
Cellular Immunotherapy in Mice Prevents Maternal Hypertension and Restores Anti-Inflammatory Cytokine Balance in Maternal and Fetal Tissues


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
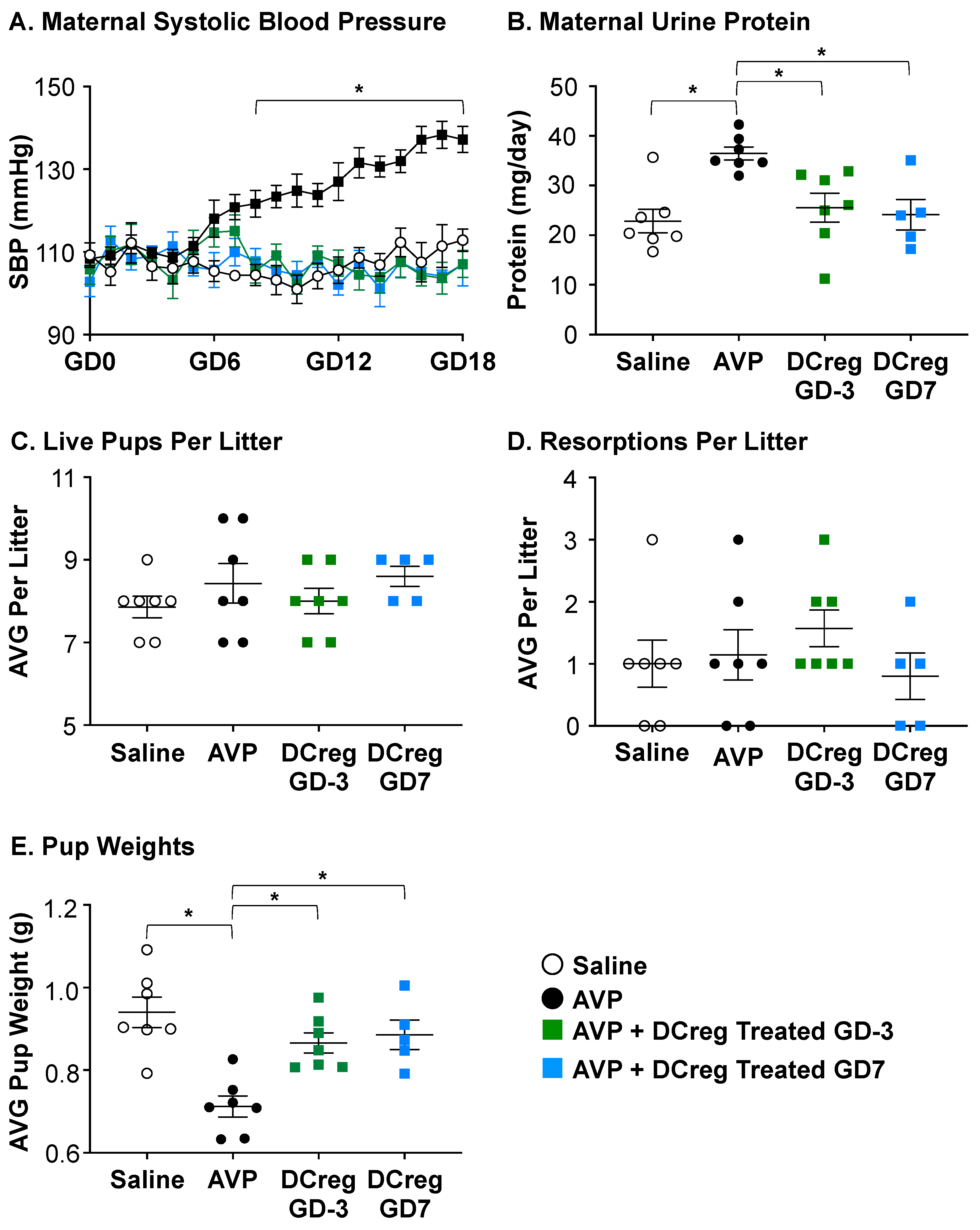
2.1. DCreg Administration Prevents Maternal and Fetal PreE-Associated Morbidities
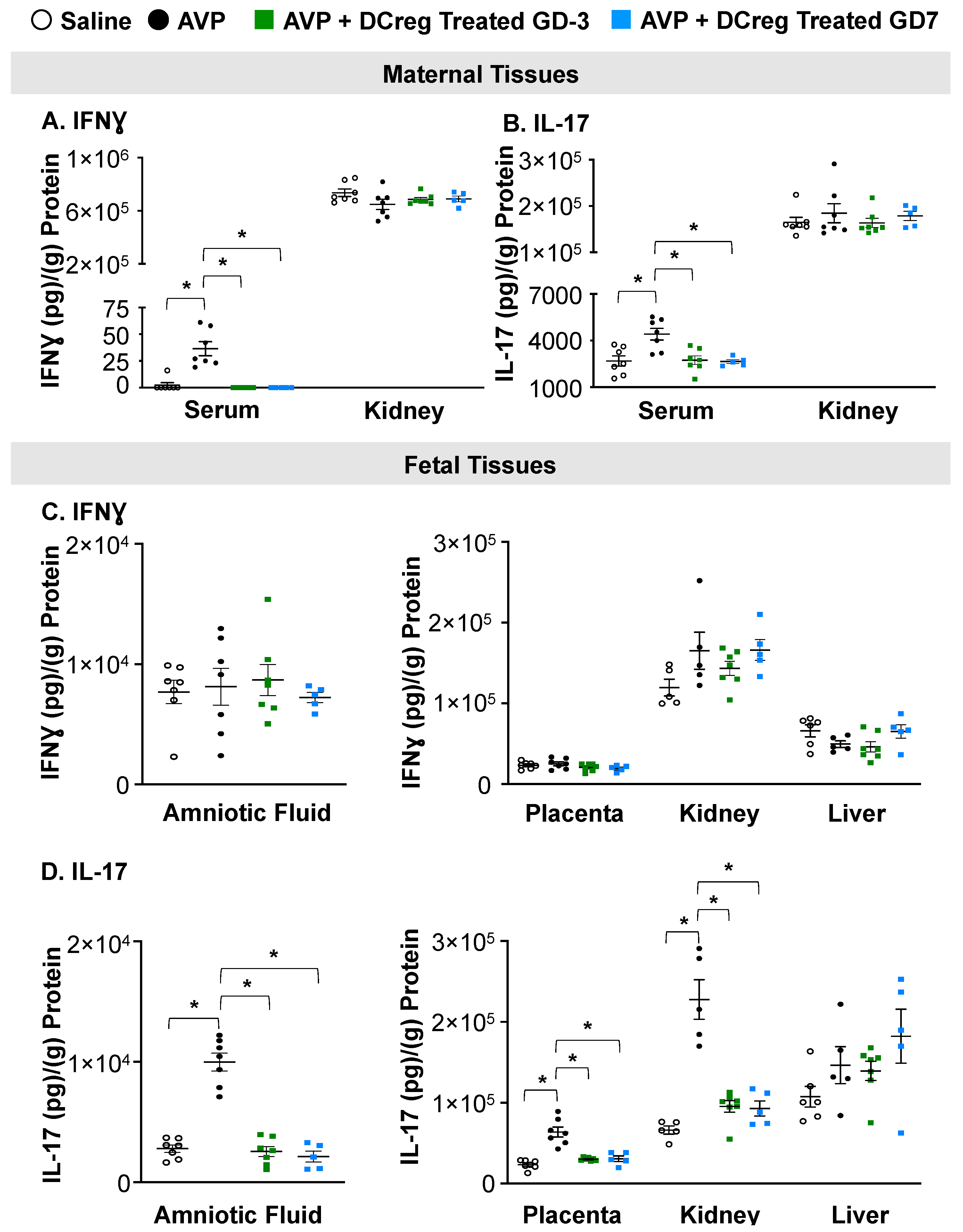
2.2. DCreg Treatment Prevents Elevated Maternal and Fetal Inflammatory Cytokines
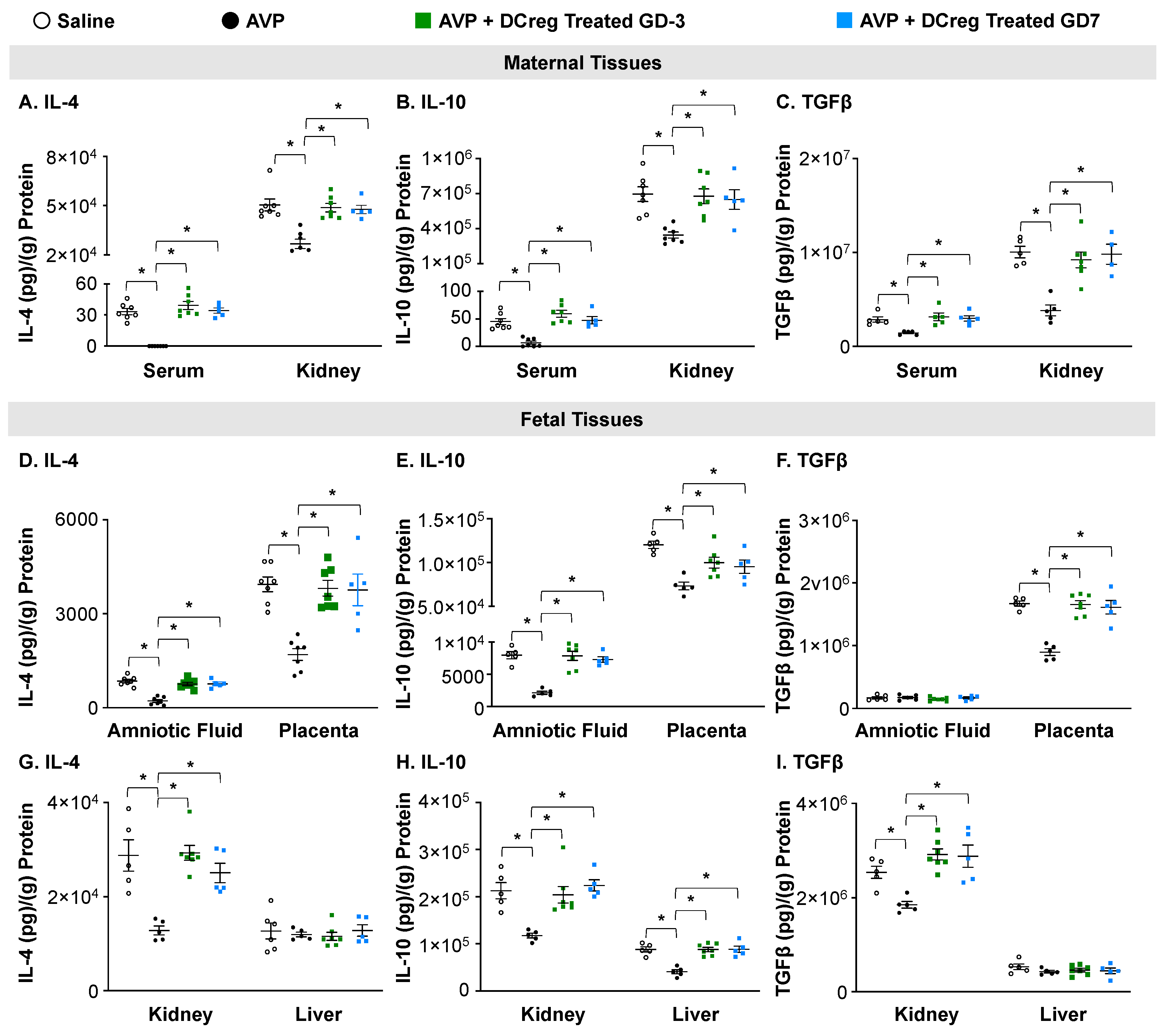
2.3. Treatment with DCreg Restores Maternal and Fetal Tissue Anti-Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Generation of DCreg
4.2. Induction of Preeclampsia
4.3. Whole Tissue Extraction of Protein and Total Protein Analysis
4.4. Blood Pressure and Proteinuria
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villar, J.; Carroli, G.; Wojdyla, D.; Abalos, E.; Giordano, D.; Ba’aqeel, H.; Farnot, U.; Bergsjo, P.; Bakketeig, L.; Lumbiganon, P.; et al. Preeclampsia, gestational hypertension and intrauterine growth restriction, related or independent conditions? Am. J. Obstet. Gynecol. 2006, 194, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obs. Gynecol. 2020, 135, e237–e260. [CrossRef]
- Butwick, A.J.; Druzin, M.L.; Shaw, G.M.; Guo, N. Evaluation of US State-Level Variation in Hypertensive Disorders of Pregnancy. JAMA Netw. Open. 2020, 3, e2018741. [Google Scholar] [CrossRef]
- Kajantie, E.; Eriksson, J.G.; Osmond, C.; Thornburg, K.; Barker, D.J. Pre-eclampsia is associated with increased risk of stroke in the adult offspring: The Helsinki birth cohort study. Stroke A J. Cereb. Circ. 2009, 40, 1176–1180. [Google Scholar] [CrossRef] [PubMed]
- Lykke, J.A.; Langhoff-Roos, J.; Sibai, B.M.; Funai, E.F.; Triche, E.W.; Paidas, M.J. Hypertensive pregnancy disorders and subsequent cardiovascular morbidity and type 2 diabetes mellitus in the mother. Hypertension 2009, 53, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Nohr, E.A.; Bech, B.H.; Vestergaard, M.; Catov, J.M.; Olsen, J. Health of children born to mothers who had preeclampsia: A population-based cohort study. Am. J. Obstet. Gynecol. 2009, 201, 269.e1–269.e10. [Google Scholar] [CrossRef]
- Magnussen, E.B.; Vatten, L.J.; Smith, G.D.; Romundstad, P.R. Hypertensive disorders in pregnancy and subsequently measured cardiovascular risk factors. Obs. Gynecol. 2009, 114, 961–970. [Google Scholar] [CrossRef]
- Kuklina, E.V.; Ayala, C.; Callaghan, W.M. Hypertensive disorders and severe obstetric morbidity in the United States. Obs. Gynecol. 2009, 113, 1299–1306. [Google Scholar] [CrossRef]
- Baecke, M.; Spaanderman, M.E.; van der Werf, S.P. Cognitive function after pre-eclampsia: An explorative study. J. Psychosom. Obs. Gynaecol. 2009, 30, 58–64. [Google Scholar] [CrossRef]
- Brusse, I.; Duvekot, J.; Jongerling, J.; Steegers, E.; De Koning, I. Impaired maternal cognitive functioning after pregnancies complicated by severe pre-eclampsia: A pilot case-control study. Acta Obstet. Et. Gynecol. Scand. 2008, 87, 408–412. [Google Scholar] [CrossRef]
- Fields, J.A.; Garovic, V.D.; Mielke, M.M.; Kantarci, K.; Jayachandran, M.; White, W.M.; Butts, A.M.; Graff-Radford, J.; Lahr, B.D.; Bailey, K.R.; et al. Preeclampsia and cognitive impairment later in life. Am. J. Obstet. Gynecol. 2017, 217, 74.e1–74.e11. [Google Scholar] [CrossRef]
- Postma, I.R.; Bouma, A.; Ankersmit, I.F.; Zeeman, G.G. Neurocognitive functioning following preeclampsia and eclampsia: A long-term follow-up study. Am. J. Obstet. Gynecol. 2014, 211, 37.e1–37.e9. [Google Scholar] [CrossRef]
- Postma, I.R.; Groen, H.; Easterling, T.R.; Tsigas, E.Z.; Wilson, M.L.; Porcel, J.; Zeeman, G.G. The brain study: Cognition, quality of life and social functioning following preeclampsia; An observational study. Pregnancy Hypertens. 2013, 3, 227–234. [Google Scholar] [CrossRef]
- Postma, I.R.; Wessel, I.; Aarnoudse, J.G.; Zeeman, G.G. Neurocognitive functioning in women with a history of eclampsia: Executive functioning and sustained attention. Am. J. Perinatol. 2010, 27, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Roes, E.M.; Raijmakers, M.T.; Schoonenberg, M.; Wanner, N.; Peters, W.H.; Steegers, E.A. Physical well-being in women with a history of severe preeclampsia. J. Matern. Fetal Neonatal Med. 2005, 18, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.C.; Tremble, S.M.; Cipolla, M.J. Experimental Preeclampsia Causes Long-Lasting Hippocampal Vascular Dysfunction and Memory Impairment. Front. Physiol. 2022, 13, 889918. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wei, K.; Lee, P.M.Y.; Qin, G.; Yu, Y.; Li, J. Maternal hypertensive disorder of pregnancy and mortality in offspring from birth to young adulthood: National population based cohort study. BMJ 2022, 379, e072157. [Google Scholar] [CrossRef]
- Korzeniewski, S.J.; Sutton, E.; Escudero, C.; Roberts, J.M. The Global Pregnancy Collaboration (CoLab) symposium on short- and long-term outcomes in offspring whose mothers had preeclampsia: A scoping review of clinical evidence. Front. Med. 2022, 9, 984291. [Google Scholar] [CrossRef]
- Santillan, M.K.; Santillan, D.A.; Sigmund, C.D.; Hunter, S.K. From molecules to medicine: A future cure for preeclampsia? Drug. News Perspect. 2009, 22, 531–541. [Google Scholar] [CrossRef]
- Kadyrov, M.; Kingdom, J.C.; Huppertz, B. Divergent trophoblast invasion and apoptosis in placental bed spiral arteries from pregnancies complicated by maternal anemia and early-onset preeclampsia/intrauterine growth restriction. Am. J. Obstet. Gynecol. 2006, 194, 557–563. [Google Scholar] [CrossRef]
- Darmochwal-Kolarz, D.; Kludka-Sternik, M.; Tabarkiewicz, J.; Kolarz, B.; Rolinski, J.; Leszczynska-Gorzelak, B.; Oleszczuk, J. The predominance of Th17 lymphocytes and decreased number and function of Treg cells in preeclampsia. J. Reprod. Immunol. 2012, 93, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Wallace, K.; Richards, S.; Dhillon, P.; Weimer, A.; Edholm, E.S.; Bengten, E.; Wilson, M.; Martin, J.N., Jr.; LaMarca, B. CD4+ T-helper cells stimulated in response to placental ischemia mediate hypertension during pregnancy. Hypertension 2011, 57, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Laresgoiti-Servitje, E.; Gómez-López, N.; Olson, D.M. An immunological insight into the origins of pre-eclampsia. Hum. Reprod. Update 2010, 16, 510–524. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L. Immunology of pre-eclampsia. Am. J. Reprod. Immunol. 2010, 63, 534–543. [Google Scholar] [CrossRef]
- Scroggins, S.M.; Santillan, D.A.; Lund, J.M.; Sandgren, J.A.; Krotz, L.K.; Hamilton, W.S.; Devor, E.J.; Davis, H.A.; Pierce, G.L.; Gibson-Corley, K.N.; et al. Elevated vasopressin in pregnant mice induces T-helper subset alterations consistent with human preeclampsia. Clin. Sci. 2018, 132, 419–436. [Google Scholar] [CrossRef]
- Coombes, J.L.; Powrie, F. Dendritic cells in intestinal immune regulation. Nat. Rev. Immunol. 2008, 8, 435–446. [Google Scholar] [CrossRef]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef]
- Joffre, O.P.; Segura, E.; Savina, A.; Amigorena, S. Cross-presentation by dendritic cells. Nat. Rev. Immunol. 2012, 12, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Morelli, A.E.; Thomson, A.W. Tolerogenic dendritic cells and the quest for transplant tolerance. Nat. Rev. Immunol. 2007, 7, 610–621. [Google Scholar] [CrossRef]
- Rutella, S.; Lemoli, R.M. Regulatory T cells and tolerogenic dendritic cells: From basic biology to clinical applications. Immunol. Lett. 2004, 94, 11–26. [Google Scholar] [CrossRef]
- Steinman, R.M.; Hawiger, D.; Nussenzweig, M.C. Tolerogenic dendritic cells. Annu. Rev. Immunol. 2003, 21, 685–711. [Google Scholar] [CrossRef] [PubMed]
- Stenger, E.O.; Turnquist, H.R.; Mapara, M.Y.; Thomson, A.W. Dendritic cells and regulation of graft-versus-host disease and graft-versus-leukemia activity. Blood 2012, 119, 5088–5103. [Google Scholar] [CrossRef] [PubMed]
- Cools, N.; Ponsaerts, P.; Van Tendeloo, V.F.; Berneman, Z.N. Balancing between immunity and tolerance: An interplay between dendritic cells, regulatory T cells, and effector T cells. J. Leukoc. Biol. 2007, 82, 1365–1374. [Google Scholar] [CrossRef]
- Torres-Aguilar, H.; Aguilar-Ruiz, S.R.; Gonzalez-Perez, G.; Munguia, R.; Bajana, S.; Meraz-Rios, M.A.; Sanchez-Torres, C. Tolerogenic dendritic cells generated with different immunosuppressive cytokines induce antigen-specific anergy and regulatory properties in memory CD4+ T Cells. J. Immunol. 2010, 184, 1765–1775. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, S.; Hong, L.; Diao, L.; Cai, S.; Yin, T.; Zeng, Y. Characterization of progesterone-induced dendritic cells in metabolic and immunologic reprogramming. J. Reprod. Immunol. 2023, 159, 104128. [Google Scholar] [CrossRef]
- Marin, E.; Bouchet-Delbos, L.; Renoult, O.; Louvet, C.; Nerriere-Daguin, V.; Managh, A.J.; Even, A.; Giraud, M.; Vu Manh, T.P.; Aguesse, A.; et al. Human Tolerogenic Dendritic Cells Regulate Immune Responses through Lactate Synthesis. Cell. Metab. 2019, 30, 1075–1090.e8. [Google Scholar] [CrossRef]
- Navarro-Barriuso, J.; Mansilla, M.J.; Quirant-Sanchez, B.; Teniente-Serra, A.; Ramo-Tello, C.; Martinez-Caceres, E.M. Vitamin D3-Induced Tolerogenic Dendritic Cells Modulate the Transcriptomic Profile of T CD4(+) Cells towards a Functional Hyporesponsiveness. Front. Immunol. 2020, 11, 599623. [Google Scholar] [CrossRef]
- Fujita, S.; Seino, K.; Sato, K.; Sato, Y.; Eizumi, K.; Yamashita, N.; Taniguchi, M. Regulatory dendritic cells act as regulators of acute lethal systemic inflammatory response. Blood 2006, 107, 3656–3664. [Google Scholar] [CrossRef]
- Fujita, S.; Yamashita, N.; Ishii, Y.; Sato, Y.; Sato, K.; Eizumi, K.; Fukaya, T.; Nozawa, R.; Takamoto, Y.; Taniguchi, M. Regulatory dendritic cells protect against allergic airway inflammation in a murine asthmatic model. J. Allergy Clin. Immunol. 2008, 121, 95–104. [Google Scholar] [CrossRef]
- Lan, Y.Y.; Wang, Z.; Raimondi, G.; Wu, W.; Colvin, B.L.; de Creus, A.; Thomson, A.W. “Alternatively activated” dendritic cells preferentially secrete IL-10, expand Foxp3+CD4+ T cells, and induce long-term organ allograft survival in combination with CTLA4-Ig. J. Immunol. 2006, 177, 5868–5877. [Google Scholar] [CrossRef] [PubMed]
- Chorny, A.; Gonzalez-Rey, E.; Fernandez-Martin, A.; Ganea, D.; Delgado, M. Vasoactive intestinal peptide induces regulatory dendritic cells that prevent acute graft-versus-host disease while maintaining the graft-versus-tumor response. Blood 2006, 107, 3787–3794. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yamashita, N.; Baba, M.; Matsuyama, T. Regulatory dendritic cells protect mice from murine acute graft-versus-host disease and leukemia relapse. Immunity 2003, 18, 367–379. [Google Scholar] [CrossRef]
- Scroggins, S.M.; Olivier, A.K.; Meyerholz, D.K.; Schlueter, A.J. Characterization of regulatory dendritic cells that mitigate acute graft-versus-host disease in older mice following allogeneic bone marrow transplantation. PLoS ONE 2013, 8, e75158. [Google Scholar] [CrossRef]
- Wan, X.; Bao, L.; Ma, G.; Long, T.; Li, H.; Zhang, Y.; Jiang, H. Tolerogenic dendritic cells alleviate collagen-induced arthritis by forming microchimerism and affecting the expression of immune checkpoint molecules. Eur. J. Immunol. 2022, 52, 1980–1992. [Google Scholar] [CrossRef]
- Chorny, A.; Gonzalez-Rey, E.; Fernandez-Martin, A.; Pozo, D.; Ganea, D.; Delgado, M. Vasoactive intestinal peptide induces regulatory dendritic cells with therapeutic effects on autoimmune disorders. Proc. Natl. Acad. Sci. USA 2005, 102, 13562–13567. [Google Scholar] [CrossRef] [PubMed]
- Sandgren, J.A.; Deng, G.; Linggonegoro, D.W.; Scroggins, S.M.; Perschbacher, K.J.; Nair, A.R.; Nishimura, T.E.; Zhang, S.Y.; Agbor, L.N.; Wu, J.; et al. Arginine vasopressin infusion is sufficient to model clinical features of preeclampsia in mice. JCI Insight 2018, 3, e99403. [Google Scholar] [CrossRef] [PubMed]
- Santillan, M.K.; Santillan, D.A.; Scroggins, S.M.; Min, J.Y.; Sandgren, J.A.; Pearson, N.A.; Leslie, K.K.; Hunter, S.K.; Zamba, G.K.; Gibson-Corley, K.N.; et al. Vasopressin in preeclampsia: A novel very early human pregnancy biomarker and clinically relevant mouse model. Hypertension 2014, 64, 852–859. [Google Scholar] [CrossRef]
- Ramdin, S.; Naicker, T.; Pillay, V.; Singh, S.D.; Baijnath, S.; Mkhwanazi, B.N.; Govender, N. Physiological characterization of an arginine vasopressin rat model of preeclampsia. Syst. Biol. Reprod. Med. 2022, 68, 55–69. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Tian, M.; Hu, X.; Wang, L.; Ji, J.; Liao, A. The altered PD-1/PD-L1 pathway delivers the ‘one-two punch’ effects to promote the Treg/Th17 imbalance in pre-eclampsia. Cell. Mol. Immunol. 2018, 15, 710–723. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Lian, D.; Zheng, X.; Zhang, Z.X.; Ichim, T.E.; Xia, X.; Huang, X.; Vladau, C.; Suzuki, M.; et al. Generation of therapeutic dendritic cells and regulatory T cells for preventing allogeneic cardiac graft rejection. Clin. Immunol. 2008, 127, 313–321. [Google Scholar] [CrossRef]
- Littlejohn, N.K.; Siel, R.B., Jr.; Ketsawatsomkron, P.; Pelham, C.J.; Pearson, N.A.; Hilzendeger, A.M.; Buehrer, B.A.; Weidemann, B.J.; Li, H.; Davis, D.R.; et al. Hypertension in mice with transgenic activation of the brain renin-angiotensin system is vasopressin dependent. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R818–R828. [Google Scholar] [CrossRef] [PubMed]
- Santillan, M.K.; Pelham, C.J.; Ketsawatsomkron, P.; Santillan, D.A.; Davis, D.R.; Devor, E.J.; Gibson-Corley, K.N.; Scroggins, S.M.; Grobe, J.L.; Yang, B.; et al. Pregnant mice lacking indoleamine 2,3-dioxygenase exhibit preeclampsia phenotypes. Physiol. Rep. 2015, 3, e1225. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gray, G.; Scroggins, D.G.; Wilson, K.T.; Scroggins, S.M. Cellular Immunotherapy in Mice Prevents Maternal Hypertension and Restores Anti-Inflammatory Cytokine Balance in Maternal and Fetal Tissues. Int. J. Mol. Sci. 2023, 24, 13594. https://doi.org/10.3390/ijms241713594
Gray G, Scroggins DG, Wilson KT, Scroggins SM. Cellular Immunotherapy in Mice Prevents Maternal Hypertension and Restores Anti-Inflammatory Cytokine Balance in Maternal and Fetal Tissues. International Journal of Molecular Sciences. 2023; 24(17):13594. https://doi.org/10.3390/ijms241713594
Chicago/Turabian StyleGray, Gabrielle, Douglas G. Scroggins, Katlin T. Wilson, and Sabrina M. Scroggins. 2023. "Cellular Immunotherapy in Mice Prevents Maternal Hypertension and Restores Anti-Inflammatory Cytokine Balance in Maternal and Fetal Tissues" International Journal of Molecular Sciences 24, no. 17: 13594. https://doi.org/10.3390/ijms241713594
APA StyleGray, G., Scroggins, D. G., Wilson, K. T., & Scroggins, S. M. (2023). Cellular Immunotherapy in Mice Prevents Maternal Hypertension and Restores Anti-Inflammatory Cytokine Balance in Maternal and Fetal Tissues. International Journal of Molecular Sciences, 24(17), 13594. https://doi.org/10.3390/ijms241713594