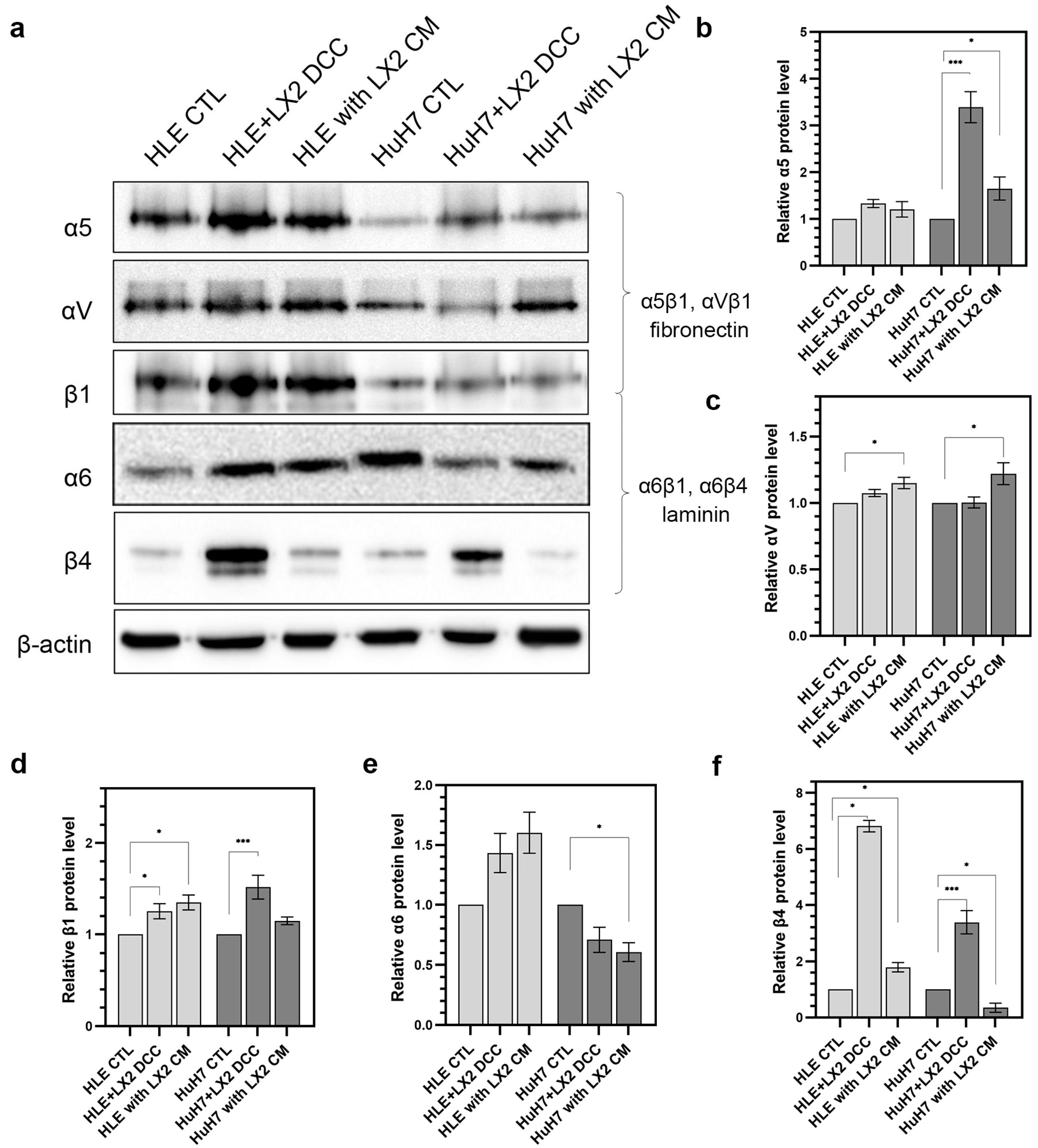
2.5. The Expression of Integrins and Their Cooperation with Adhesive Glycoproteins Are Hepatoma Cell Type-Dependent
2.6. Conditioned Medium of LX2 Cells Upregulates CXCL12 in the Hepatoma Cell Lines
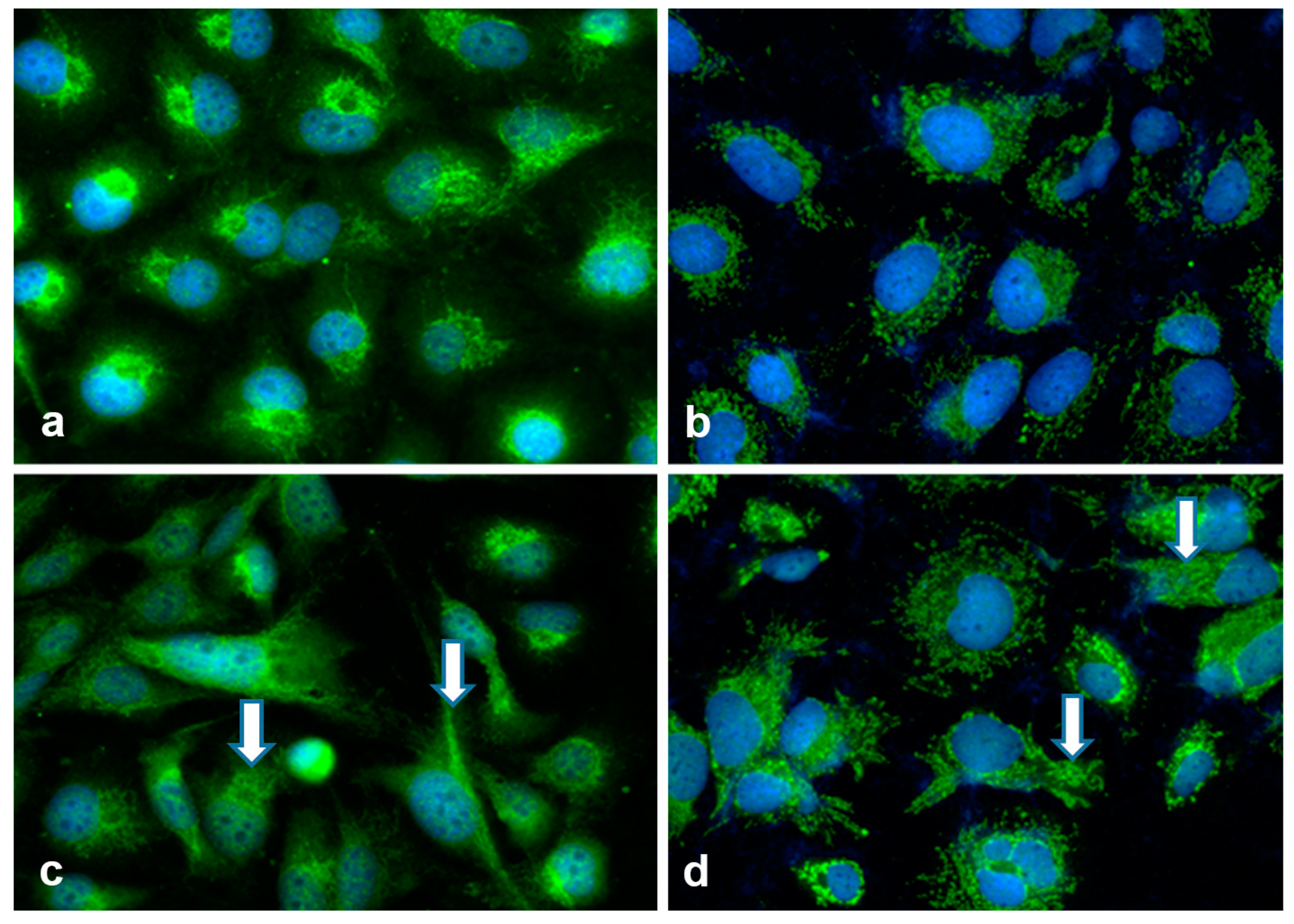
2.7. Hepatoma Cell Lines and Fibroblasts Communicate by EVs
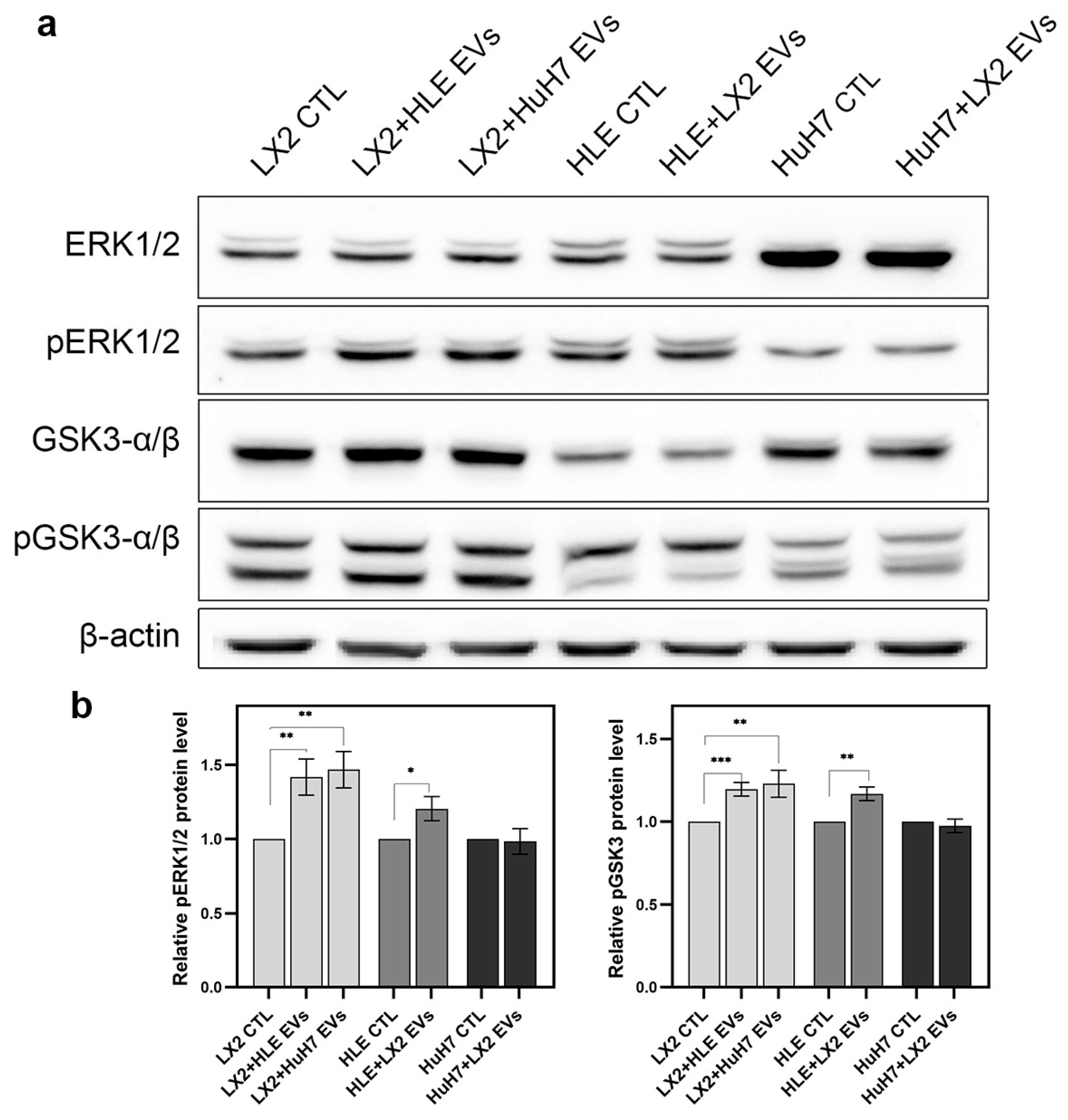
2.7.1. Hepatoma EVs Activate ERK1/2 and Inhibit GSK3 Function in LX2 Cells
2.7.2. LX2 EVs Alter the Cargo Composition of Hepatoma EVs
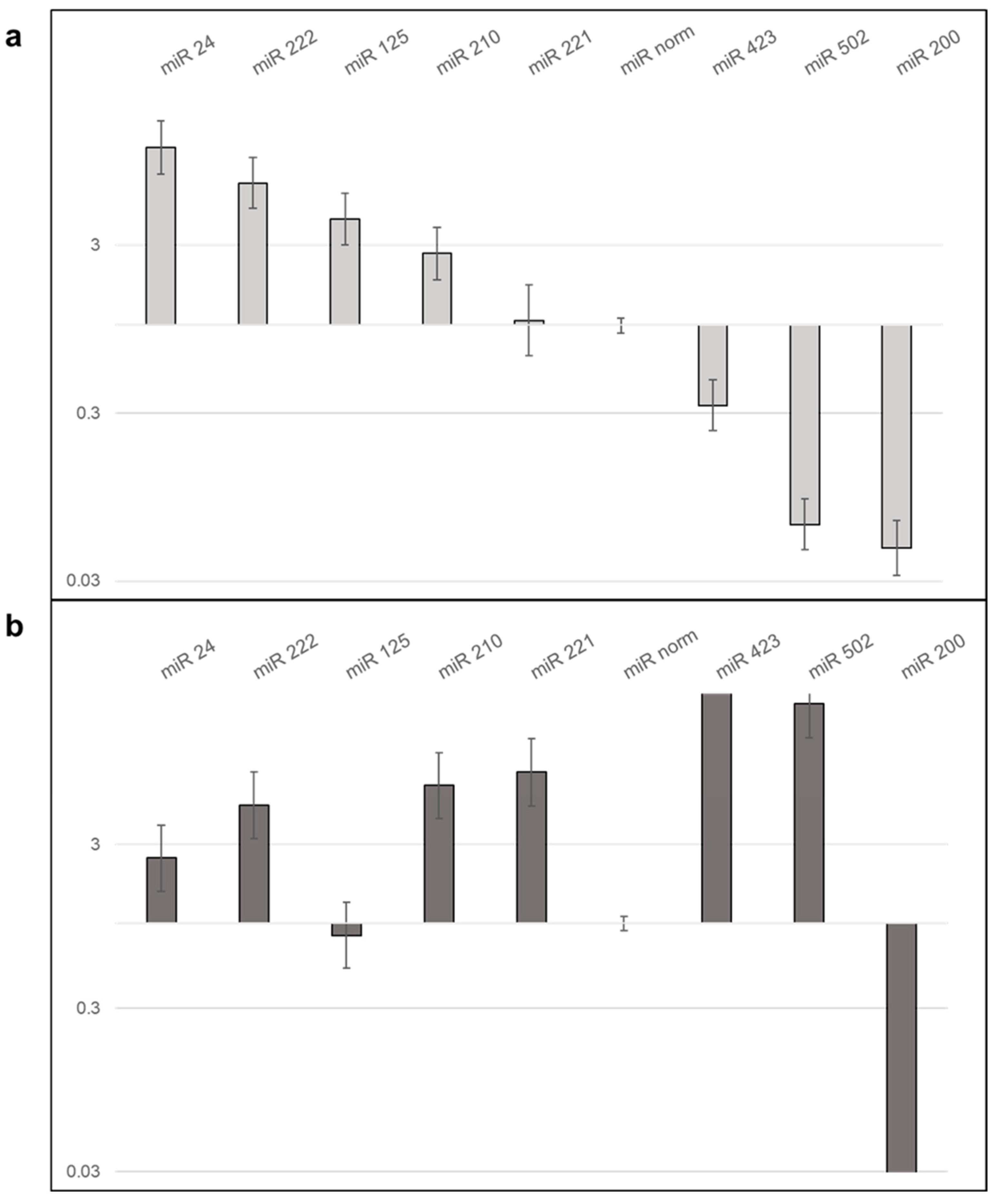
2.7.3. LX2 EVs Modify the Expression of miRNA of Hepatoma Cell Lines
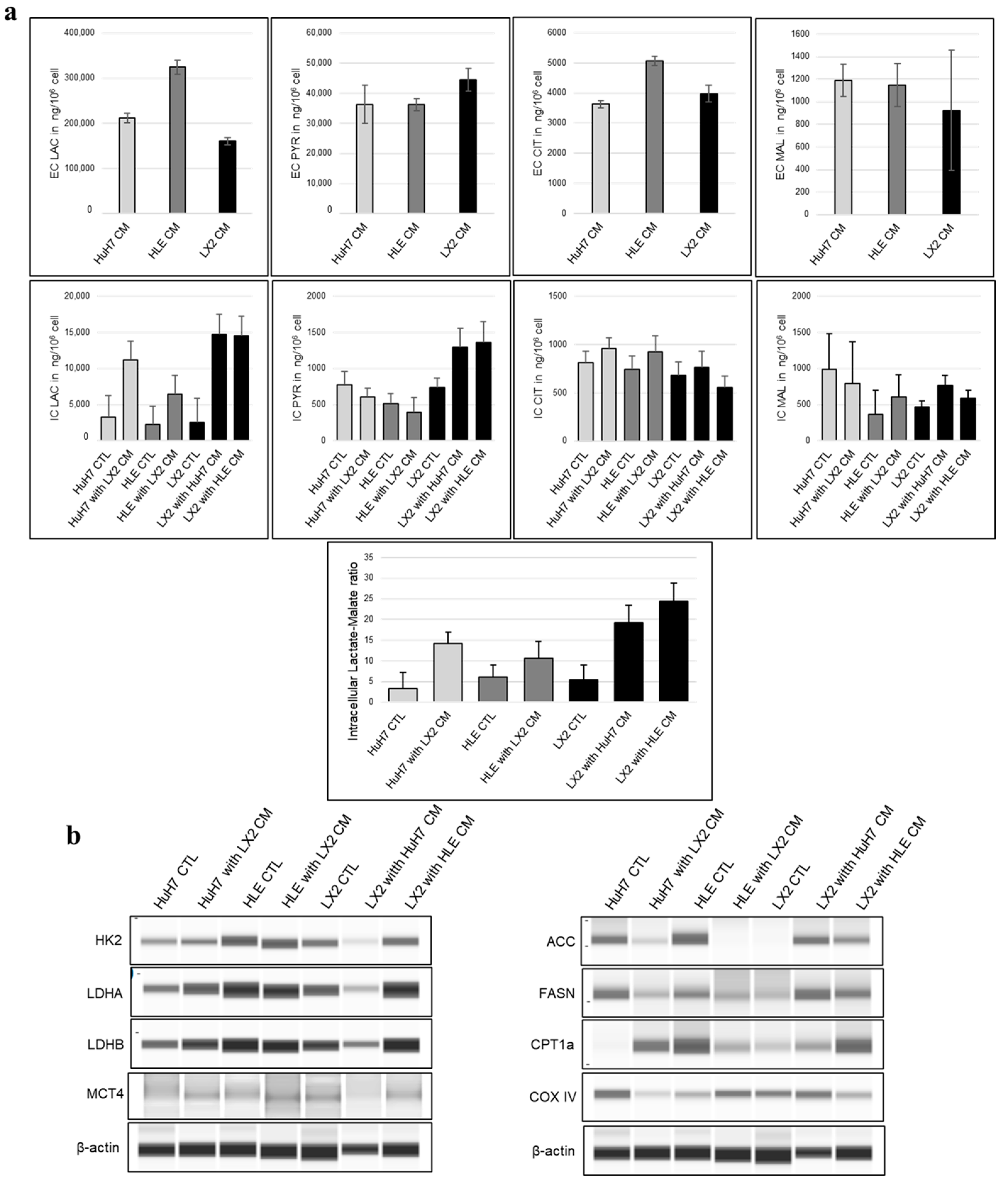
2.8. Conditioned Media of Tumor Cells and Fibroblasts Mutually Alter the Intermediary Metabolism
3. Discussion
3.1. HLE and HuH7 as Co-Culture Partners Modeling Poorly Differentiated vs. Differentiated Hepatoma
3.2. Conditioned Medium of LX2 Cells Upregulated CXCL12 Expression in Hepatoma Cell Lines
3.3. EVs as New Messengers between Hepatomas and Fibroblast Communication
3.4. EVs Isolated from the Hepatoma Cell Lines Contain Regulatory miRNAs
3.5. Modulation of Metabolism by HCC–Fibroblast Interactions
3.6. Cancer–Stromal Interactions in Hepatocellular Carcinoma: Insights, Limitations, and Future Prospects
4. Materials and Methods
4.1. Cell Cultures
4.2. Co-Culture Systems
4.3. Cell Proliferation Assay
4.4. Chemotaxis Assay
4.5. Wound Healing Assay
4.6. Immunofluorescence
4.7. Expression Analysis of Proteins by Western Blot, Dot Blot, and WES Simple Capillary Immunoassay
4.8. Zymography Assay
4.9. ELISA
4.10. Extracellular Vesicle (EV) Isolation, Total EV RNA Isolation, and miRNA Expression Profiling
4.11. Chromatography and Mass Spectrometry for Proteomics Analyses
4.12. Metabolite Analysis by Liquid Chromatography Mass Spectrometry
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sebestyén, A.; Dankó, T.; Sztankovics, D.; Moldvai, D.; Raffay, R.; Cervi, C.; Krencz, I.; Zsiros, V.; Jeney, A.; Petővári, G. The role of metabolic ecosystem in cancer progression—Metabolic plasticity and mTOR hyperactivity in tumor tissues. Cancer Metastasis Rev. 2021, 40, 989–1033. [Google Scholar] [CrossRef]
- Martinez-Outschoorn, U.E.; Bartrons, M.; Bartrons, R. Cancer Ecosystems. Front. Oncol. 2019, 9, 718. [Google Scholar] [CrossRef] [PubMed]
- De Wever, O.; Mareel, M. Role of tissue stroma in cancer cell invasion. J. Pathol. 2003, 200, 429–447. [Google Scholar] [CrossRef]
- Karta, J.; Bossicard, Y.; Kotzamanis, K.; Dolznig, H.; Letellier, E. Mapping the Metabolic Networks of Tumor Cells and Cancer-Associated Fibroblasts. Cells 2021, 10, 304. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, J.; Ogawa, M. Cell biologic factors and cancer spread. Int. J. Oncol. 1997, 10, 807–813. [Google Scholar] [CrossRef]
- Dvorak, H.F. Tumors: Wounds that do not heal. Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Louise Jones, J. Jekyll and Hyde: The role of the microenvironment on the progression of cancer. J. Pathol. 2011, 223, 162–176. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Sahai, E.; Rullan, A. SnapShot: Cancer-Associated Fibroblasts. Cell 2020, 181, 486–486.e481. [Google Scholar] [CrossRef]
- Arcucci, A.; Ruocco, M.R.; Granato, G.; Sacco, A.M.; Montagnani, S. Cancer: An Oxidative Crosstalk between Solid Tumor Cells and Cancer Associated Fibroblasts. BioMed Res. Int. 2016, 2016, 4502846. [Google Scholar] [CrossRef]
- Zeltz, C.; Primac, I.; Erusappan, P.; Alam, J.; Noel, A.; Gullberg, D. Cancer-associated fibroblasts in desmoplastic tumors: Emerging role of integrins. Semin. Cancer Biol. 2020, 62, 166–181. [Google Scholar] [CrossRef]
- Zeltz, C.; Lu, N.; Heljasvaara, R.; Gullberg, D. Integrins in Cancer: Refocusing on the Tumor Microenvironment. In The Extracellular Matrix and the Tumor Microenvironment; Kovalszky, I., Franchi, M., Alaniz, L.D., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 279–314. [Google Scholar]
- Lin, N.; Meng, L.; Lin, J.; Chen, S.; Zhang, P.; Chen, Q.; Lin, Y. Activated hepatic stellate cells promote angiogenesis in hepatocellular carcinoma by secreting angiopoietin-1. J. Cell. Biochem. 2020, 121, 1441–1451. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Jaillon, S.; Garlanda, C.; Allavena, P. Tumor-associated myeloid cells: Diversity and therapeutic targeting. Cell. Mol. Immunol. 2021, 18, 566–578. [Google Scholar] [CrossRef]
- Akkaya, M.; Kwak, K.; Pierce, S.K. B cell memory: Building two walls of protection against pathogens. Nat. Rev. Immunol. 2020, 20, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Oncology Meets Immunology: The Cancer-Immunity Cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Tan, Z.W.; Zhu, P.; Tan, N.S. Cancer-associated fibroblasts in tumor microenvironment—Accomplices in tumor malignancy. Cell. Immunol. 2019, 343, 103729. [Google Scholar] [CrossRef] [PubMed]
- Goenka, A.; Khan, F.; Verma, B.; Sinha, P.; Dmello, C.C.; Jogalekar, M.P.; Gangadaran, P.; Ahn, B.C. Tumor microenvironment signaling and therapeutics in cancer progression. Cancer Commun. 2023, 43, 525–561. [Google Scholar] [CrossRef]
- Rimal, R.; Desai, P.; Daware, R.; Hosseinnejad, A.; Prakash, J.; Lammers, T.; Singh, S. Cancer-associated fibroblasts: Origin, function, imaging, and therapeutic targeting. Adv. Drug Deliv. Rev. 2022, 189, 114504. [Google Scholar] [CrossRef]
- Yeon, J.H.; Jeong, H.E.; Seo, H.; Cho, S.; Kim, K.; Na, D.; Chung, S.; Park, J.; Choi, N.; Kang, J.Y. Cancer-derived exosomes trigger endothelial to mesenchymal transition followed by the induction of cancer-associated fibroblasts. Acta Biomater. 2018, 76, 146–153. [Google Scholar] [CrossRef]
- Delinassios, J.G.; Hoffman, R.M. The cancer-inhibitory effects of proliferating tumor-residing fibroblasts. Biochim. Biophys. Acta (BBA) Rev. Cancer 2022, 1877, 188673. [Google Scholar] [CrossRef]
- Biffi, G.; Tuveson, D.A. Diversity and Biology of Cancer-Associated Fibroblasts. Physiol. Rev. 2021, 101, 147–176. [Google Scholar] [CrossRef]
- Mason, J.; Öhlund, D. Key aspects for conception and construction of co-culture models of tumor-stroma interactions. Front. Bioeng. Biotechnol. 2023, 11, 1150764. [Google Scholar] [CrossRef]
- Fullár, A.; Dudás, J.; Oláh, L.; Hollósi, P.; Papp, Z.; Sobel, G.; Karászi, K.; Paku, S.; Baghy, K.; Kovalszky, I. Remodeling of extracellular matrix by normal and tumor-associated fibroblasts promotes cervical cancer progression. BMC Cancer 2015, 15, 256. [Google Scholar] [CrossRef] [PubMed]
- Teicher, B.A.; Fricker, S.P. CXCL12 (SDF-1)/CXCR4 Pathway in Cancer. Clin. Cancer Res. 2010, 16, 2927–2931. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, I.; Riveiro, M.E.; Paradis, V.; Faivre, S.; De Parga, P.M.V.; Raymond, E. Insights on the CXCL12-CXCR4 axis in hepatocellular carcinoma carcinogenesis. Am. J. Transl. Res. 2014, 6, 340–352. [Google Scholar] [PubMed]
- Liepelt, A.; Tacke, F. Stromal cell-derived factor-1 (SDF-1) as a target in liver diseases. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G203–G209. [Google Scholar] [CrossRef]
- Zhou, W.; Guo, S.; Liu, M.; Burow, M.E.; Wang, G. Targeting CXCL12/CXCR4 Axis in Tumor Immunotherapy. Curr. Med. Chem. 2019, 26, 3026–3041. [Google Scholar] [CrossRef]
- Mitchell, M.I.; Ma, J.; Carter, C.L.; Loudig, O. Circulating Exosome Cargoes Contain Functionally Diverse Cancer Biomarkers: From Biogenesis and Function to Purification and Potential Translational Utility. Cancers 2022, 14, 3350. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Huda, M.N.; Nafiujjaman, M.; Deaguero, I.G.; Okonkwo, J.; Hill, M.L.; Kim, T.; Nurunnabi, M. Potential Use of Exosomes as Diagnostic Biomarkers and in Targeted Drug Delivery: Progress in Clinical and Preclinical Applications. ACS Biomater. Sci. Eng. 2021, 7, 2106–2149. [Google Scholar] [CrossRef]
- Han, J.; Rong, Y.; Gao, X. Multiomic analysis of the function of SPOCK1 across cancers: An integrated bioinformatics approach. J. Int. Med. Res. 2021, 49, 300060520962659. [Google Scholar] [CrossRef]
- Marimuthu, A.; Jacob, H.K.; Jakharia, A.; Subbannayya, Y.; Keerthikumar, S.; Kashyap, M.K.; Goel, R.; Balakrishnan, L.; Dwivedi, S.; Pathare, S.; et al. Gene Expression Profiling of Gastric Cancer. J. Proteom. Bioinform. 2011, 4, 74–82. [Google Scholar] [CrossRef]
- Zhao, P.; Guan, H.-T.; Dai, Z.-J.; Ma, Y.-G.; Liu, X.-X.; Wang, X.-J. Knockdown of SPOCK1 Inhibits the Proliferation and Invasion in Colorectal Cancer Cells by Suppressing the PI3K/Akt Pathway. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2016, 24, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Váncza, L.; Karászi, K.; Péterfia, B.; Turiák, L.; Dezső, K.; Sebestyén, A.; Reszegi, A.; Petővári, G.; Kiss, A.; Schaff, Z.; et al. SPOCK1 Promotes the Development of Hepatocellular Carcinoma. Front. Oncol. 2022, 12, 819883. [Google Scholar] [CrossRef] [PubMed]
- Váncza, L.; Horváth, A.; Seungyeon, L.; Rókusz, A.; Dezső, K.; Reszegi, A.; Petővári, G.; Götte, M.; Kovalszky, I.; Baghy, K. SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer. Cancers 2023, 15, 2037. [Google Scholar] [CrossRef]
- Jiang, S.; Li, Q.; Wang, C.; Pang, Y.; Sun, Z.; Xiao, R. In Situ Exosomal MicroRNA Determination by Target-Triggered SERS and Fe3O4@TiO2-Based Exosome Accumulation. ACS Sensors 2021, 6, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Yang, Y.; Kannisto, E.; Zeng, X.; Yu, G.; Patnaik, S.K.; Dy, G.K.; Reid, M.E.; Gan, Q.; Wu, Y. Simultaneous Detection of Tumor Derived Exosomal Protein–MicroRNA Pairs with an Exo-PROS Biosensor for Cancer Diagnosis. ACS Nano 2023, 17, 8108–8122. [Google Scholar] [CrossRef] [PubMed]
- Joshi, G.K.; Deitz-McElyea, S.; Liyanage, T.; Lawrence, K.; Mali, S.; Sardar, R.; Korc, M. Label-Free Nanoplasmonic-Based Short Noncoding RNA Sensing at Attomolar Concentrations Allows for Quantitative and Highly Specific Assay of MicroRNA-10b in Biological Fluids and Circulating Exosomes. ACS Nano 2015, 9, 11075–11089. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Luo, L.; Chen, W.; Xu, H.-X.; Chen, F.; Chen, L.-Z.; Zeng, W.-T.; Chen, J.-S.; Huang, X.-H. MicroRNA-24 increases hepatocellular carcinoma cell metastasis and invasion by targeting p53: MiR-24 targeted p53. Biomed. Pharmacother. 2016, 84, 1113–1118. [Google Scholar] [CrossRef]
- Ma, Y.; She, X.-G.; Ming, Y.-Z.; Wan, Q.-Q. miR-24 promotes the proliferation and invasion of HCC cells by targeting SOX7. Tumor Biol. 2014, 35, 10731–10736. [Google Scholar] [CrossRef]
- Oura, K.; Fujita, K.; Morishita, A.; Iwama, H.; Nakahara, M.; Tadokoro, T.; Sakamoto, T.; Nomura, T.; Yoneyama, H.; Mimura, S.; et al. Serum microRNA-125a-5p as a potential biomarker of HCV-associated hepatocellular carcinoma. Oncol. Lett. 2019, 18, 882–890. [Google Scholar] [CrossRef]
- Coppola, N.; de Stefano, G.; Panella, M.; Onorato, L.; Iodice, V.; Minichini, C.; Mosca, N.; Desiato, L.; Farella, N.; Starace, M.; et al. Lowered expression of microRNA-125a-5p in human hepatocellular carcinoma and up-regulation of its oncogenic targets sirtuin-7, matrix metalloproteinase-11, and c-Raf. Oncotarget 2017, 8, 25289–25299. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Yu, W.; Chen, J.; Luo, J. Expression of serum miR-221 in human hepatocellular carcinoma and its prognostic significance. Biochem. Biophys. Res. Commun. 2011, 406, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jiang, J.; Badawi, M.; Schmittgen, T.D. miR-221 regulates CD44 in hepatocellular carcinoma through the PI3K-AKT-mTOR pathway. Biochem. Biophys. Res. Commun. 2017, 487, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; An, P.; Winkler, C.A.; Yu, Y. Dysregulated microRNAs in Hepatitis B Virus-Related Hepatocellular Carcinoma: Potential as Biomarkers and Therapeutic Targets. Front. Oncol. 2020, 10, 1271. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cui, Y. Bim’s Effect on the Expression of miR-423-3p in Promoting Primary Hepatic Cancer (PHC) and Role of miR-423-3p in PHC Proliferation and Invasion. Biochem. Genet. 2021, 59, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.-S.; Liu, H.-H.; Liu, J.-J.; Yeh, C.-T.; Chang, T.-C.; Wu, C.-H.; Ho, Y.-S.; Wei, P.-L.; Chang, Y.-J. MicroRNA-200a and -200b Mediated Hepatocellular Carcinoma Cell Migration through the Epithelial to Mesenchymal Transition Markers. Ann. Surg. Oncol. 2013, 20 (Suppl. 3), S360–S368. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.-S.; Wang, F.M.; Chen, T.-C.; Yeh, C.-N.; Yu, M.-C.; Jan, Y.-Y.; Chen, M.-F. Expression Profile of MicroRNA-200 Family in Hepatocellular Carcinoma with Bile Duct Tumor Thrombus. Ann. Surg. 2014, 259, 346–354. [Google Scholar] [CrossRef]
- Fang, M.; Yuan, J.; Chen, M.; Sun, Z.; Liu, L.; Cheng, G.; Ying, H.; Yang, S.; Chen, M. The heterogenic tumor microenvironment of hepatocellular carcinoma and prognostic analysis based on tumor neo-vessels, macrophages and α-SMA. Oncol. Lett. 2018, 15, 4805–4812. [Google Scholar] [CrossRef]
- Lau, E.Y.T.; Lo, J.; Cheng, B.Y.L.; Ma, M.K.F.; Lee, J.M.F.; Ng, J.K.Y.; Chai, S.; Lin, C.H.; Tsang, S.Y.; Ma, S.; et al. Cancer-Associated Fibroblasts Regulate Tumor-Initiating Cell Plasticity in Hepatocellular Carcinoma through c-Met/FRA1/HEY1 Signaling. Cell Rep. 2016, 15, 1175–1189. [Google Scholar] [CrossRef]
- Ding, S.; Chen, G.; Zhang, W.; Xing, C.; Xu, X.; Xie, H.; Lu, A.; Chen, K.; Guo, H.; Ren, Z.; et al. MRC-5 fibroblast-conditioned medium influences multiple pathways regulating invasion, migration, proliferation, and apoptosis in hepatocellular carcinoma. J. Transl. Med. 2015, 13, 237. [Google Scholar] [CrossRef] [PubMed]
- Skorupa, A.; Ciszek, M.; Turska-D’amico, M.; Stobiecka, E.; Chmielik, E.; Szumniak, R.; D’amico, A.; Boguszewicz, L.; Sokół, M. The Relationship between Histological Composition and Metabolic Profile in Breast Tumors and Peritumoral Tissue Determined with 1H HR-MAS NMR Spectroscopy. Cancers 2023, 15, 1283. [Google Scholar] [CrossRef]
- Jordan, K.; Stanton, E.H.; Milenkovic, V.M.; Federlin, M.; Drexler, K.; Buchalla, W.; Gaumann, A.; Adamski, J.; Proescholdt, M.; Haferkamp, S.; et al. Potential Involvement of Extracellular Citrate in Brain Tumor Progression. Curr. Mol. Med. 2022, 22, 506–513. [Google Scholar] [CrossRef]
- Andersen, M.K.; Høiem, T.S.; Claes, B.S.R.; Balluff, B.; Martin-Lorenzo, M.; Richardsen, E.; Krossa, S.; Bertilsson, H.; Heeren, R.M.A.; Rye, M.B.; et al. Spatial differentiation of metabolism in prostate cancer tissue by MALDI-TOF MSI. Cancer Metab. 2021, 9, 9. [Google Scholar] [CrossRef]
- Lu, L.; Huang, J.; Mo, J.; Da, X.; Li, Q.; Fan, M.; Lu, H. Exosomal lncRNA TUG1 from cancer-associated fibroblasts promotes liver cancer cell migration, invasion, and glycolysis by regulating the miR-524-5p/SIX1 axis. Cell. Mol. Biol. Lett. 2022, 27, 17. [Google Scholar] [CrossRef] [PubMed]
- Rattigan, Y.I.; Patel, B.B.; Ackerstaff, E.; Sukenick, G.; Koutcher, J.A.; Glod, J.W.; Banerjee, D. Lactate is a mediator of metabolic cooperation between stromal carcinoma associated fibroblasts and glycolytic tumor cells in the tumor microenvironment. Exp. Cell Res. 2012, 318, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Pavlides, S.; Whitaker-Menezes, D.; Castello-Cros, R.; Flomenberg, N.; Witkiewicz, A.K.; Frank, P.G.; Casimiro, M.C.; Wang, C.; Fortina, P.; Addya, S.; et al. The reverse Warburg effect: Aerobic glycolysis in cancer associated fibroblasts and the tumor stroma. Cell Cycle 2009, 8, 3984–4001. [Google Scholar] [CrossRef]
- Fu, Y.; Zou, T.; Shen, X.; Nelson, P.J.; Li, J.; Wu, C.; Yang, J.; Zheng, Y.; Bruns, C.; Zhao, Y.; et al. Lipid metabolism in cancer progression and therapeutic strategies. MedComm 2021, 2, 27–59. [Google Scholar] [CrossRef]
- Maan, M.; Peters, J.M.; Dutta, M.; Patterson, A.D. Lipid metabolism and lipophagy in cancer. Biochem. Biophys. Res. Commun. 2018, 504, 582–589. [Google Scholar] [CrossRef]
- Wang, H.; Luo, J.; Chen, Z.; Cao, W.T.; Xu, H.F.; Gou, D.M.; Zhu, J.J. MicroRNA-24 can control triacylglycerol synthesis in goat mammary epithelial cells by targeting the fatty acid synthase gene. J. Dairy Sci. 2015, 98, 9001–9014. [Google Scholar] [CrossRef]
- Wang, J.-J.; Zhang, Y.-T.; Tseng, Y.J.; Zhang, J. miR-222 targets ACOX1, promotes triglyceride accumulation in hepatocytes. Hepatobiliary Pancreat. Dis. Int. 2019, 18, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Giatromanolaki, A.; Koukourakis, M.I.; Koutsopoulos, A.; Mendrinos, S.; Sivridis, E. The metabolic interactions between tumor cells and tumor-associated stroma (TAS) in prostatic cancer. Cancer Biol. Ther. 2012, 13, 1284–1289. [Google Scholar] [CrossRef]
- Witkiewicz, A.K.; Whitaker-Menezes, D.; Dasgupta, A.; Philp, N.J.; Lin, Z.; Gandara, R.; Sneddon, S.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Using the “reverse Warburg effect” to identify high-risk breast cancer patients: Stromal MCT4 predicts poor clinical outcome in triple-negative breast cancers. Cell Cycle 2012, 11, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Curry, J.M.; Tuluc, M.; Whitaker-Menezes, D.; Ames, J.A.; Anantharaman, A.; Butera, A.; Leiby, B.; Cognetti, D.M.; Sotgia, F.; Lisanti, M.P.; et al. Cancer metabolism, stemness and tumor recurrence: MCT1 and MCT4 are functional biomarkers of metabolic symbiosis in head and neck cancer. Cell Cycle 2013, 12, 1371–1384. [Google Scholar] [CrossRef]
- Icard, P.; Kafara, P.; Steyaert, J.-M.; Schwartz, L.; Lincet, H. The metabolic cooperation between cells in solid cancer tumors. Biochim. Biophys. Acta (BBA) Rev. Cancer 2014, 1846, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Outschoorn, U.E.; Lisanti, M.P.; Sotgia, F. Catabolic cancer-associated fibroblasts transfer energy and biomass to anabolic cancer cells, fueling tumor growth. Semin. Cancer Biol. 2014, 25, 47–60. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Szoboszlai, N.; Guo, X.; Ozohanics, O.; Oláh, J.; Gömöry, A.; Mihucz, V.G.; Jeney, A.; Vékey, K. Determination of energy metabolites in cancer cells by porous graphitic carbon liquid chromatography electrospray ionization mass spectrometry for the assessment of energy metabolism. Anal. Chim. Acta 2014, 819, 108–115. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petővári, G.; Tóth, G.; Turiák, L.; L. Kiss, A.; Pálóczi, K.; Sebestyén, A.; Pesti, A.; Kiss, A.; Baghy, K.; Dezső, K.; et al. Dynamic Interplay in Tumor Ecosystems: Communication between Hepatoma Cells and Fibroblasts. Int. J. Mol. Sci. 2023, 24, 13996. https://doi.org/10.3390/ijms241813996
Petővári G, Tóth G, Turiák L, L. Kiss A, Pálóczi K, Sebestyén A, Pesti A, Kiss A, Baghy K, Dezső K, et al. Dynamic Interplay in Tumor Ecosystems: Communication between Hepatoma Cells and Fibroblasts. International Journal of Molecular Sciences. 2023; 24(18):13996. https://doi.org/10.3390/ijms241813996
Chicago/Turabian StylePetővári, Gábor, Gábor Tóth, Lilla Turiák, Anna L. Kiss, Krisztina Pálóczi, Anna Sebestyén, Adrián Pesti, András Kiss, Kornélia Baghy, Katalin Dezső, and et al. 2023. "Dynamic Interplay in Tumor Ecosystems: Communication between Hepatoma Cells and Fibroblasts" International Journal of Molecular Sciences 24, no. 18: 13996. https://doi.org/10.3390/ijms241813996
APA StylePetővári, G., Tóth, G., Turiák, L., L. Kiss, A., Pálóczi, K., Sebestyén, A., Pesti, A., Kiss, A., Baghy, K., Dezső, K., Füle, T., Tátrai, P., Kovalszky, I., & Reszegi, A. (2023). Dynamic Interplay in Tumor Ecosystems: Communication between Hepatoma Cells and Fibroblasts. International Journal of Molecular Sciences, 24(18), 13996. https://doi.org/10.3390/ijms241813996