
ZmHMA3, a Member of the Heavy-Metal-Transporting ATPase Family, Regulates Cd and Zn Tolerance in Maize

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Phylogenetic Tree and the Conserved Structural Domain Analysis of Maize HMA Family Genes
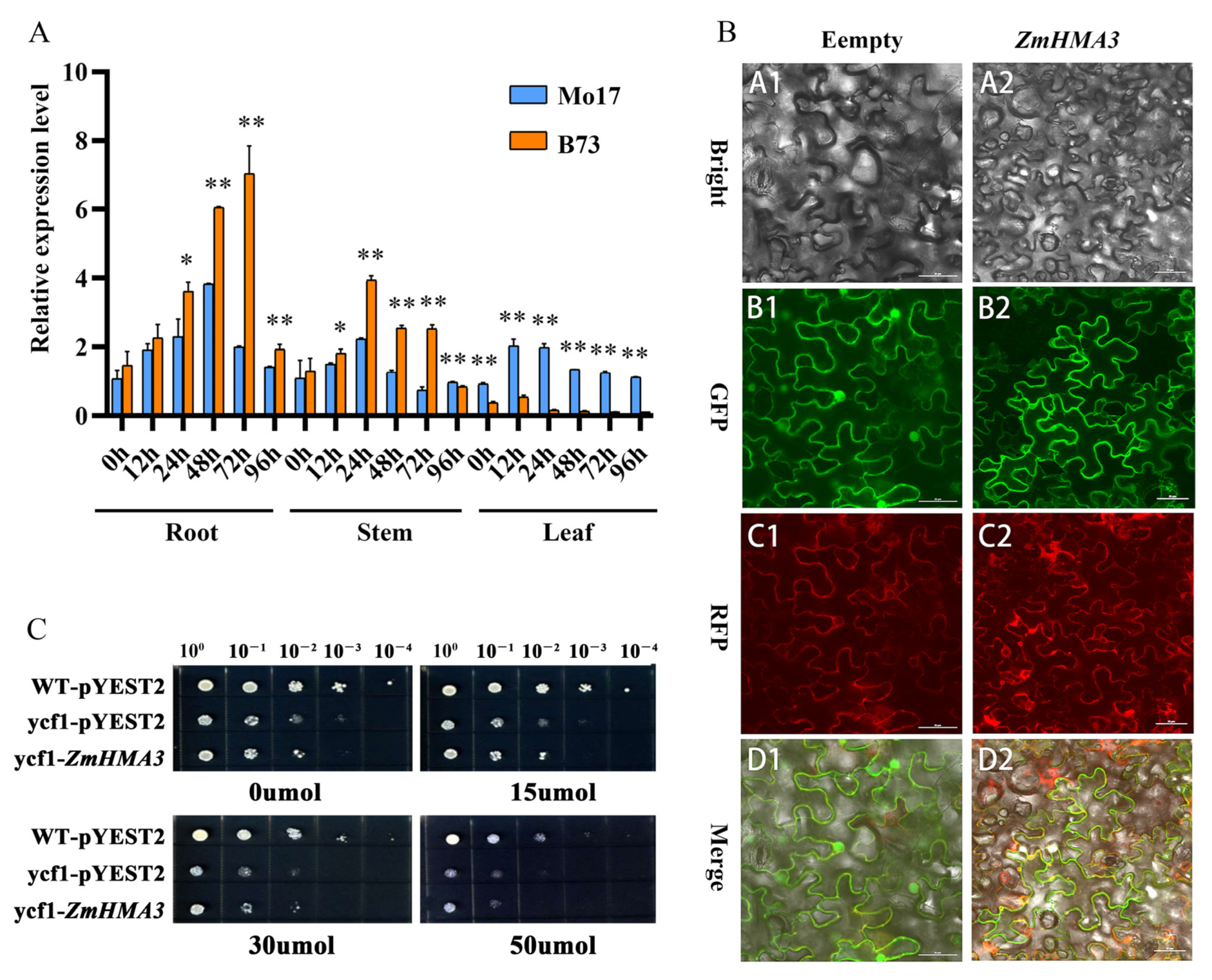
2.2. Functional Expression of ZmHMA3 in Response to Cd Stress
2.3. Subcellular Localization of ZmHMA3 and Cd Sensibility in Yeast
2.4. Mutant of AtHMA2 Decreased Resistance to Cd Stress in Arabidopsis
2.5. Overexpression of ZmHMA3 Increased Tolerance in Heavy Metal Stress
2.6. The Effects of Heavy Metal Stress on Maize: Root System Responses, Cd and Zn Accumulation and Translocation, and Subcellular Cd and Zn Distribution
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. ZmHMA3 Expression Analysis Using Real-Time Quantitative PCR
4.3. Subcellular Localization Assay
4.4. Yeast Heterologous Expression
4.5. Genetic Transformation of Arabidopsis Thaliana
4.6. Overexpression of ZmHMA3 in Maize
4.7. Treatment of Cd and Zn Stress in Maize Seedlings
4.8. Determination of Agronomic Traits and Physiological Indicators in Maize Seedlings
4.9. Uptake and Accumulation of Cd and Zn in Maize Seedlings
- (1)
- Determination of Cd or Zn content
- (2)
- Determination of different Cd and Zn concentrations in the subcellular analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, R.; Zhang, Z.; Chen, X.; Gu, C.; Zhang, Q. The latest research progress on the stress responses of bZIP involved in plants. Biotechnol. Bull. 2019, 35, 143–155. (In Chinese) [Google Scholar]
- Passow, H.; Rothstein, A.; Clarkson, T. The general pharmacology of the heavy metals. Pharmacol. Revs. 1961, 13, 185–224. [Google Scholar]
- Antonovics, J.; Bradshaw, A.D.; Turner, R. Heavy metal tolerance in plants. Adv. Ecol. Res. 1971, 7, 1–85. [Google Scholar]
- Xiao, J.; Yuan, Y. Remediation and management of heavy metal contaminated arable land in ChangZhuTan: Exploration, dilemma and breakthrough. Jiangxi Soc. Sci. 2019, 39, 73–81. (In Chinese) [Google Scholar]
- Hasan, S.A.; Fariduddin, Q.; Ali, B.; Hayat, S.; Ahmad, A. Cadmium: Toxicity and tolerance in plants. J. Environ. Biol. 2009, 30, 165–174. [Google Scholar] [PubMed]
- Li, B.; Wang, C.; Tan, T.; Li, H.; Yang, J.; Li, Q.; Yuan, Q. Regional distribution and pollution evaluation of heavy metal pollution in yopsoils of the chengdu plain. J. Nucl. Agric. 2009, 23, 308–315. (In Chinese) [Google Scholar]
- Sun, Q.; Wang, X.; Din, S. Rhizosphere effects in metal absorption by hyperaccumulators and its research advances. J. Ecol. 2005, 24, 30–36. (In Chinese) [Google Scholar]
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef]
- Xu, W.; Lu, G.; Dang, Z.; Liao, C.; Chen, Q.; Yi, X. Uptake and distribution of Cd in sweet maize grown on contaminated soils: A field-scale study. Bioinorg. Chem. Appl. 2013, 2013, 959764. [Google Scholar] [CrossRef]
- Assem, S.K. Maize, tropical (Zea mays L.). In Agrobacterium Protocols; Springer: New York, NY, USA, 2015; Volume 1, pp. 119–134. [Google Scholar]
- Aslam, Z.; Khattak, J.Z.K.; Ahmed, M. Drought tolerance in cereal grain crops under changing climate. In Quantification of Climate Variability, Adaptation and Mitigation for Agricultural Sustainability; Springer International Publishing: Cham, Switzerland, 2017; pp. 181–209. [Google Scholar]
- Usmani, M.M.; Nawaz, F.; Majeed, S.; Shehzad, M.A.; Ahmad, K.S.; Akhtar, G.; Aqib, M.; Shabbir, R.N. Sulfate-mediated drought tolerance in maize involves regulation at physiological and biochemical levels. Sci. Rep. 2020, 10, 1147. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B. Global maize production, consumption and trade: Trends and R&D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar]
- Yang, Y.; Nan, Z.; Zhao, Z. Bioaccumulation and translocation of cadmium in wheat (Triticum aestivum L.) and maize (Zea mays L.) from the polluted oasis soil of Northwestern China. Chem. Speciat. Bioavailab. 2014, 26, 43–51. [Google Scholar] [CrossRef]
- Castillo-Michel, H.A.; Hernandez, N.; Martinez-Martinez, A.; Parsons, J.G.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Coordination and speciation of cadmium in corn seedlings and its effects on macro-and micronutrients uptake. Plant Physiol. Biochem. 2009, 47, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Xiao, J.; Li, J.; Du, Q.; Zhu, L.; Wang, H.; Zhu, R.; Zhao, H. Accumulation and transport characteristics of Cd, Pb, Zn and As in different maize varieties. J. Environ. Sci. 2022, 43, 4232–4252. (In Chinese) [Google Scholar]
- Han, J.; Li, E. Detection of influencing factors of heavy metals in maize and analysis of pollution characteristics based on geographical detector. J. Arid. Land Resour. Environ. 2023, 37, 113–120. (In Chinese) [Google Scholar]
- Li, D.; Xu, X.; Hu, X.; Liu, Q.; Wang, Z.; Zhang, H.; Wang, H.; Wei, M.; Wang, H.; Liu, H.; et al. Genome-wide analysis and heavy metal-induced expression profiling of the HMA gene family in Populus trichocarpa. Front. Plant Sci. 2015, 6, 1149. [Google Scholar] [CrossRef]
- Williams, L.E.; Mills, R.F. P1B-ATPases-an ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef]
- Baxter, I.; Tchieu, J.; Sussman, M.R.; Boutry, M.; Palmgren, M.G.; Gribskov, M.; Harper, J.F.; Axelsen, K.B. Genomic comparison of P-Type ATPase ion pumps in Arabidopsis and rice. Plant Physiol. 2003, 2, 618–628. [Google Scholar] [CrossRef]
- Cobbett, C.S.; Hussain, D.; Haydon, M.J. Structural and functional relationships between type 1B heavy meta-transporting P-type ATPases in Arabidopsis. New Phytol. 2003, 2, 315–321. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Kono, I.; Yokosho, K.; Ando, T.; Yano, M.; Ma, J.F. A major quantitative trait locus controlling cadmium translocation in rice (Oryza sativa). New Phytol. 2009, 182, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Deng, F.; Yamaji, N.; Pinson, S.; Kashino, M.; Danku, J.; Douglas, A.; Guerinot, M.; Salt, D.; Ma, J. A heavy metal P-type ATPase OsHMA4 prevents copper accumulation in rice grain. Nat. Commun. 2016, 7, 12138. [Google Scholar] [CrossRef]
- Kulich, I.; Vojtíková, Z.; Glanc, M.; Ortmannová, J.; Rasmann, S.; Žárský, V. Cell wall maturation of Arabidopsis trichomes is dependent on exocyst subunit EXO70H4 and involves callose deposition. Plant Physiol. 2015, 168, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Horváth, E.; Janda, T.; Páldi, E.; Szalai, G. Physiological changes and defense mechanisms induced by cadmium stress in maize. J. Plant Nutr. Soil Sci. 2006, 169, 239–246. [Google Scholar] [CrossRef]
- Khaliq, A.; Ali, S.; Hameed, A.; Farooq, M.A.; Farid, M.; Shakoor, M.B.; Mahmood, K.; Ishaque, W.; Rizwan, M. Silicon alleviates nickel toxicity in cotton seedlings through enhancing growth, photosynthesis, and suppressing Ni uptake and oxidative stress. Arch. Agron. Soil Sci. 2016, 62, 633–647. [Google Scholar] [CrossRef]
- Verret, F.; Gravot, A.; Auroy, P.; Preveral, S.; Forestier, C.; Vavasseur, A.; Richaud, P. Heavy metal transport by AtHMA4 involves the N-terminal degenerated metal binding domain and the C-terminal His11 stretch. FEBS Lett. 2005, 579, 1515–1522. [Google Scholar] [CrossRef]
- Eren, E.; Arguello, J.M. Arabidopsis HMA2, a divalent heavy metal-transporting PIB-type ATPase, is involved in cytoplasmic Zn2+ homeostasis. Plant Physiol. 2004, 136, 3712–3723. [Google Scholar] [CrossRef]
- Bækgaard, L.; Mikkelsen, M.D.; Sørensen, D.M.; Hegelund, J.N.; Persson, D.P.; Mills, R.F.; Yang, Z.; Husted, S.; Andersen, J.P.; Buch-Pedersen, M.J. A combined zinc/cadmium sensor and zinc/cadmium export regulator in a heavy metal pump. J. Biol. Chem. 2010, 285, 31243–31252. [Google Scholar] [CrossRef]
- Guo, J.; Long, L.; Chen, A.; Dong, X.; Liu, Z.; Chen, L.; Wang, J.; Yuan, L. Tonoplast-localized transporter ZmNRAMP2 confers root-to-shoot translocation of manganese in maize. Plant Physiol. 2022, 190, 2601–2616. [Google Scholar] [CrossRef] [PubMed]
- Pace, J.; Lee, N.; Naik, H.S.; Ganapathysubramanian, B.; Lübberstedt, T. Analysis of maize (Zea mays L.) seedling roots with the high-throughput image analysis tool ARIA (Automatic Root Image Analysis). PLoS ONE 2014, 9, e108255. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Sun, L.; Tan, L. Progress in ZIP transporter gene family in rice. Hereditas 2018, 40, 33–43. (In Chinese) [Google Scholar] [PubMed]
- Sinclair, S.A.; Sherson, S.M.; Jarvis, R.; Camakaris, J.; Cobbett, C.S. The use of the zinc-fluorophore, Zinpyr-1, in the study of zinc homeostasis in Arabidopsis roots. New Phytol. 2007, 174, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Pei, K. Effect of Cd and Zn Stress on Seed Germination, Physiology and Biochemistry of Nelumbo nucifera. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2010. (In Chinese). [Google Scholar]
- Xu, Z.-M.; Mei, X.-Q.; Tan, L.; Li, Q.-S.; Wang, L.-L.; He, B.-Y.; Guo, S.-H.; Zhou, C.; Ye, H.-J. Low root/shoot (R/S) biomass ratio can be an indicator of low cadmium accumulation in the shoot of Chinese flowering cabbage (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee) cultivars. Environ. Sci. Pollut. Res. 2018, 25, 36328–36340. [Google Scholar] [CrossRef]
- Du, C.; Zhang, N.; Lei, B.; Hu, W.; Fu, B.; Chen, A.; Mao, Y.; Mu, L.; Wang, H.; Yan, T.; et al. Differences of cadmium and zinc accumulation and translocation in different varieties of Zea mays. J. Agro-Environ. Sci. 2017, 36, 16–23. (In Chinese) [Google Scholar]
- Ma, G.; Yao, Y. Effect of zinc on the growth of maize under cadmium stress. J. Green Sci. Technol. 2013, 166–167. (In Chinese) [Google Scholar]
- Dresler, S.; Hanaka, A.; Bednarek, W.; Maksymiec, W. Accumulation of low-molecular-weight organic acids in roots and leaf segments of Zea mays plants treated with cadmium and copper. Acta Physiol. Plant. 2014, 36, 1565–1575. [Google Scholar] [CrossRef]
- Weigel, H.J.; Jäger, H.J. Subcellular distribution and chemical form of cadmium in bean plants. Plant Physiol. 1980, 65, 480–482. [Google Scholar] [CrossRef]
- Tang, B.; Luo, M.; Zhang, Y.; Guo, H.; Li, J.; Song, W.; Zhang, R.; Feng, Z.; Kong, M.; Li, H. Natural variations in the P-type ATPase heavy metal transporter gene ZmHMA3 control cadmium accumulation in maize grains. J. Exp. Bot. 2021, 72, 6230–6246. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.; Li, Y.; Wu, X.; Wu, W.; Zhang, Y.; Zhan, P.; Meng, X.; Hu, G.; Yang, S.; Lin, H. ZmHMA3, a Member of the Heavy-Metal-Transporting ATPase Family, Regulates Cd and Zn Tolerance in Maize. Int. J. Mol. Sci. 2023, 24, 13496. https://doi.org/10.3390/ijms241713496
Liao C, Li Y, Wu X, Wu W, Zhang Y, Zhan P, Meng X, Hu G, Yang S, Lin H. ZmHMA3, a Member of the Heavy-Metal-Transporting ATPase Family, Regulates Cd and Zn Tolerance in Maize. International Journal of Molecular Sciences. 2023; 24(17):13496. https://doi.org/10.3390/ijms241713496
Chicago/Turabian StyleLiao, Changjian, Youqiang Li, Xiaohong Wu, Wenmei Wu, Yang Zhang, Penglin Zhan, Xin Meng, Gaojiao Hu, Shiqi Yang, and Haijian Lin. 2023. "ZmHMA3, a Member of the Heavy-Metal-Transporting ATPase Family, Regulates Cd and Zn Tolerance in Maize" International Journal of Molecular Sciences 24, no. 17: 13496. https://doi.org/10.3390/ijms241713496
APA StyleLiao, C., Li, Y., Wu, X., Wu, W., Zhang, Y., Zhan, P., Meng, X., Hu, G., Yang, S., & Lin, H. (2023). ZmHMA3, a Member of the Heavy-Metal-Transporting ATPase Family, Regulates Cd and Zn Tolerance in Maize. International Journal of Molecular Sciences, 24(17), 13496. https://doi.org/10.3390/ijms241713496