

Effects of Biological Nitrogen Metabolism on Glufosinate-Susceptible and -Resistant Goosegrass (Eleusine indica L.)

 ,
,
Abstract
:1. Introduction
2. Results
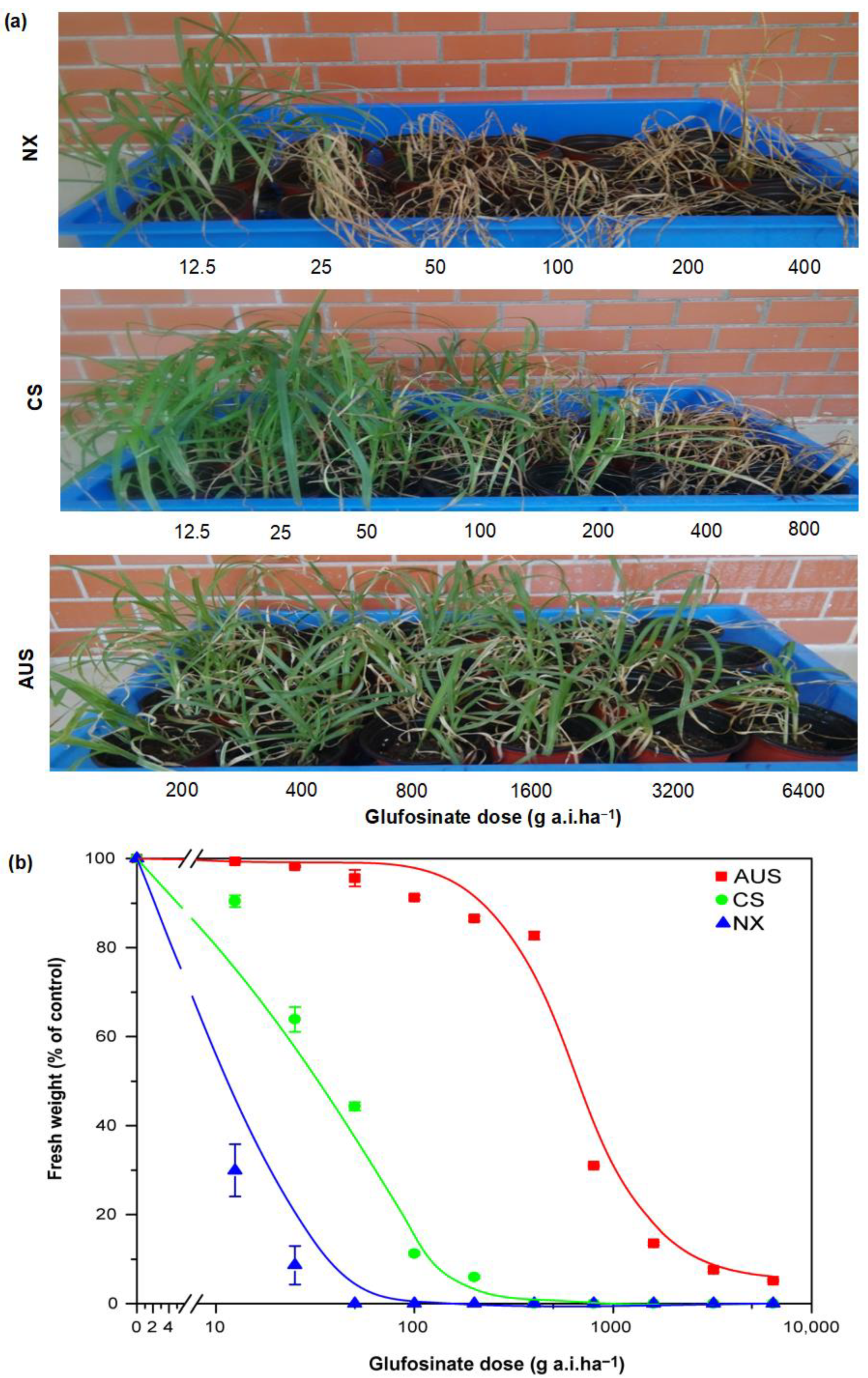
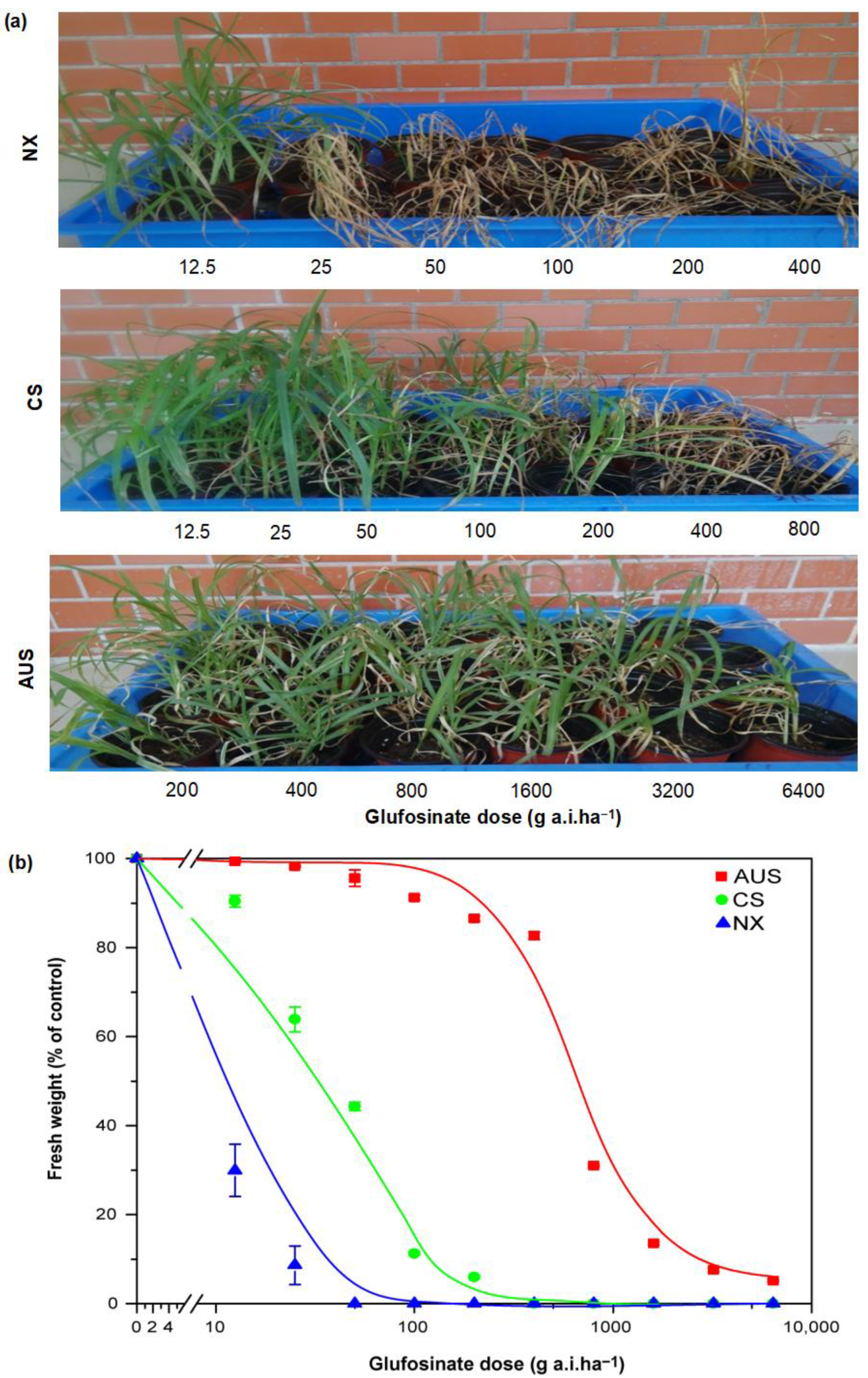
2.1. Resistant Phenotypes and Dose Responses to Glufosinate Herbicides
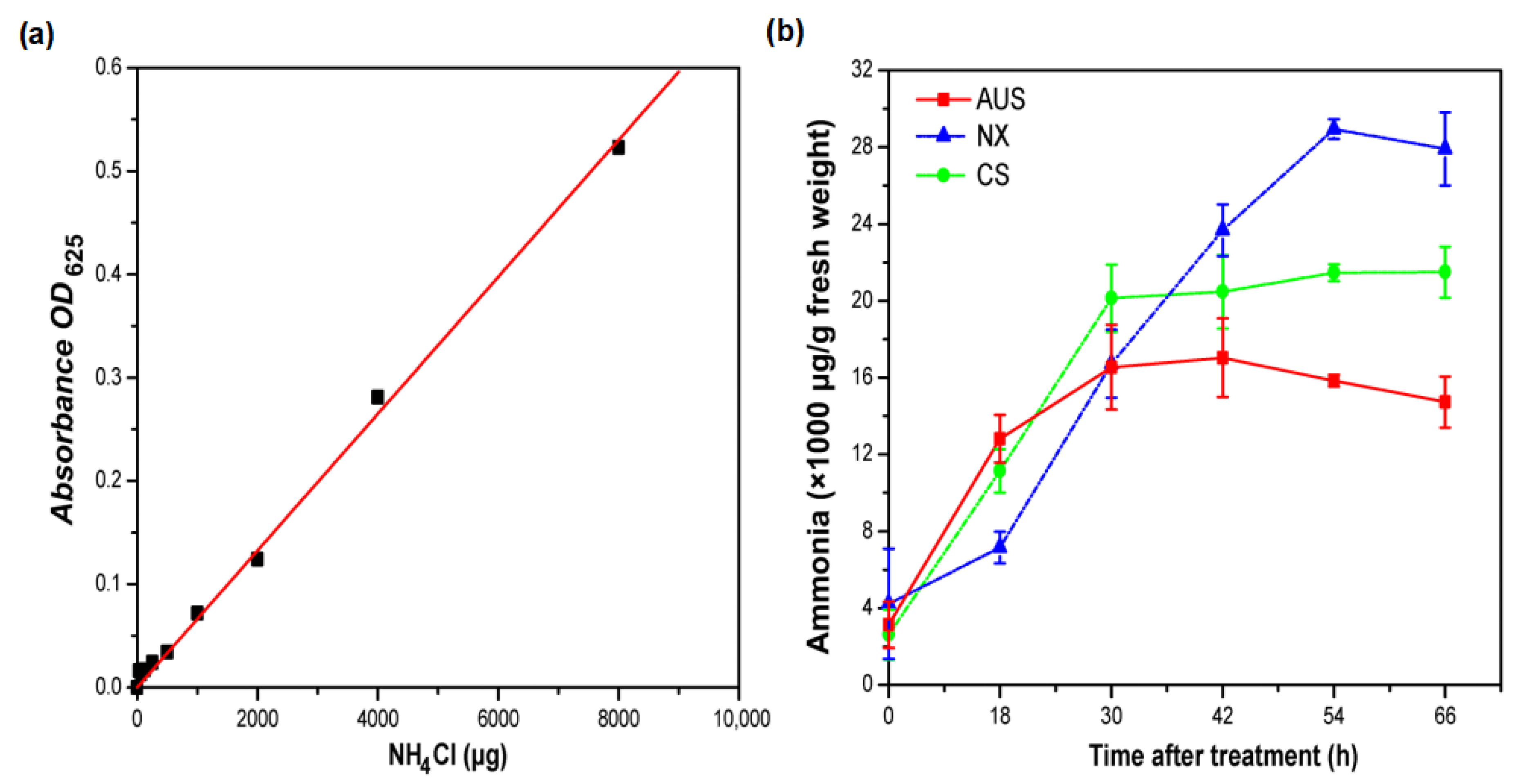
2.2. Glufosinate-Resistant Effects on Ammonia Content
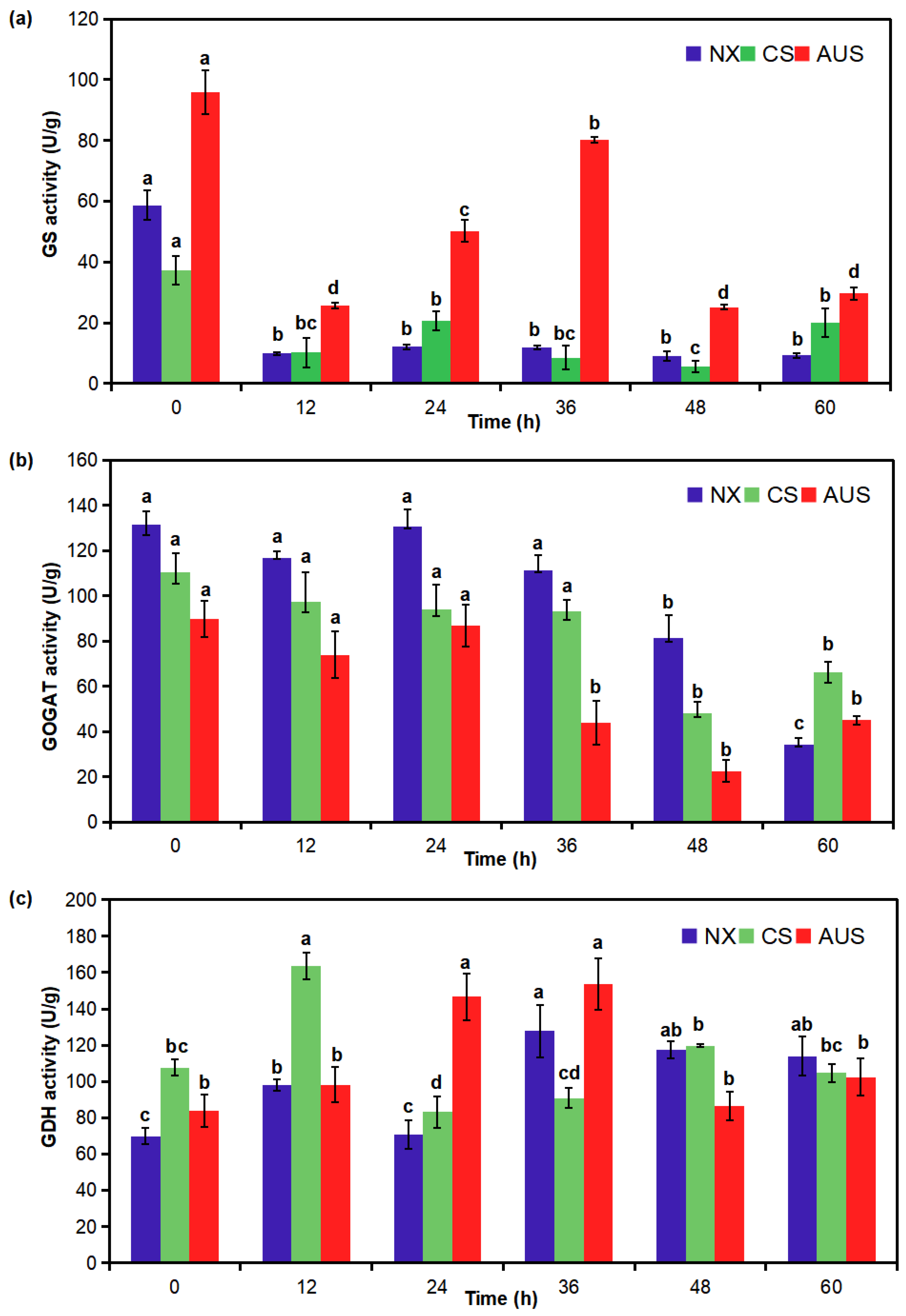
2.3. Glufosinate-Resistant Effects on GS, GOGAT, and GDH Activities
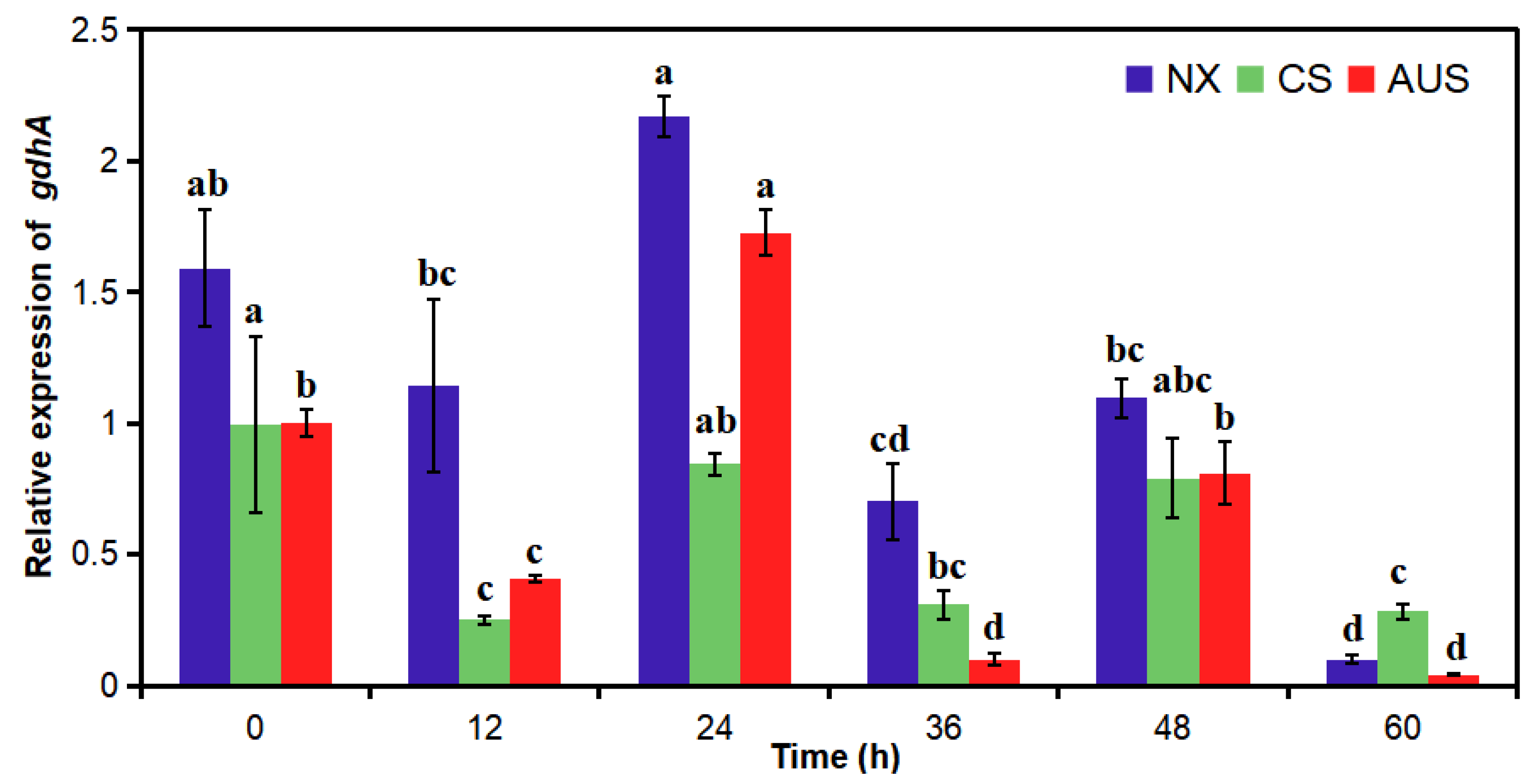
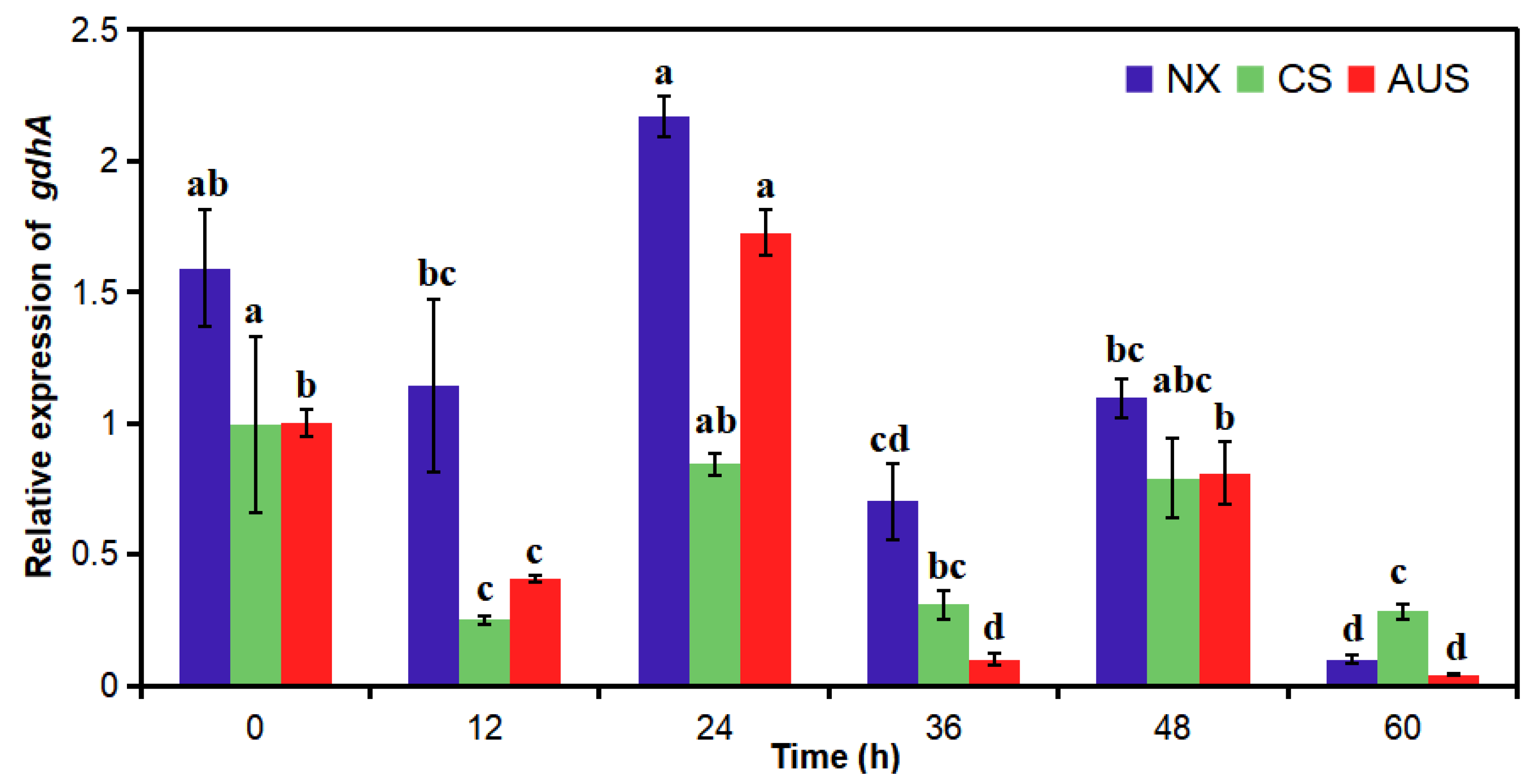
2.4. Relative gdhA Gene Expression by RT-PCR
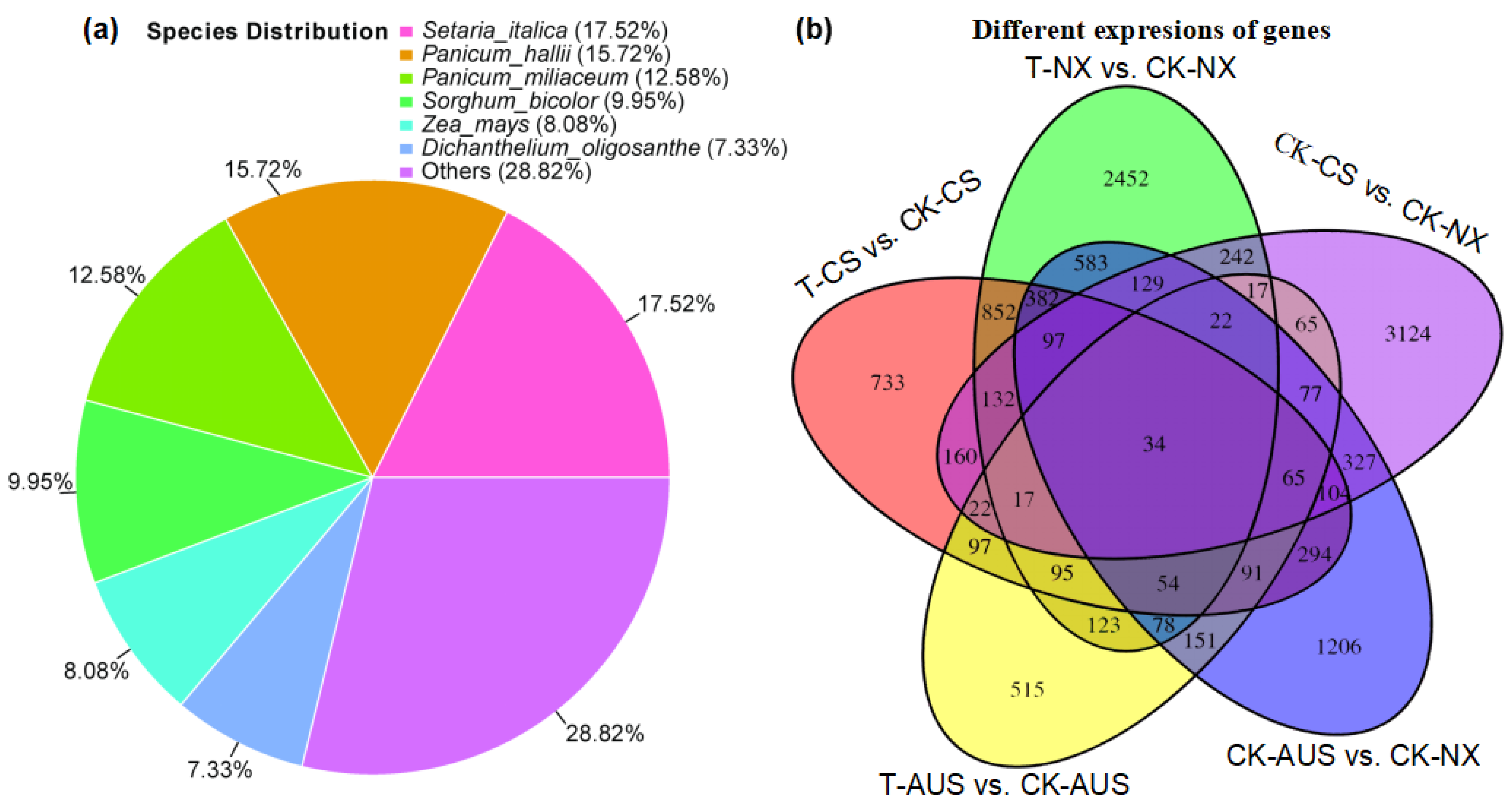
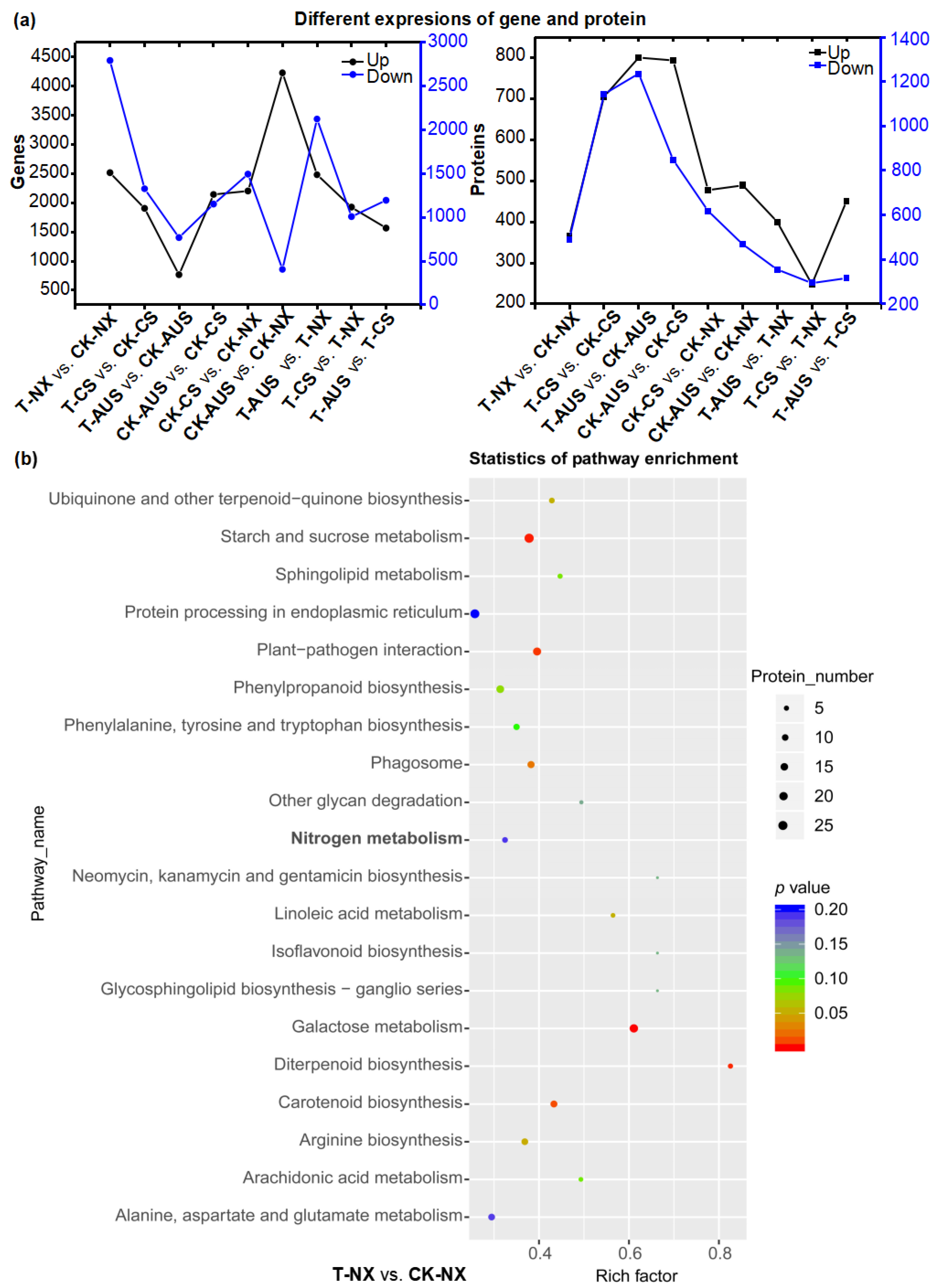
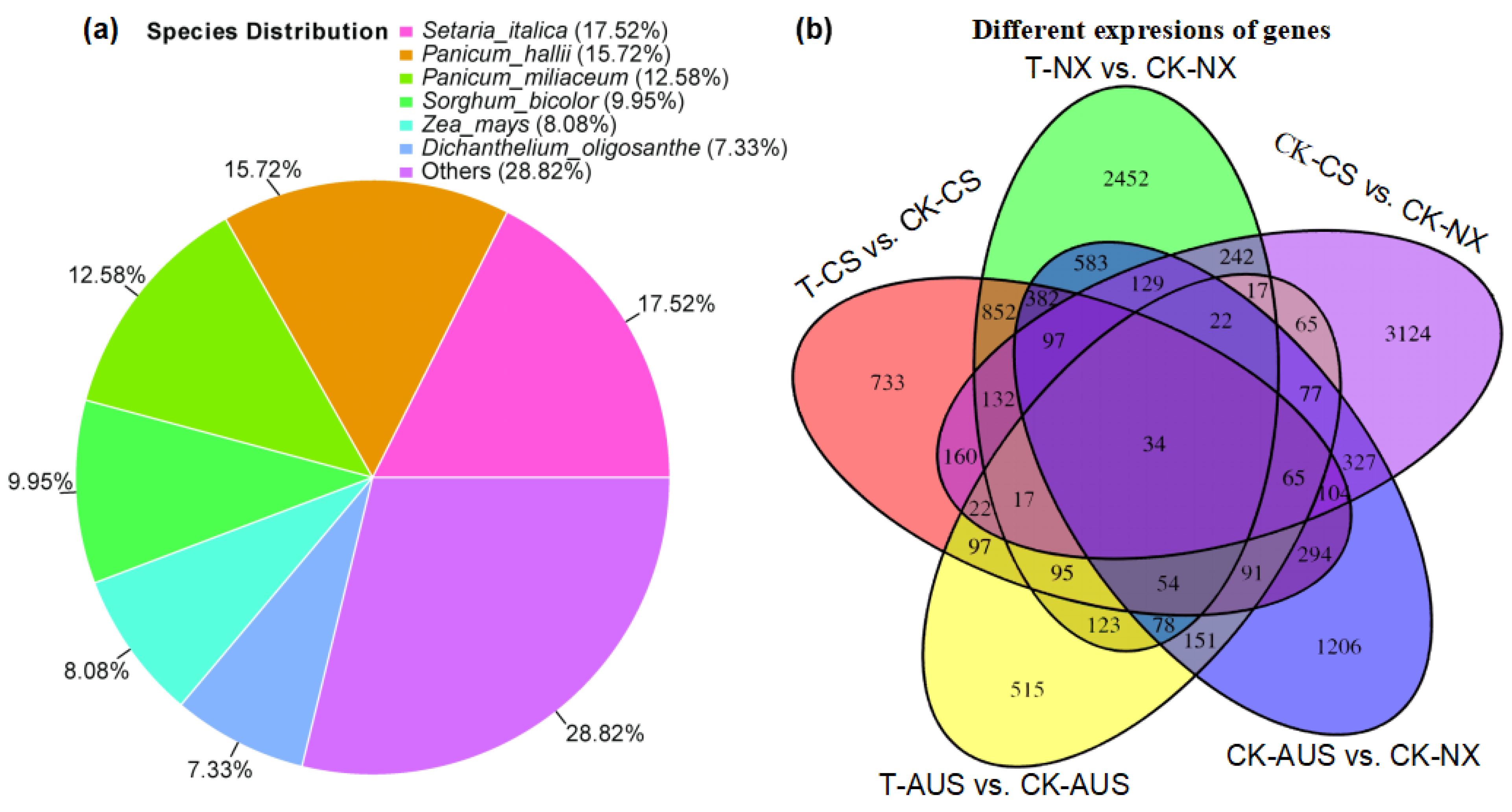
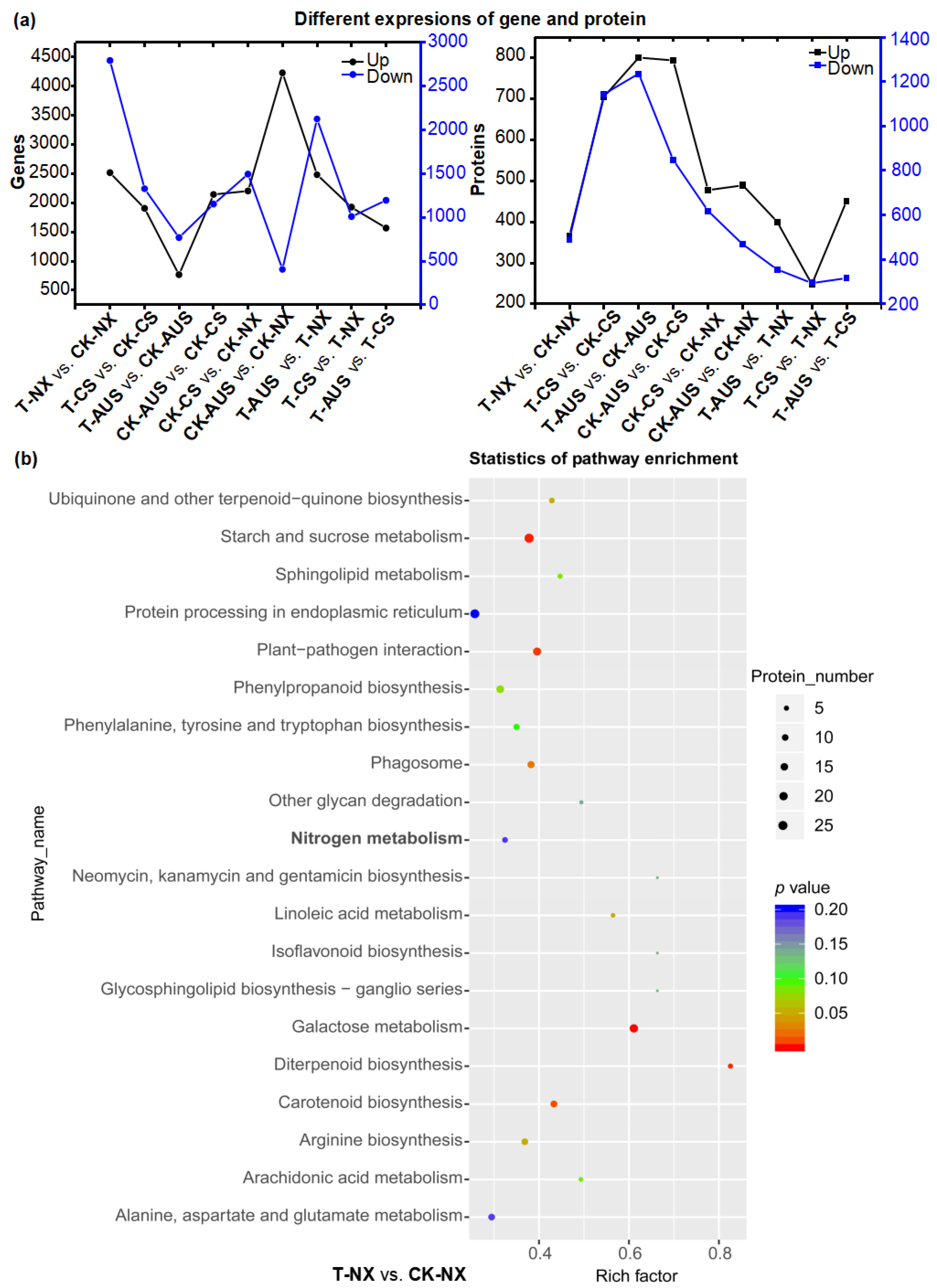
2.5. Effects of Glufosinate on Transcriptome Characterizations
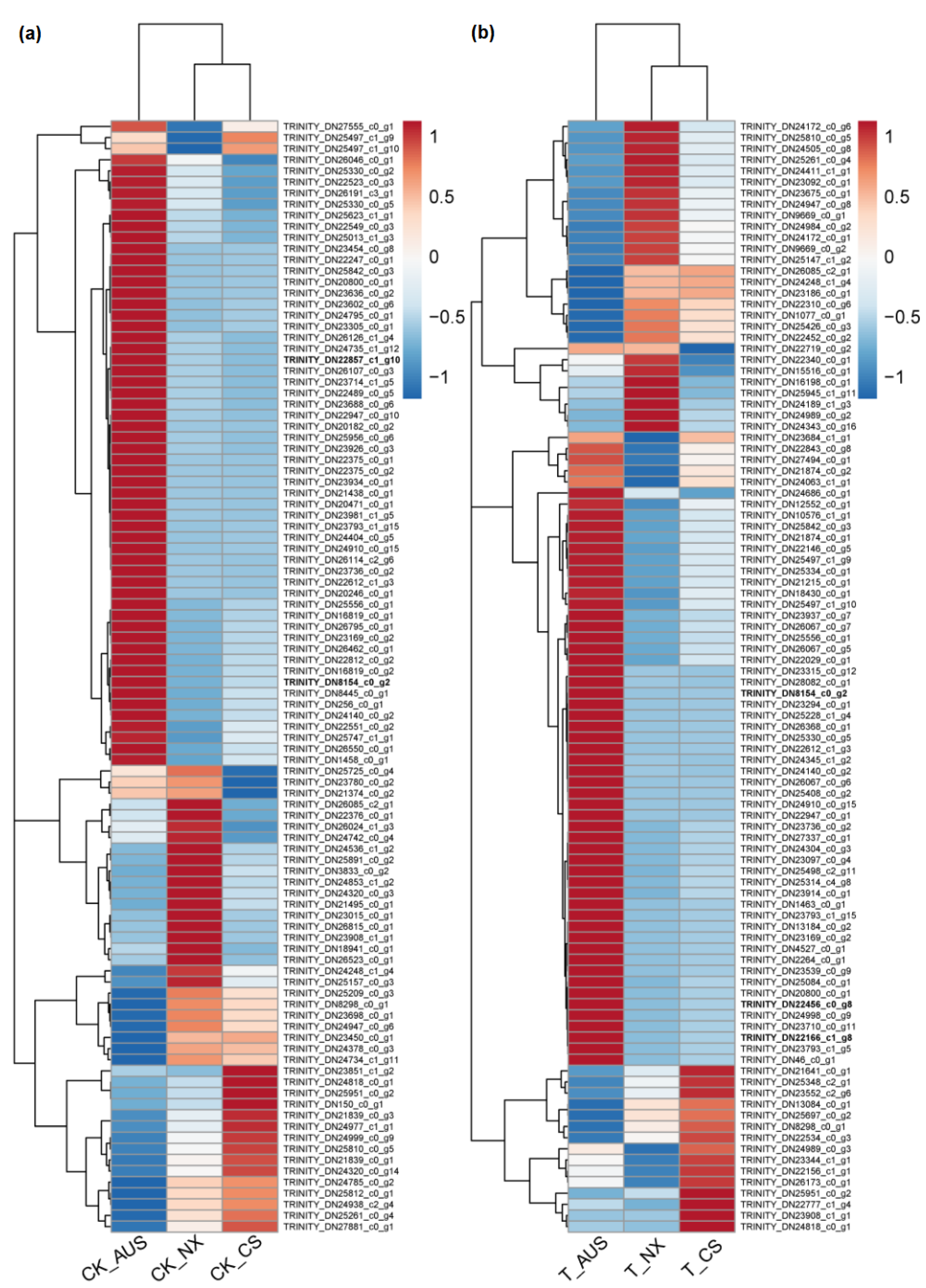
2.6. Effects of Glufosinate on Proteomic Characterizations
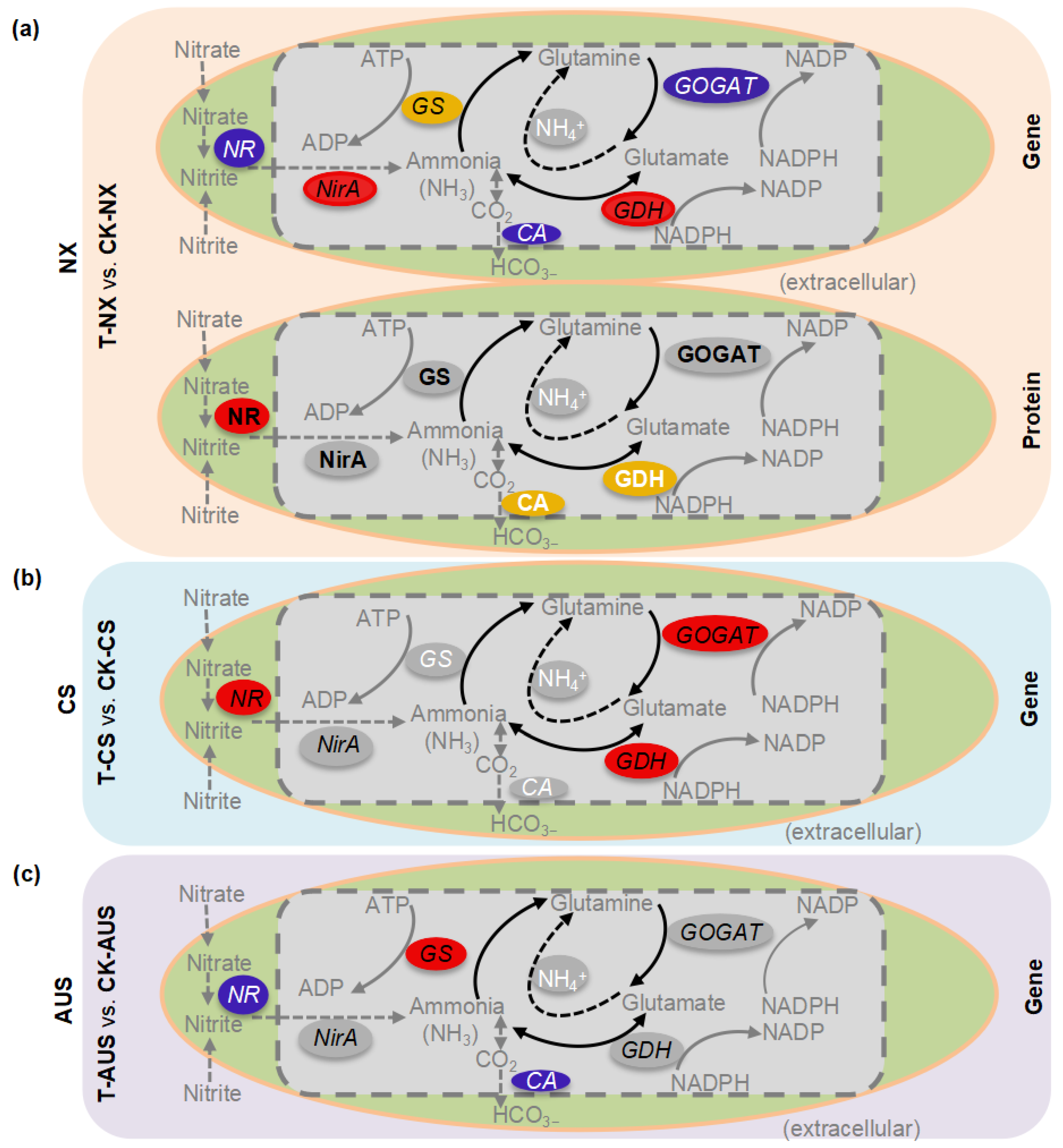
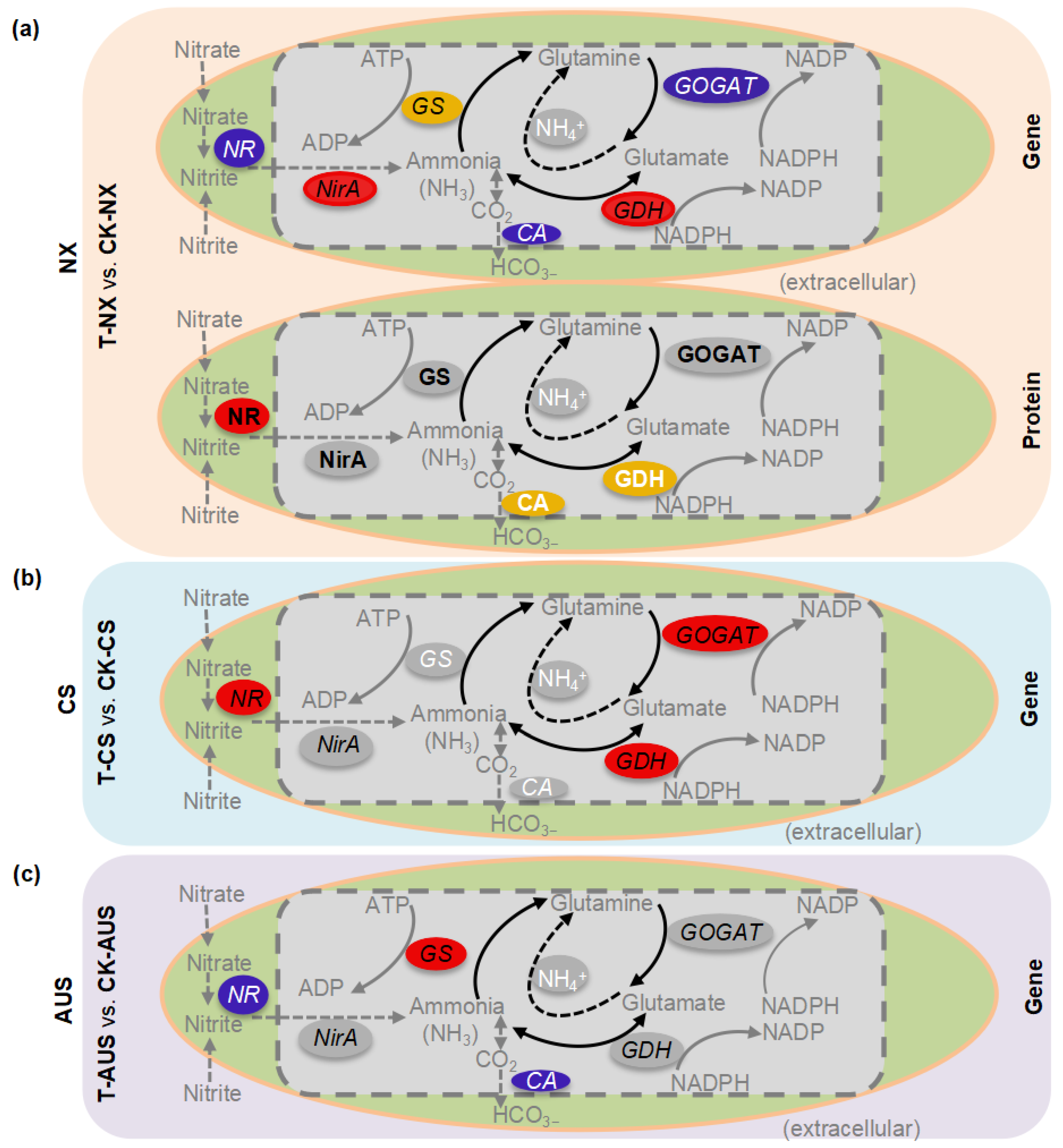
2.7. The Patterns of Expressed Genes and Proteins in the Nitrogen Metabolism
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. GR50 of the Glufosinate Treatments
4.3. Ammonium Content
4.4. qRT-PCR
4.5. Assay Enzyme Activity of GS, GOGAT, and GDH
4.6. Illumina Sequencing and iTRAQ-TMT Proteome Analysis
4.7. Bioinformatics Analyses
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, D.; Borphukan, B.; Fartyal, D.; Ram, B.; Singh, J.; Manna, M.; Sheri, V.; Panditi, V.; Yadav, R.; Achary, M.M.; et al. Concurrent overexpression of OsGS1-1 and OsGS2 genes in transgenic rice (Oryza sativa L.): Impact on tolerance to abiotic stresses. Front. Plant Sci. 2018, 9, 786. [Google Scholar] [CrossRef] [PubMed]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Burns, E.E.; Keith, B.K.; Refai, M.Y.; Bothner, B.; Dyer, W.E. Proteomic and biochemical assays of glutathione-related proteins in susceptible and multiple herbicide resistant Avena fatua L. Pest. Biochem. Physiol. 2017, 140, 69–78. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 28 February 2023).
- Cummins, I.; Dixon, D.P.; Freitag-Pohl, S.; Skipsey, M.; Edwards, R. Multiple roles for plant glutathione transferases in xenobiotic detoxification. Drug Metab. Rev. 2011, 43, 266–280. [Google Scholar] [CrossRef]
- Yu, H.D.; Zhang, Y.M.; Zhang, Z.Y.; Zhang, J.; Wei, Y.H.; Jia, X.T.; Wang, X.C.; Ma, X.M. Towards identification of molecular mechanism in which the overexpression of wheat cytosolic and plastid glutamine synthetases in tobacco enhanced drought tolerance. Plant Physiol. Biochem. 2020, 151, 608–620. [Google Scholar] [CrossRef]
- Gherekhloo, J.; Hassanpour-Bourkheili, S.; Hejazirad, P.; Golmohammadzadeh, S.; Vazquez-Garcia, J.G.; De Prado, R. Herbicide resistance in phalaris species: A review. Plants 2021, 10, 2248. [Google Scholar] [CrossRef] [PubMed]
- Avila-Garcia, W.V.; Sanchez-Olguin, E.; Hulting, A.G.; Mallory-Smith, C. Target-site mutation associated with glufosinate resistance in Italian ryegrass (Lolium perenne L. ssp. multiflorum). Pest Manag. Sci. 2012, 68, 1248–1254. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, Q.; Han, H.P.; Yu, C.J.; Nyporko, A.; Tian, X.S.; Beckie, H.; Powles, S. A naturally evolved mutation (Ser59Gly) in glutamine synthetase confers glufosinate resistance in plants. J. Exp. Bot. 2022, 73, 2251–2262. [Google Scholar] [CrossRef]
- Takano, H.K.; Beffa, R.; Preston, C.; Westra, P.; Dayan, F.E. A novel insight into the mode of action of glufosinate: How reactive oxygen species are formed. Photosynth. Res. 2020, 144, 361–372. [Google Scholar] [CrossRef]
- Takano, H.K.; Dayan, F.E. Glufosinate-ammonium: A review of the current state of knowledge. Pest Manag. Sci. 2020, 76, 3911–3925. [Google Scholar] [CrossRef]
- Unno, H.; Uchida, T.; Sugawara, H.; Kurisu, G.; Sugiyama, T.; Yamaya, T.; Sakakibara, H.; Hase, T.; Kusunoki, M. Atomic structure of plant glutamine synthetase: A key enzyme for plant productivity. J. Biol. Chem. 2006, 281, 29287–29296. [Google Scholar] [CrossRef] [PubMed]
- Salas-Perez, R.A.; Saski, C.A.; Noorai, R.E.; Srivastava, S.K.; Lawton-Rauh, A.L.; Nichols, R.L.; Roma-Burgos, N. RNA-Seq transcriptome analysis of Amaranthus palmeri with differential tolerance to glufosinate herbicide. PLoS ONE 2018, 13, e0195488. [Google Scholar] [CrossRef]
- Brunharo, C.A.C.G.; Takano, H.K.; Mallory-Smith, C.A.; Dayan, F.E.; Hanson, B.D. Role of glutamine synthetase isogenes and herbicide metabolism in the mechanism of resistance to glufosinate in Lolium perenne L. spp. multiflorum biotypes from oregon. J. Agric. Food Chem. 2019, 67, 8431–8440. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.K.; Beffa, R.; Preston, C.; Westra, P.; Dayan, F.E. Reactive oxygen species trigger the fast action of glufosinate. Planta 2019, 249, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.P.; Lima, L.M.; Santos, I.M.; Carvalho, H.G.; Cullimore, J.V. Expression of the Plastid-Located Glutamine Synthetase of Medicago truncatula. Accumulation of the Precursor in Root Nodules Reveals an in Vivo Control at the Level of Protein Import into Plastids. Plant Physiol. 2003, 132, 390–399. [Google Scholar] [CrossRef]
- Melooliveira, R.; Oliveira, I.C.; Coruzzi, G.M. Arabidopsis mutant analysis and gene regulation define a nonredundant role for glutamate dehydrogenase in nitrogen assimilation. Proc. Natl. Acad. Sci. USA 1996, 93, 4718–4723. [Google Scholar] [CrossRef]
- Liang, T.; Yuan, Z.Q.; Fu, L.; Zhu, M.H.; Luo, X.Y.; Xu, W.W.; Yuan, H.R.; Zhu, R.S.; Hu, Z.L.; Wu, X.T. Integrative transcriptomic and proteomic analysis reveals an alternative molecular network of glutamine synthetase 2 corresponding to nitrogen deficiency in rice (Oryza sativa L.). Int. J. Mol. Sci. 2021, 22, 7674. [Google Scholar] [CrossRef]
- Marino, D.; Cañas, R.A.; Betti, M. Is plastidic glutamine synthetase essential for C3 plants? A tale of photorespiratory mutants, ammonium tolerance and conifers. New Phytol. 2022, 234, 1559–1565. [Google Scholar] [CrossRef]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J. Exp. Bot. 2002, 370, 979–987. [Google Scholar] [CrossRef]
- Nolte, S.A.; Young, B.G.; Mungur, R.; Lightfoot, D.A. The glutamate dehydrogenase gene gdhA increased the resistance of tobacco to glufosinate. Weed Res. 2004, 44, 335–339. [Google Scholar] [CrossRef]
- Kurniadie, D.; Widianto, R.; Umiyati, U.; Widayat, D.; Nasahi, C.; Budiawan, A. Management of Eleusine indica (L.) Gaertn Resistance to Glyphosate Herbicide in Indonesia. Agronomy 2023, 13, 1649. [Google Scholar] [CrossRef]
- Corbett, J.L.; Askew, S.D.; Thomas, W.E.; Wilcut, J.W. Weed efficacy evaluations for bromoxynil, glufosinate, glyphosate, pyrithiobac, and sulfosate. Weed Technol. 2004, 18, 443–453. [Google Scholar] [CrossRef]
- Jalaludin, A.; Ngim, J.; Bakar, B.H.J.; Alias, Z. Preliminary findings of potentially resistant goosegrass (Eleusine indica) to glufosinate-ammonium in Malaysia. Weed Biol. Manag. 2010, 10, 256–260. [Google Scholar] [CrossRef]
- Jalaludin, A.; Yu, Q.; Powles, S.B. Multiple resistance across glufosinate, glyphosate, paraquat and ACCase-inhibiting herbicides in an Eleusine indica population. Weed Res. 2015, 55, 82–89. [Google Scholar] [CrossRef]
- Seng, C.T.; Lun, L.V.; San, C.T.; Sahid, I.B. Initial report of glufosinate and paraquat multiple resistance that evolved in a biotype of goosegrass (Eleusine indica) in Malaysia. Weed Biol. Manag. 2010, 10, 229–233. [Google Scholar] [CrossRef]
- Rojano-Delgado, A.M.; Priego-Capote, F.; Barro, F.; de Castro, M.D.L.; De Prado, R. Liquid chromatography-diode array detection to study the metabolism of glufosinate in Triticum aestivum T-590 and influence of the genetic modification on its resistance. Phytochemistry 2013, 96, 117–122. [Google Scholar] [CrossRef]
- Avila-Garcia, W.V.; Carol, M.S. Glyphosate-resistant Italian Ryegrass (Lolium perenne) populations also exhibit resistance to glufosinate. Weed Sci. 2011, 59, 305–309. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harrington, K.C.; James, T.K. Glyphosate-resistant Lolium multiflorum and Lolium perenne populations from New Zealand are also resistant to glufosinate and amitrole. Crop Prot. 2015, 78, 1–4. [Google Scholar] [CrossRef]
- Karn, E.; Beffa, R.; Jasieniuk, M. Variation in response and resistance to glyphosate and glufosinate in California populations of Italian ryegrass (Lolium perenne ssp. multiflorum). Weed Sci. 2018, 66, 168–179. [Google Scholar] [CrossRef]
- Travlos, I.S.; Cheimona, N.; De Prado, R.; Jhala, A.J.; Chachalis, D.; Tani, E. First case of glufosinate-resistant rigid ryegrass (Lolium rigidum Gaud.) in Greece. Agronomy 2018, 8, 35. [Google Scholar] [CrossRef]
- Carvalho-Moore, P.; Norsworthy, J.K.; González-Torralva, F.; Hwang, J.I.; Patel, J.D.; Barber, L.T.; Butts, T.R.; McElroy, J.S. Unraveling the mechanism of resistance in a glufosinate-resistant Palmer amaranth (Amaranthus palmeri) accession. Weed Sci. 2022, 70, 370–379. [Google Scholar] [CrossRef]
- Takano, H.K.; Beffa, R.; Preston, C.; Westra, P.; Dayan, F.E. Glufosinate enhances the activity of protoporphyrinogen oxidase inhibitors. Weed Sci. 2020, 68, 324–332. [Google Scholar] [CrossRef]
- Everman, W.J.; Burke, I.C.; Allen, J.R.; Collins, J.; Wilcut, J.W. Weed control and yield with glufosinate-resistant cotton weed management systems. Weed Technol. 2007, 21, 695–701. [Google Scholar] [CrossRef]
- Everman, W.J.; Clewis, S.B.; York, A.C.; Wilcut, J.W. Weed control and yield with flumioxazin, fomesafen, and S-metolachlor systems for glufosinate-resistant cotton residual weed management. Weed Technol. 2009, 23, 391–397. [Google Scholar] [CrossRef]
- Everman, W.J.; Mayhew, C.R.; Burton, J.D.; York, A.C.; Wilcut, J.W. Absorption, translocation, and metabolism of 14C-glufosinate in glufosinate-resistant corn, goosegrass (Eleusine indica), large crabgrass (Digitaria sanguinalis), and sicklepod (Senna obtusifolia). Weed Sci. 2009, 57, 1–5. [Google Scholar] [CrossRef]
- Sharpe, S.M.; Boyd, N.S. Utility of glufosinate in postemergence row middle weed control in Florida plasticulture production. Weed Technol. 2019, 33, 495–502. [Google Scholar] [CrossRef]
- Fujita, T.; Beier, M.P.; Tabuchi-Kobayashi, M.; Hayatsu, Y.; Nakamura, H.; Umetsu-Ohashi, T.; Sasaki, K.; Ishiyama, K.; Murozuka, E.; Kojima, M.; et al. Cytosolic glutamine synthetase GS1;3 Is involved in rice grain ripening and germination. Front. Plant Sci. 2022, 13, 835835. [Google Scholar] [CrossRef]
- Olas, J.J.; Wahl, V. Tissue-specific NIA1 and NIA2 expression in Arabidopsis thaliana. Plant Signal. Behav. 2019, 14, 1656035. [Google Scholar] [CrossRef]
- Hines, K.M.; Chaudhari, V.; Edgeworth, K.N.; Owensb, T.G.; Hansona, M.R. Absence of carbonic anhydrase in chloroplasts affects C3 plant development but not photosynthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2107425118. [Google Scholar] [CrossRef]
- Burke, I.C.; Askew, S.D.; Corbett, J.L.; Wilcut, J.W. Glufosinate antagonizes clethodim control of goosegrass (Eleusine indica). Weed Technol. 2005, 19, 664–668. [Google Scholar] [CrossRef]
- Wilson, D.G.; York, A.C.; Jordan, D.L. Effect of row spacing on weed management in glufosinate-resistant cotton. Weed Technol. 2007, 21, 489–495. [Google Scholar] [CrossRef]
- Wolter, D.A.; Kyser, G.B.; Hanson, B.D. Herbicide Management of Threespike Goosegrass in California Orchards. HortTechnology 2023, 33, 176–180. [Google Scholar] [CrossRef]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Huang, Z.F.; Huang, H.J.; Wei, S.H.; Liu, Y.; Jiang, C.L.; Zhang, J.; Zhang, C.X. Selection of relatively exact reference genes for gene expression studies in goosegrass (Eleusine indica) under herbicide stress. Sci. Rep. 2017, 7, 46494. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.Y.; Wei, J.P.; Dong, Z.X.; Shen, X.F.; Chen, Y. Differences of endogenous polyamines and putative genes associated with paraquat resistance in goosegrass (Eleusine indica L.). PLoS ONE 2019, 14, e0216513. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Annotation (Species) | EC | KEGG_Pathway |
|---|---|---|---|
| CYP85A1 | Hypothetical protein GQ55_9G165800, Panicum hallii | EC:1.14.-.- | sita00906 (Brassinosteroid biosynthesis) |
| MLO1 | MLO-like protein 1, Dichanthelium oligosanthes | NA | NA |
| CER3 | Protein ECERIFERUM 3-like, Panicum hallii | EC:4.1.99.5 | smo00074 (Cutin, suberine, and wax biosynthesis) |
| GA20ox1B | Gibberellin 20 oxidase 1-D-like, Panicum miliaceum | EC:1.14.11.12 | sbi00905 (Diterpenoid biosynthesis) |
| Os04g0617900 | Germin-like protein 4-1, Setaria italica | NA | NA |
| Cht8 | Chitinase 8, Setaria italica | EC:3.2.1.14 | sita00520 (Amino sugar and nucleotide sugar metabolism) |
| Os04g0339400 | Probable aldo-keto reductase 3, Setaria italica | EC:1.1.1.65 | cre00750 (Vitamin B7 metabolism) |
| RPM1 | Hypothetical protein GQ55_2G362800, Panicum hallii | NA | bdi04627 (Plant–pathogen interaction) |
| PEP1 | Phosphoenolpyruvate carboxylase, Chloris gayana | EC:4.1.1.31 | sita00620 (Pyruvate metabolism) |
| At3g16150 | Probable isoaspartyl peptidase/L-asparaginase 2, Panicum hallii | NA | NA |
| LKR/SDH | Alpha-aminoadipic semialdehyde synthase isoform X1, Setaria italica | EC:1.5.1.8 1.5.1.9 | sita00311 (Lysine degradation) |
| Os03g0733400 | Zinc finger BED domain-containing protein ricesleeper 2-like, Setaria italica | EC:3.4.19.12 | bna04145 (Endocytosis) |
| At5g08350 | GEM-like protein 4, Panicum miliaceum | EC:2.3.2.32 | gmx04141 (Protein processing in endoplasmic reticulum) |
| LHCA4 | Chlorophyll a-b binding protein 4, chloroplastic, Sorghum bicolor | NA | sbi00197 (Photosynthesis-antenna proteins) |
| Protein | Majority Protein ID | Unique Peptides | Gene | Length | EC |
|---|---|---|---|---|---|
| GS | Gene.4172::TRINITY_DN15902_c0_g2::g.4172::m.4172 | 7 | fluG | 2716 | EC:6.3.1.2 |
| Gene.49488::TRINITY_DN25503_c1_g2::g.49488::m.49488 | 13 | GLN1-1 | 1414 | EC:6.3.1.2 | |
| Gene.49499::TRINITY_DN25503_c1_g6::g.49499::m.49499 | 13 | GLN1-2 | 1371 | EC:6.3.1.2 | |
| Gene.49492::TRINITY_DN25503_c1_g4::g.49492::m.49492 | 2 | GLN1-3 | 1487 | EC:6.3.1.2 | |
| Gene.51130::TRINITY_DN25703_c0_g2::g.51130::m.51130 | 10 | GLN2 | 2201 | EC:6.3.1.2 | |
| GDH | Gene.12781::TRINITY_DN20596_c0_g4::g.12781::m.12781 | 10 | GDH1 | 1508 | EC:1.4.1.3 |
| Gene.24624::TRINITY_DN23079_c2_g5::g.24624::m.24624 | 8 | GDH2 | 2083 | EC:1.4.1.3 | |
| GOGAT | Gene.45299::TRINITY_DN25064_c1_g1::g.45299::m.45299 | 25 | Os01g0681900 | 7729 | EC:1.4.1.13 1.4.1.14 |
| Gene.45304::TRINITY_DN25064_c1_g2::g.45304::m.45304 | 11 | Os05g0555600 | 6655 | EC:1.4.1.13 1.4.1.14 | |
| Gene.32477::TRINITY_DN23865_c0_g14::g.32477::m.32477 | 49 | GLSF | 5055 | EC:1.4.7.1 | |
| NR | Gene.41212::TRINITY_DN24663_c0_g1::g.41212::m.41212 | 5 | NIA1 | 3174 | EC:1.7.1.1 1.7.1.2 1.7.1.3 |
| Gene.51843::TRINITY_DN25791_c1_g6::g.51843::m.51843 | 1 | CB5-A | 879 | EC:1.7.1.1 1.7.1.2 1.7.1.3 | |
| NirA | Gene.11975::TRINITY_DN20338_c0_g2::g.11975::m.11975 | 19 | Os01g0357100 | 2060 | EC:1.7.7.1 |
| CA | Gene.23527::TRINITY_DN22941_c0_g1::g.23527::m.23527 | 12 | CA1 | 932 | EC:4.2.1.1 |
| Gene.23538::TRINITY_DN22941_c0_g5::g.23538::m.23538 | 4 | - | 534 | EC:4.2.1.1 | |
| Gene.42475::TRINITY_DN24781_c0_g1::g.42475::m.42475 | 1 | - | 860 | EC:4.2.1.1 |
| Proteome | Norm | Fold | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK_NX | CK_CS | CK_AUS | T_NX | T_CS | T_AUS | CK_CS/ CK_NX | CK_AUS/CK_NX | CK_AUS/CK_CS | T_CS/ T_NX | T_AUS/ T_NX | T_AUS/ T_CS | ||
| GS | fluG | 0.51 | 0.60 | 0.57 | 0.52 | 0.48 | 0.54 | 1.17 | 1.11 | 0.94 | 0.93 | 1.04 | 1.11 |
| GLN1-1 | 5.81 | 5.98 | 6.28 | 6.99 | 5.31 | 6.69 | 1.03 | 1.08 | 1.05 | 0.76 | 0.96 | 1.26 | |
| GLN1-2 | 2.46 | 2.78 | 3.63 | 3.05 | 2.99 | 3.43 | 1.13 | 1.47 | 1.31 | 0.98 | 1.12 | 1.15 | |
| GLN1-3 | 0.08 | 0.10 | 0.17 | 0.12 | 0.12 | 0.12 | 1.23 | 2.15 | 1.75 | 1.00 | 0.97 | 0.97 | |
| GLN2 | 1.99 | 2.30 | 2.57 | 2.26 | 2.70 | 2.86 | 1.16 | 1.29 | 1.12 | 1.20 | 1.27 | 1.06 | |
| GDH | GDH1 | 2.77 | 3.55 | 2.25 | 2.30 | 2.26 | 1.87 | 1.28 | 0.81 | 0.63 | 0.98 | 0.81 | 0.82 |
| GDH2 | 2.04 | 1.85 | 2.09 | 1.91 | 1.49 | 1.58 | 0.91 | 1.03 | 1.13 | 0.78 | 0.83 | 1.06 | |
| GOGAT | Os01g0681900 | 4.72 | 3.50 | 3.54 | 4.72 | 4.31 | 5.27 | 0.74 | 0.75 | 1.01 | 0.91 | 1.12 | 1.22 |
| Os05g0555600 | 0.48 | 0.37 | 0.50 | 0.51 | 0.46 | 0.57 | 0.78 | 1.03 | 1.32 | 0.91 | 1.12 | 1.23 | |
| GLSF | 9.99 | 8.89 | 7.99 | 9.39 | 9.79 | 10.61 | 0.89 | 0.80 | 0.90 | 1.04 | 1.13 | 1.08 | |
| NR | NIA1 | 0.23 | 0.20 | 0.20 | 0.29 | 0.33 | 0.30 | 0.87 | 0.87 | 1.00 | 1.13 | 1.01 | 0.90 |
| CB5-A | 0.36 | 0.37 | 0.41 | 0.35 | 0.34 | 0.38 | 1.04 | 1.16 | 1.11 | 0.97 | 1.09 | 1.13 | |
| NirA | Os01g0357100 | 2.56 | 1.44 | 1.63 | 2.70 | 3.74 | 3.16 | 0.56 | 0.63 | 1.13 | 1.38 | 1.17 | 0.84 |
| CA | CA1 | 10.38 | 10.78 | 10.25 | 12.77 | 14.23 | 11.23 | 1.04 | 0.99 | 0.95 | 1.11 | 0.88 | 0.79 |
| - | 1.52 | 1.90 | 1.69 | 1.41 | 1.28 | 1.18 | 1.25 | 1.11 | 0.89 | 0.91 | 0.83 | 0.92 | |
| - | 0.66 | 0.60 | 0.53 | 0.83 | 1.15 | 0.84 | 0.90 | 0.80 | 0.90 | 1.38 | 1.01 | 0.73 | |
| Transcriptome | TPM | Fold | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK_NX | CK_CS | CK_AUS | T_NX | T_CS | T_AUS | CK_CS/ CK_NX | CK_AUS/ CK_NX | CK_AUS/ CK_CS | T_CS/ T_NXz | T_AUS/ T_NX | T_AUS/ T_CS | ||
| GS | fluG | 7.36 | 7.09 | 8.07 | 11.06 | 8.63 | 6.87 | 0.96 | 1.10 | 1.14 | 0.78 | 0.62 | 0.80 |
| GLN1-1 | 460.01 | 647.77 | 849.01 | 440.98 | 494.79 | 972.39 | 1.41 | 1.85 | 1.31 | 1.12 | 2.21 | 1.97 | |
| GLN1-2 | 166.54 | 216.14 | 217.36 | 86.23 | 122.44 | 200.58 | 1.30 | 1.31 | 1.01 | 1.42 | 2.33 | 1.64 | |
| GLN1-3 | 1.24 | 0.10 | 12.72 | 27.75 | 199.31 | 27.53 | 0.08 | 10.26 | 127.20 | 7.18 | 0.99 | 0.14 | |
| GLN2 | 204.01 | 131.84 | 232.76 | 41.54 | 125.62 | 163.13 | 0.65 | 1.14 | 1.77 | 3.02 | 3.93 | 1.30 | |
| GDH | GDH1 | 31.20 | 55.95 | 49.60 | 20.53 | 35.14 | 29.92 | 1.79 | 1.59 | 0.89 | 1.71 | 1.46 | 0.85 |
| GDH2 | 33.42 | 40.07 | 57.72 | 95.67 | 103.85 | 68.10 | 1.20 | 1.73 | 1.44 | 1.09 | 0.71 | 0.66 | |
| GOGAT | Os01g0681900 | 68.35 | 32.71 | 70.93 | 75.89 | 116.38 | 85.39 | 0.48 | 1.04 | 2.17 | 3.64 | 5.08 | 1.40 |
| Os05g0555600 | 31.51 | 15.91 | 21.19 | 3.42 | 12.45 | 17.39 | 0.50 | 0.67 | 1.33 | 1.53 | 1.13 | 0.73 | |
| GLSF | 316.83 | 253.58 | 207.58 | 110.49 | 159.29 | 169.29 | 0.80 | 0.66 | 0.82 | 1.44 | 1.53 | 1.06 | |
| NR | NIA1 | 15.40 | 17.22 | 18.32 | 21.42 | 39.92 | 18.05 | 1.12 | 1.19 | 1.06 | 1.86 | 0.84 | 0.45 |
| CB5-A | 24.93 | 26.64 | 23.79 | 44.40 | 32.88 | 21.37 | 1.07 | 0.95 | 0.89 | 0.74 | 0.48 | 0.65 | |
| NirA | Os01g0357100 | 15.07 | 14.91 | 18.20 | 27.99 | 16.81 | 15.52 | 0.99 | 1.21 | 1.22 | 0.60 | 0.55 | 0.92 |
| CA | CA1 | 138.96 | 79.76 | 128.36 | 27.52 | 74.26 | 67.40 | 0.57 | 0.92 | 1.61 | 2.70 | 2.45 | 0.91 |
| - | 249.36 | 204.38 | 150.68 | 112.35 | 159.95 | 139.68 | 0.82 | 0.60 | 0.74 | 1.42 | 1.24 | 0.87 | |
| - | 4.22 | 5.02 | 12.40 | 0.73 | 8.37 | 13.92 | 1.19 | 2.94 | 2.47 | 11.47 | 19.07 | 1.66 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Q.; Fu, H.; Hu, F.; Li, S.; Chen, Q.; Peng, S.; Yang, C.; Liu, Y.; Chen, Y. Effects of Biological Nitrogen Metabolism on Glufosinate-Susceptible and -Resistant Goosegrass (Eleusine indica L.). Int. J. Mol. Sci. 2023, 24, 13791. https://doi.org/10.3390/ijms241813791
Luo Q, Fu H, Hu F, Li S, Chen Q, Peng S, Yang C, Liu Y, Chen Y. Effects of Biological Nitrogen Metabolism on Glufosinate-Susceptible and -Resistant Goosegrass (Eleusine indica L.). International Journal of Molecular Sciences. 2023; 24(18):13791. https://doi.org/10.3390/ijms241813791
Chicago/Turabian StyleLuo, Qiyu, Hao Fu, Fang Hu, Shiguo Li, Qiqi Chen, Shangming Peng, Cunyi Yang, Yaoguang Liu, and Yong Chen. 2023. "Effects of Biological Nitrogen Metabolism on Glufosinate-Susceptible and -Resistant Goosegrass (Eleusine indica L.)" International Journal of Molecular Sciences 24, no. 18: 13791. https://doi.org/10.3390/ijms241813791
APA StyleLuo, Q., Fu, H., Hu, F., Li, S., Chen, Q., Peng, S., Yang, C., Liu, Y., & Chen, Y. (2023). Effects of Biological Nitrogen Metabolism on Glufosinate-Susceptible and -Resistant Goosegrass (Eleusine indica L.). International Journal of Molecular Sciences, 24(18), 13791. https://doi.org/10.3390/ijms241813791