

Genotypes Distribution of Epstein–Barr Virus among Lymphoma Patients in Ethiopia
 , , ,
, , , 
Abstract
1. Introduction
2. Results
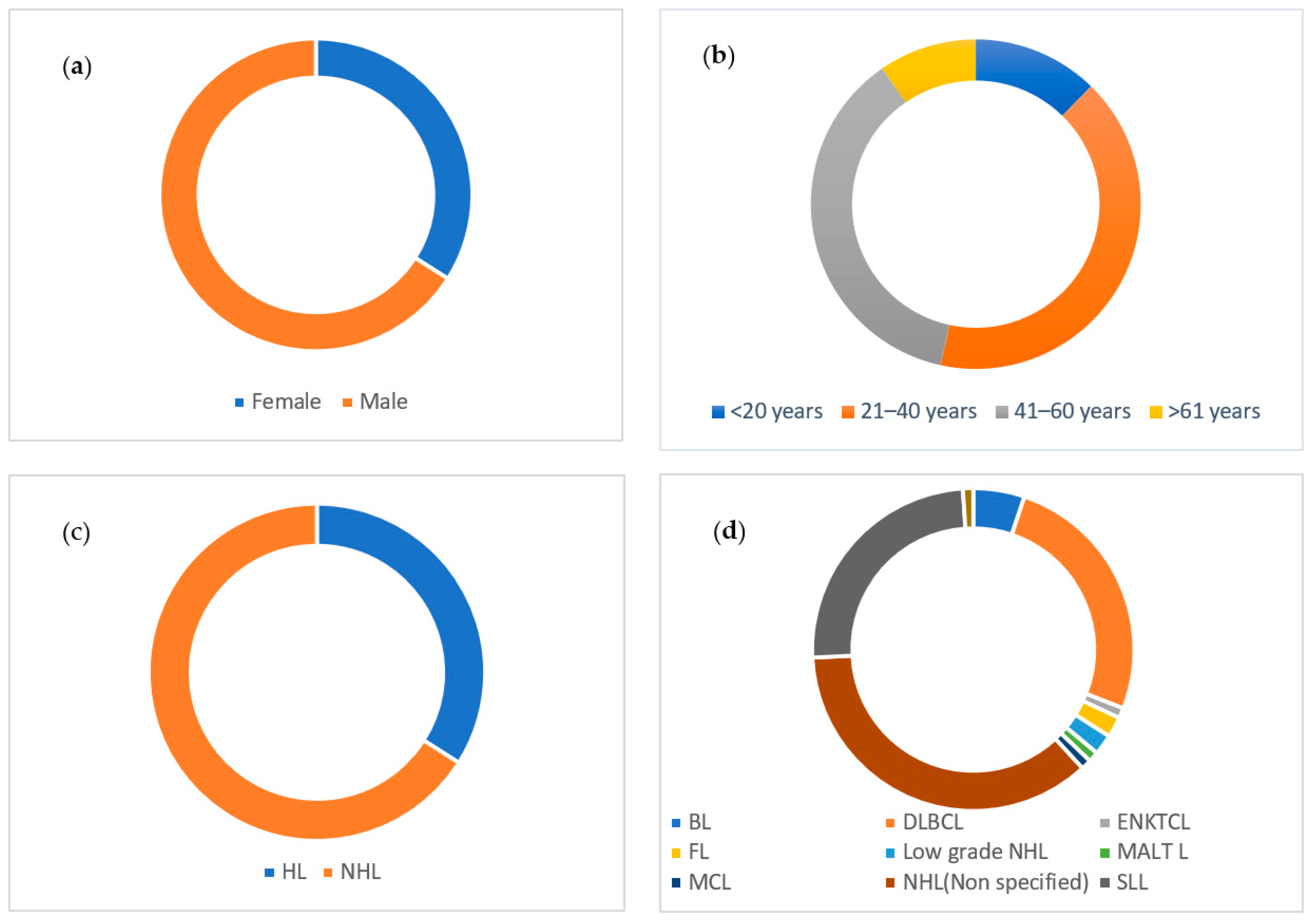
2.1. Sociodemographic and Clinical Characteristics
2.2. EBV Detection
2.3. EBNA3C Typing
3. Discussion
4. Materials and Methods
4.1. Study Design and Setting
4.2. Lymphoma Samples and Clinical Data
4.3. Extraction of Genomic DNA
4.4. EBV EBNA1 Gene Detection
4.5. EBV Typing Using EBNA3C Gene
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Epstein, M.; Achong, B.; Barr, Y. Virus particles in cultured lymphoblasts from burkitt’s lymphoma. Lancet 1964, 1, 702–703. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.P.; Kurzrock, R. Epstein-Barr virus and cancer. Clin. Cancer Res. 2004, 10, 803–821. [Google Scholar] [CrossRef] [PubMed]
- Santpere, G.; Darre, F.; Blanco, S.; Alcami, A.; Villoslada, P.; Albà, M.M.; Navarro, A. Genome-Wide Analysis of Wild-Type Epstein–Barr Virus Genomes Derived from Healthy Individuals of the 1000 Genomes Project. Genome Biol. Evol. 2014, 6, 846–860. [Google Scholar] [CrossRef] [PubMed]
- Maeda, E.; Akahane, M.; Kiryu, S.; Kato, N.; Yoshikawa, T.; Hayashi, N.; Aoki, S.; Minami, M.; Uozaki, H.; Fukayama, M.; et al. Spectrum of Epstein-Barr virus-related diseases: A pictorial review. Jpn. J. Radiol. 2009, 27, 4–19. [Google Scholar] [CrossRef]
- Speck, S.H.; Ganem, D. Viral Latency and Its Regulation: Lessons from the γ-Herpesviruses. Cell Host Microbe 2010, 8, 100–115. [Google Scholar] [CrossRef]
- Ansari, M.A.; Singh, V.V.; Dutta, S.; Veettil, M.V.; Dutta, D.; Chikoti, L.; Lu, J.; Everly, D.; Chandran, B. Constitutive interferon-inducible protein 16-inflammasome activation during Epstein-Barr virus latency I, II, and III in B and epithelial cells. J. Virol. 2013, 87, 8606–8623. [Google Scholar] [CrossRef]
- Münz, C. Epstein Barr Virus Volume 2: One Herpes Virus: Many Diseases; Springer: Berlin/Heidelberg, Germany, 2015; Volume 391. [Google Scholar]
- Wang, J.; Ge, J.; Wang, Y.; Xiong, F.; Guo, J.; Jiang, X.; Zhang, L.; Deng, X.; Gong, Z.; Zhang, S.; et al. EBV miRNAs BART11 and BART17-3p promote immune escape through the enhancer-mediated transcription of PD-L1. Nat. Commun. 2022, 13, 866. [Google Scholar] [CrossRef]
- Kenney, S.C. Reactivation and lytic replication of EBV. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- AArvey, A.; Tempera, I.; Tsai, K.; Chen, H.-S.; Tikhmyanova, N.; Klichinsky, M.; Leslie, C.; Lieberman, P.M. An Atlas of the Epstein-Barr Virus Transcriptome and Epigenome Reveals Host-Virus Regulatory Interactions. Cell Host Microbe 2012, 12, 233–245. [Google Scholar] [CrossRef]
- Ma, S.-D.; Hegde, S.; Young, K.H.; Sullivan, R.; Rajesh, D.; Zhou, Y.; Jankowska-Gan, E.; Burlingham, W.J.; Sun, X.; Gulley, M.L.; et al. A New Model of Epstein-Barr Virus Infection Reveals an Important Role for Early Lytic Viral Protein Expression in the Development of Lymphomas. J. Virol. 2011, 85, 165–177. [Google Scholar] [CrossRef]
- Chêne, A.; Donati, D.; Guerreiro-Cacais, A.O.; Levitsky, V.; Chen, Q.; I Falk, K.; Orem, J.; Kironde, F.; Wahlgren, M.; Bejarano, M.T. A Molecular Link between Malaria and Epstein–Barr Virus Reactivation. PLoS Pathog. 2007, 3, e80. [Google Scholar] [CrossRef]
- Wang, F.; Gregory, C.; Sample, C.; Rowe, M.; Liebowitz, D.; Murray, R.; Rickinson, A.; Kieff, E. Epstein-Barr virus latent membrane protein (LMP1) and nuclear proteins 2 and 3C are effectors of phenotypic changes in B lymphocytes: EBNA-2 and LMP1 cooperatively induce CD23. J. Virol. 1990, 64, 2309–2318. [Google Scholar] [CrossRef]
- Farrell, P.J. Epstein–Barr virus strain variation. In Epstein Barr Virus Volume 1: One Herpes Virus: Many Diseases; Springer: Berlin/Heidelberg, Germany, 2015; pp. 45–69. [Google Scholar]
- Dambaugh, T.; Hennessy, K.; Chamnankit, L.; Kieff, E. U2 region of Epstein-Barr virus DNA may encode Epstein-Barr nuclear antigen 2. Proc. Natl. Acad. Sci. USA 1984, 81, 7632–7636. [Google Scholar] [CrossRef] [PubMed]
- Sample, J.; Young, L.; Martin, B.; Chatman, T.; Kieff, E.; Rickinson, A.; Kieff, E. Epstein-Barr virus types 1 and 2 differ in their EBNA-3A, EBNA-3B, and EBNA-3C genes. J. Virol. 1990, 64, 4084–4092. [Google Scholar] [CrossRef] [PubMed]
- Tzellos, S. Mechanism of Superior B Cell Immortalisation Activity of Type 1 Epstein-Barr Virus. Ph.D. Thesis, Imperial College London, London, UK, 2014. [Google Scholar]
- Griffiths, P.D.; Zuckerman, A.; Banatvala, J. Principles and Practice of Clinical Virology; John Wiley & Sons: Hoboken, NJ, USA, 2000. [Google Scholar]
- Coleman, C.B.; Lang, J.; Sweet, L.A.; Smith, N.A.; Freed, B.M.; Pan, Z.; Haverkos, B.; Pelanda, R.; Rochford, R. Epstein-Barr Virus Type 2 Infects T Cells and Induces B Cell Lymphomagenesis in Humanized Mice. J. Virol. 2018, 92, e00813-18. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.B.; Daud, I.; O Ogolla, S.; A Ritchie, J.; A Smith, N.; O Sumba, P.; E Dent, A.; Rochford, R. Epstein-Barr Virus Type 2 Infects T Cells in Healthy Kenyan Children. J. Infect. Dis. 2017, 216, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.; Fitzmaurice, C.; Naghavi, M.; Ahmed, L.A. Global and regional incidence, mortality and disability-adjusted life-years for Epstein-Barr virus-attributable malignancies, 1990–2017. BMJ Open 2020, 10, e037505. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L. A review of human carcinogens—Part B: Biological agents. Lancet Oncol. 2009, 10, 321–322. [Google Scholar] [CrossRef]
- Dolan, A.; Addison, C.; Gatherer, D.; Davison, A.J.; McGeoch, D.J. The genome of Epstein–Barr virus type 2 strain AG876. Virology 2006, 350, 164–170. [Google Scholar] [CrossRef]
- Tiwawech, D.; Srivatanakul, P.; Karalak, A.; Ishida, T. Association between EBNA2 and LMP1 subtypes of Epstein-Barr virus and nasopharyngeal carcinoma in Thais. J. Clin. Virol. 2008, 42, 1–6. [Google Scholar] [CrossRef]
- Brooks, J.M.; Croom-Carter, D.S.G.; Leese, A.M.; Tierney, R.J.; Habeshaw, G.; Rickinson, A.B. Cytotoxic T-Lymphocyte Responses to a Polymorphic Epstein-Barr Virus Epitope Identify Healthy Carriers with Coresident Viral Strains. J. Virol. 2000, 74, 1801–1809. [Google Scholar] [CrossRef]
- Montes-Mojarro, I.A.; Chen, B.J.; Ramirez-Ibarguen, A.F.; Quezada-Fiallos, C.M.; Pérez-Báez, W.B.; Dueñas, D.; Casavilca-Zambrano, S.; Ortiz-Mayor, M.; Rojas-Bilbao, E.; García-Rivello, H.; et al. Mutational profile and EBV strains of extranodal NK/T-cell lymphoma, nasal type in Latin America. Mod. Pathol. 2020, 33, 781–791. [Google Scholar] [CrossRef]
- Wang, Y.J.; Ba, Y.; Chen, Q.Y.; Han, Y.Q. Clinical Significance of Peripheral Blood EBV-DNA Determination and Genotyping in Lymphoma Patients. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2021, 29, 1802–1806. [Google Scholar] [PubMed]
- Palma, I.; Sánchez, A.E.; Jiménez-Hernández, E.; Alvarez-Rodríguez, F.; Nava-Frias, M.; Valencia-Mayoral, P.; Salinas-Lara, C.; Velazquez-Guadarrama, N.; Portilla-Aguilar, J.; Pena, R.Y.; et al. Detection of Epstein-Barr Virus and Genotyping Based on EBNA2 Protein in Mexican Patients with Hodgkin Lymphoma: A Comparative Study in Children and Adults. Clin. Lymphoma Myeloma Leuk. 2013, 13, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Salahuddin, S.; Khan, J.; Azhar, J.; Whitehurs, C.B.; Qadri, I.; Shackelford, J.; Pagano, J.S.; Muhammad, D.; Richards, K.L. Prevalence of Epstein–Barr Virus Genotypes in Pakistani Lymphoma Patients. Asian Pac. J. Cancer Prev. 2018, 19, 3153–3159. [Google Scholar] [CrossRef] [PubMed]
- Begić, V.; Korać, P.; Gašparov, S.; Rozman, M.; Simicic, P.; Zidovec-Lepej, S. Molecular Characterisation of Epstein–Barr Virus in Classical Hodgkin Lymphoma. Int. J. Mol. Sci. 2022, 23, 15635. [Google Scholar] [CrossRef] [PubMed]
- Kafita, D.; Kaile, T.; Malyangu, E.; Tembo, R.; Zulu, E.; Chisanga, C.; Kalonda, A.; Samutela, M.; Polepole, P.; Kwenda, G. Evidence of EBV infection in lymphomas diagnosed in Lusaka, Zambia. Pan Afr. Med. J. 2018, 29, 1–11. [Google Scholar] [CrossRef]
- Tzellos, S.; Farrell, P.J. Epstein-Barr Virus Sequence Variation—Biology and Disease. Pathogens 2012, 1, 156–175. [Google Scholar] [CrossRef]
- Mechelli, R.; Manzari, C.; Policano, C.; Annese, A.; Picardi, E.; Umeton, R.; Fornasiero, A.; D’Erchia, A.M.; Buscarinu, M.C.; Agliardi, C.; et al. Epstein-Barr virus genetic variants are associated with multiple sclerosis. Neurology 2015, 84, 1362–1368. [Google Scholar] [CrossRef]
- Feng, F.-T.; Cui, Q.; Liu, W.-S.; Guo, Y.-M.; Feng, Q.-S.; Chen, L.-Z.; Xu, M.; Luo, B.; Li, D.-J.; Hu, L.-F.; et al. A single nucleotide polymorphism in the Epstein-Barr virus genome is strongly associated with a high risk of nasopharyngeal carcinoma. Chin. J. Cancer 2015, 34, 1–10. [Google Scholar] [CrossRef]
- Kwok, H.; Tong, A.H.; Lin, C.H.; Lok, S.; Farrell, P.J.; Kwong, D.L.; Chiang, A.K. Genomic sequencing and comparative analysis of Epstein-Barr virus genome isolated from primary nasopharyngeal carcinoma biopsy. PLoS ONE 2012, 7, e36939. [Google Scholar] [CrossRef]
- Li, H.P.; Chang, Y.S. Epstein-Barr virus latent membrane protein 1: Structure and functions. J. Biomed. Sci. 2003, 10, 490–504. [Google Scholar] [CrossRef] [PubMed]
- Mainou, B.A.; Raab-Traub, N. LMP1 Strain Variants: Biological and Molecular Properties. J. Virol. 2006, 80, 6458–6468. [Google Scholar] [CrossRef] [PubMed]
- Görzer, I.; Niesters, H.G.; Cornelissen, J.J.; Puchhammer-Stöckl, E. Characterization of Epstein-Barr virus Type I variants based on linked polymorphism among EBNA3A, -3B, and-3C genes. Virus Res. 2006, 118, 105–114. [Google Scholar] [CrossRef]
- White, R.E.; Rämer, P.C.; Naresh, K.N.; Meixlsperger, S.; Pinaud, L.; Rooney, C.; Savoldo, B.; Coutinho, R.; Bödör, C.; Gribben, J.; et al. EBNA3B-deficient EBV promotes B cell lymphomagenesis in humanized mice and is found in human tumors. J. Clin. Investig. 2012, 122, 1487–1502. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.G.; Chang, K.L.; Chen, W.-G.; Chen, Y.-Y.; Shibata, D.; Hayashi, K.; Bacchi, C.; Bacchi, M.; Weiss, L.M. Epstein-Barr Virus (EBV) Nuclear Antigen (EBNA)-4 Mutation in EBV-Associated Malignancies in Three Different Populations. Am. J. Pathol. 1999, 155, 941–947. [Google Scholar] [CrossRef]
- Hui, K.F.; Chan, T.F.; Yang, W.; Shen, J.J.; Lam, K.P.; Kwok, H.; Sham, P.C.; Tsao, S.W.; Kwong, D.L.; Lung, M.L.; et al. High risk Epstein-Barr virus variants characterized by distinct polymorphisms in the EBER locus are strongly associated with nasopharyngeal carcinoma. Int. J. Cancer 2018, 144, 3031–3042. [Google Scholar] [CrossRef]
- Palser, A.L.; Grayson, N.E.; White, R.E.; Corton, C.; Correia, S.; Ba Abdullah, M.M.; Watson, S.J.; Cotten, M.; Arrand, J.R.; Murray, P.G.; et al. Genome Diversity of Epstein-Barr Virus from Multiple Tumor Types and Normal Infection. J. Virol. 2015, 89, 5222–5237. [Google Scholar] [CrossRef]
- Peng, R.-J.; Han, B.-W.; Cai, Q.-Q.; Zuo, X.-Y.; Xia, T.; Chen, J.-R.; Feng, L.-N.; Lim, J.Q.; Chen, S.-W.; Zeng, M.-S.; et al. Genomic and transcriptomic landscapes of Epstein-Barr virus in extranodal natural killer T-cell lymphoma. Leukemia 2018, 33, 1451–1462. [Google Scholar] [CrossRef]
- Kwok, H.; Chan, K.W.; Chan, K.H.; Chiang AK, S. Distribution, persistence and interchange of Epstein-Barr virus strains among PBMC, plasma and saliva of primary infection subjects. PLoS ONE 2015, 10, e0120710. [Google Scholar] [CrossRef]
- Monteiro, T.A.F.; Costa, I.B.; Costa, I.B.; dos Santos Corrêa, T.L.; Coelho, B.M.R.; Polaro, A.A.; da Silva, A.E.S.; de Paula Ramos, F.L.; Martins Filho, A.J.; Monteiro, J.L.F.; et al. Genotypes of Epstein–Barr virus (EBV1/EBV2) in individuals with infectious mononucleosis in the metropolitan area of Belém, Brazil, between 2005 and 2016. Braz. J. Infect. Dis. 2020, 24, 322–329. [Google Scholar] [CrossRef]
- Tabibzadeh, A.; Niya, M.H.K.; Esghaei, M.; Bokharaei-Salim, F.; Ataei-Pirkooh, A.; Kiani, S.J.; Monavari, S.H.R. Molecular Epidemiology of Epstein-Barr virus (EBV) in Patients with Hematologic Malignancies. Asian Pac. J. Cancer Prev. 2020, 21, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, Y.; Liu, X.; Chao, Y.; Xing, X.; Zhao, C.; Liu, C.; Luo, B. Genotypic Analysis of Epstein-Barr Virus Isolates Associated with Nasopharyngeal Carcinoma in Northern China. Intervirology 2011, 54, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Smatti, M.K.; Yassine, H.M.; AbuOdeh, R.; AlMarawani, A.; Taleb, S.A.; Althani, A.A.; Nasrallah, G.K. Prevalence and molecular profiling of Epstein Barr virus (EBV) among healthy blood donors from different nationalities in Qatar. PLoS ONE 2017, 12, e0189033. [Google Scholar] [CrossRef] [PubMed]
- Smatti, M.K.; Al-Sadeq, D.W.; Ali, N.H.; Pintus, G.; Abou-Saleh, H.; Nasrallah, G.K. Epstein–Barr Virus Epidemiology, Serology, and Genetic Variability of LMP-1 Oncogene Among Healthy Population: An Update. Front. Oncol. 2018, 8, 211. [Google Scholar] [CrossRef]
- Sixbey, J.; Chesney, P.; Shirley, P.; Buntin, D.; Resnick, L. Detection of a second widespread strain of epstein-barr virus. Lancet 1989, 334, 761–765. [Google Scholar] [CrossRef]
- Lin, J.C.; Lin, S.C.; De, B.K.; Chan, W.C.; Evatt, B.L.; Chan, W.P. Precision of genotyping of Epstein-Barr virus by polymerase chain reaction using three gene loci (EBNA-2, EBNA-3C, and EBER): Predominance of type A virus associated with Hodgkin’s disease. Blood 1993, 81, 3372–3381. [Google Scholar]
- Pereira, L.M.S.; França, E.d.S.; Costa, I.B.; Lima, I.T.; Freire, A.B.C.; Ramos, F.L.d.P.; Monteiro, T.A.F.; Macedo, O.; Sousa, R.C.M.; Freitas, F.B.; et al. Epstein–Barr Virus (EBV) Genotypes Associated with the Immunopathological Profile of People Living with HIV-1: Immunological Aspects of Primary EBV Infection. Viruses 2022, 14, 168. [Google Scholar] [CrossRef]
- Ayee, R.; Ofori, M.E.O.; Tagoe, E.A.; Languon, S.; Searyoh, K.; Armooh, L.; Bilson-Amoah, E.; Baidoo, K.; Kitcher, E.; Wright, E.; et al. Genotypic Characterization of Epstein Barr Virus in Blood of Patients with Suspected Nasopharyngeal Carcinoma in Ghana. Viruses 2020, 12, 766. [Google Scholar] [CrossRef]
- Tinguely, M.; A Bründler, M.; Gogos, S.; Kerl, K.; Borisch, B. Epstein-Barr virus association in pediatric abdominal non-Hodgkin-lymphomas from Turkey. Arch. Immunol. Ther. Exp. 2000, 48, 317–322. [Google Scholar]
- Ibrahim, H.A.; Menasce, L.P.; Pomplun, S.; Burke, M.; Bower, M.; Naresh, K.N. Epstein–Barr virus (EBV) genotypes among human immunodeficiency virus (HIV)-related B-cell Lymphomas and B-cell post-transplant lymphoproliferative disorders (B-PTLD)–late-onset lymphomas, especially in the HIV setting, are associated with type-B-EBV. Eur. J. Haematol. 2010, 85, 227–230. [Google Scholar] [CrossRef]
- Chabay, P.A.; Preciado, M.V. EBV primary infection in childhood and its relation to B-cell lymphoma development: A mini-review from a developing region. Int. J. Cancer 2012, 133, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Renzette, N.; Somasundaran, M.; Brewster, F.; Coderre, J.; Weiss, E.R.; McManus, M.; Greenough, T.; Tabak, B.; Garber, M.; Kowalik, T.F.; et al. Epstein-Barr virus latent membrane protein 1 genetic variability in peripheral blood B cells and oropharyngeal fluids. J. Virol. 2014, 88, 3744–3755. [Google Scholar] [CrossRef] [PubMed]
- Trimèche, M.; Bonnet, C.; Korbi, S.; Boniver, J.; Leval, L.D. Association between Epstein-Barr virus and Hodgkin’s lymphoma in Belgium: A pathological and virological study. Leuk. Lymphoma 2007, 48, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, T.A.F.; Costa, I.B.; Costa, I.B.; dos Santos Corrêa, T.L.; Coelho, B.M.R.; Polaro, A.A.; da Silva, A.E.S.; de Paula Ramos, F.L.; Martins Filho, A.J.; Monteiro, J.L.F.; et al. Gene EBNA3C: Type of Infection by EBV (EBV1 and EBV2) Correlation with Clinical and Biochemical Parameters (AST, ALT and GGT) in Individuals with Infectious Mononucleosis in the Metropolitan Area of Belém, Pará, 2005–2016. Res. Sq. 2019. [Google Scholar] [CrossRef]
- Yao, Q.Y.; Tierney, R.J.; Croom-Carter, D.; Dukers, D.; Cooper, G.M.; Ellis, C.J.; Rowe, M.; Rickinson, A.B. Frequency of multiple Epstein-Barr virus infections in T-cell-immunocompromised individuals. J. Virol. 1996, 70, 4884–4894. [Google Scholar] [CrossRef]
- Correa, R.M.; Fellner, M.D.; Durand, K.; Redini, L.; Alonio, V.; Yampolsky, C.; Colobraro, A.; Sevlever, G.; Teyssié, A.; Benetucci, J.; et al. Epstein Barr virus genotypes and LMP-1 variants in HIV-infected patients. J. Med. Virol. 2007, 79, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Chen, Y.; Hui, J.; Guo, Y.; Peng, X.; Wang, M.; Hu, C.; Xie, Y.; Su, J.; Huang, Y.; et al. Epstein-Barr virus variation in people living with human immunodeficiency virus in southeastern China. Virol. J. 2023, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Number | Percent |
|---|---|---|
| Study participants (n = 207) | ||
| FFPE blocks | 63 | 30.4 |
| Lymphoma patients | 144 | 69.6 |
| Sex | ||
| Male | 135 | 65.2 |
| Female | 72 | 34.8 |
| Age | ||
| <20 years | 32 | 15.5 |
| 21–40 years | 79 | 38.2 |
| 41–60 years | 75 | 36.2 |
| >61 years | 20 | 9.7 |
| Not applicable | 1 | 0.5 |
| HIV status | ||
| Positive | 28 | 13.5 |
| Negative | 98 | 47.3 |
| Unknown | 81 | 39.2 |
| Lymphoma types | ||
| Hodgkin’s lymphoma | 70 | 33.8 |
| Non-Hodgkin’s lymphoma | 137 | 66.2 |
| Characteristics | EBV1 | EBV2 | Coinfections | Total | p-Value |
|---|---|---|---|---|---|
| Age <20 years 21–40 years 41–60 years >60 years Not applicable | 20 (62.5%) 39 (49.4%) 36 (48%) 12 (60%) 1 | 11 (34.4%) 37 (46.8%) 25 (33.3%) 6 (30%) | 1 (3.1%) 3 (3.8%) 14 (18.7%) 2 (10%) | 32 79 75 20 1 | 0.027 |
| Sex Male Female | 70 (51.8%) 38 (52.8%) | 52 (38.5%) 27 (37.5%) | 13(9.6%) 7(9.7%) | 135 72 | 0.99 |
| HIV status HIV positive HIV negative Not applicable | 13 (46.4%) 45 (45.9%) 50 (61.7%) | 12 (42.9%) 42 (42.9%) 25 (30.9%) | 3 (10.7%) 11 (11.2%) 6 (7.4%) | 28 98 81 | 0.169 |
| Lymphoma type HL NHL | 37 (52.9%) 71 (51.8%) | 30 (42.6%) 49 (35.7% | 3 (4.3%) 17 (12.4%) | 70 137 | 0.15 |
| PCR Reactions | Forward | Reverse |
|---|---|---|
| EBNA1 | TCATCATCATCCGGGTCTCC | CCTACAGGGTGGAAAAATGGC |
| ACTB | TCGTGCGTGACATTAAGGAG | CAGGCAGCTCGTAGCTCTTC |
| EBNA3C | AGAAGGGGAGCGTGTGTTG | GGCTCGTTTTTGACGTCGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teshome, S.; Ahmed, E.H.; Zealiyas, K.; Abubeker, A.; Tadesse, F.; Weigel, C.; Baiocchi, R.A.; Abebe, T. Genotypes Distribution of Epstein–Barr Virus among Lymphoma Patients in Ethiopia. Int. J. Mol. Sci. 2023, 24, 13891. https://doi.org/10.3390/ijms241813891
Teshome S, Ahmed EH, Zealiyas K, Abubeker A, Tadesse F, Weigel C, Baiocchi RA, Abebe T. Genotypes Distribution of Epstein–Barr Virus among Lymphoma Patients in Ethiopia. International Journal of Molecular Sciences. 2023; 24(18):13891. https://doi.org/10.3390/ijms241813891
Chicago/Turabian StyleTeshome, Seifegebriel, Elshafa Hassan Ahmed, Kidist Zealiyas, Abdulaziz Abubeker, Fisihatsion Tadesse, Christoph Weigel, Robert A. Baiocchi, and Tamrat Abebe. 2023. "Genotypes Distribution of Epstein–Barr Virus among Lymphoma Patients in Ethiopia" International Journal of Molecular Sciences 24, no. 18: 13891. https://doi.org/10.3390/ijms241813891
APA StyleTeshome, S., Ahmed, E. H., Zealiyas, K., Abubeker, A., Tadesse, F., Weigel, C., Baiocchi, R. A., & Abebe, T. (2023). Genotypes Distribution of Epstein–Barr Virus among Lymphoma Patients in Ethiopia. International Journal of Molecular Sciences, 24(18), 13891. https://doi.org/10.3390/ijms241813891