
Simultaneous Irradiation with UV-A, -B, and -C Lights Promotes Effective Decontamination of Planktonic and Sessile Bacteria: A Pilot Study

 ,
,  , , ,
, , ,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Combination of UV-A, UV-B, and UV-C Radiations: Determination of Irradiance and Energy Dose (Fluence)
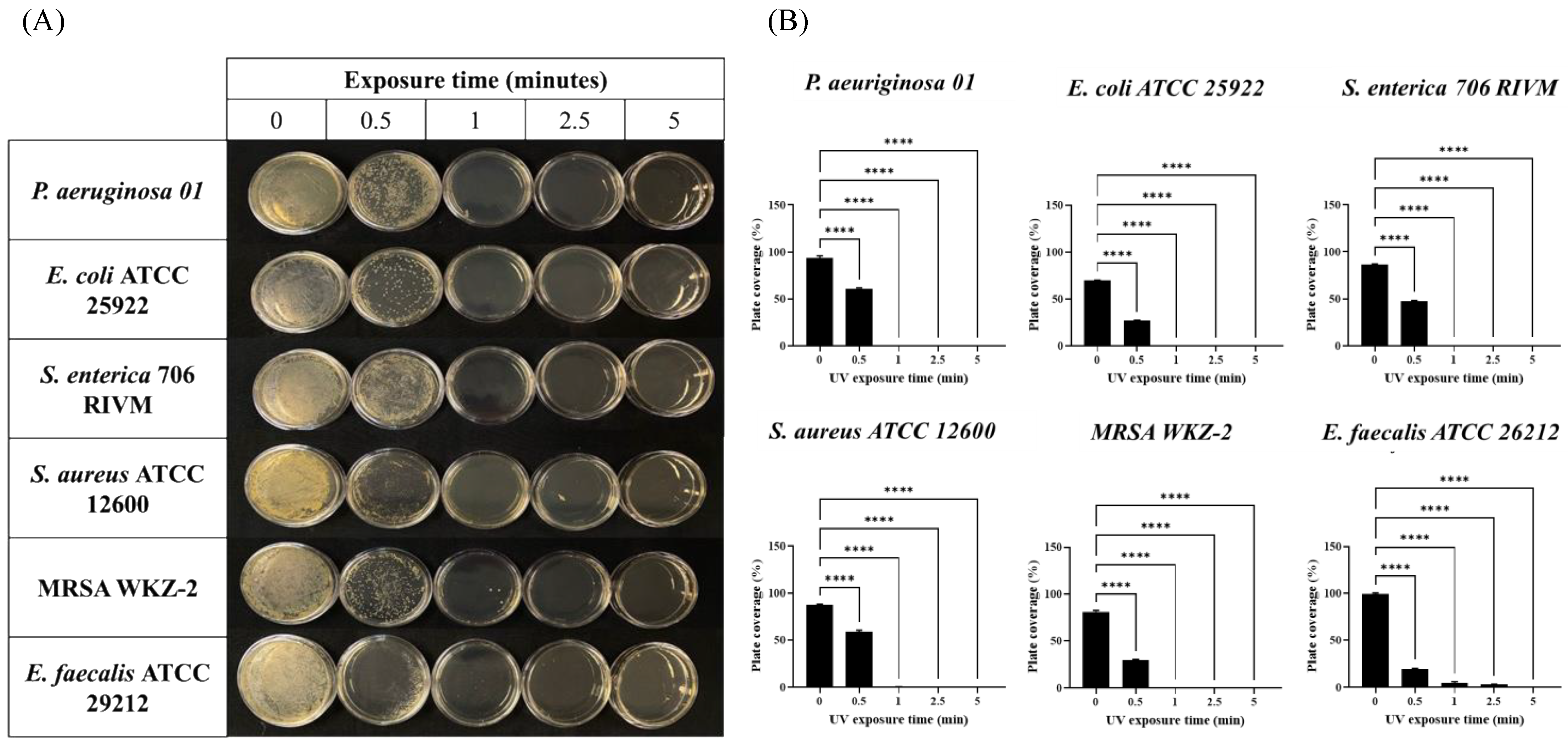
2.2. Combined UV Radiations Possess Fast Antimicrobial Activity
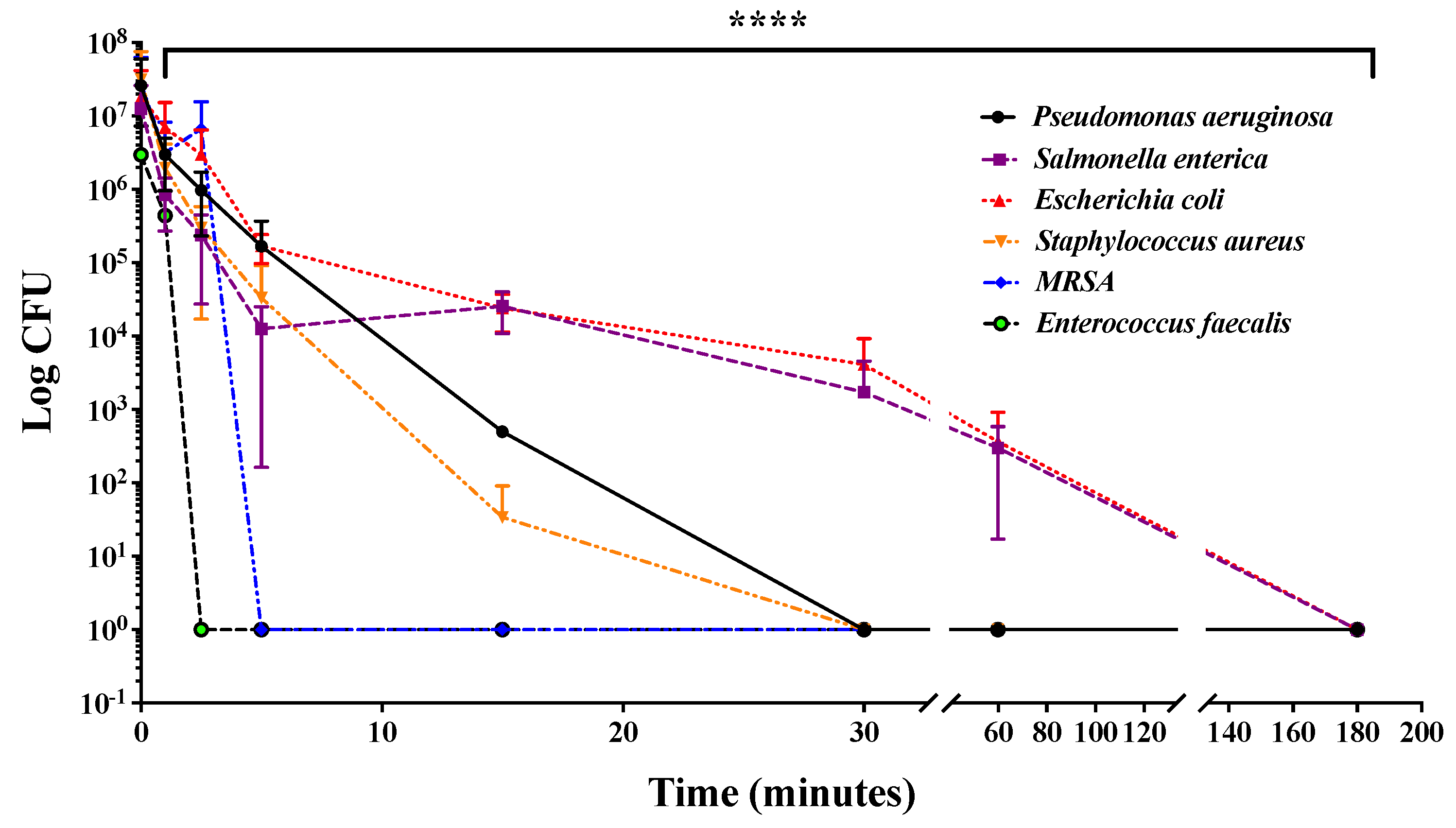
2.3. Combined UV Radiations Reduce Bacterial Contamination and Delay Bacterial Growth
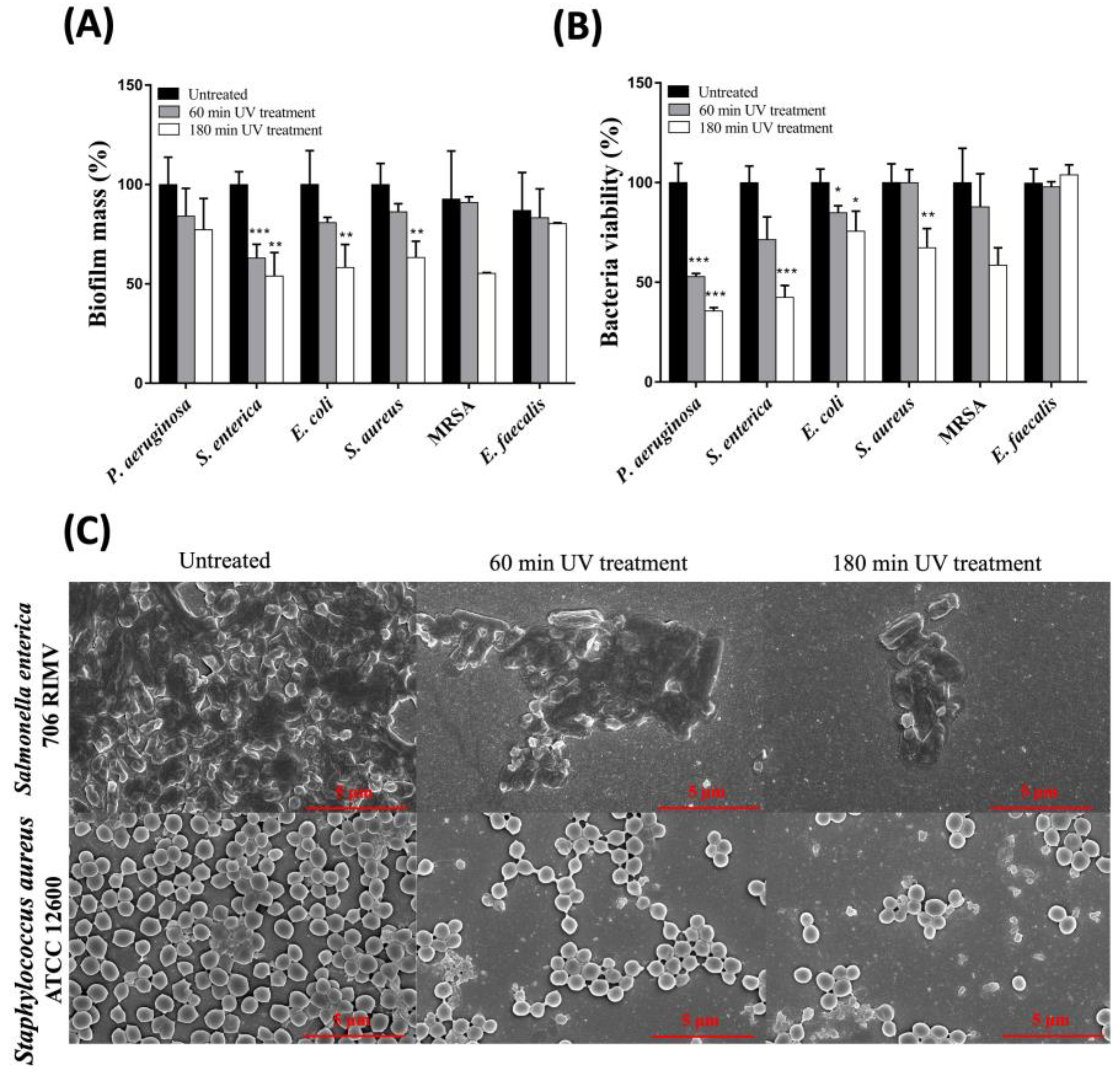
2.4. Combination of UV-A, -B, and -C Radiations Effectively Hinders the Stability of Biofilm Matrix and Reduces the Viability of Biofilm-Embedded Bacteria
3. Discussion
4. Materials and Methods
4.1. Irradiance and Energy Dose Measurement
4.2. Bacterial Strains
4.3. Antimicrobial Activity
4.4. Antibiofilm Activity
4.5. Scanning Electron Microscopy (SEM)
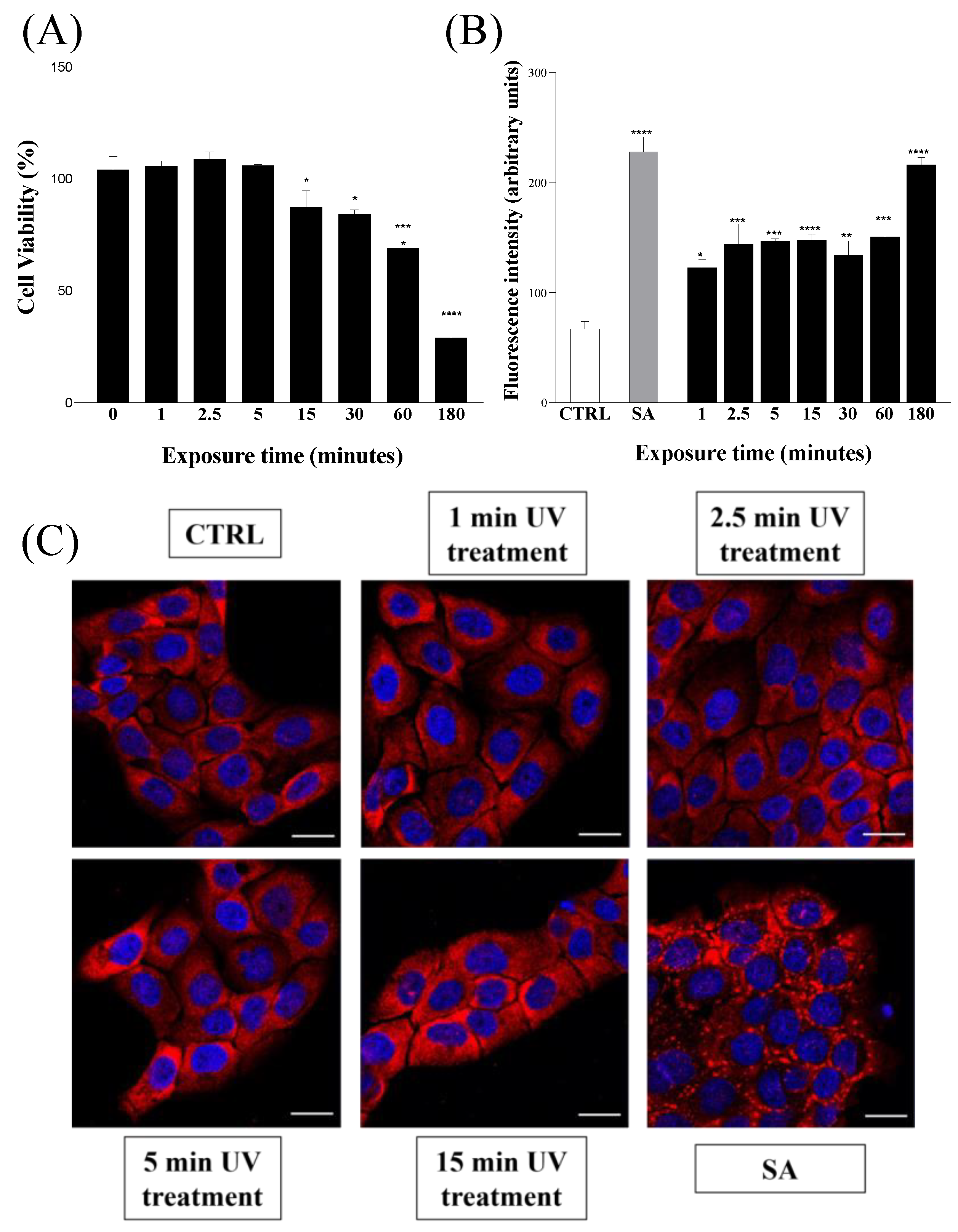
4.6. Metabolic Activity of Eukaryotic Cells
4.7. ROS Quantification Assay
4.8. Immunofluorescence
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Livingston, S.H.; Cadnum, J.L.; Benner, K.J.; Donskey, C.J. Efficacy of an Ultraviolet-A Lighting System for Continuous Decontamination of Health Care–Associated Pathogens on Surfaces. Am. J. Infect. Control 2020, 48, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Greene, A.K.; Few, B.K.; Serafini, J.C. Serafini A Comparison of Ozonation and Chlorination for the Disinfection of Stainless Steel Surfaces. J. Dairy Sci. 1993, 76, 3617–3620. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.M. Alcohols as Surface Disinfectants in Healthcare Settings. Infect. Control Hosp. Epidemiol. 2018, 39, 323–328. [Google Scholar] [CrossRef]
- Srivastav, A.L.; Patel, N.; Chaudhary, V.K. Disinfection By-Products in Drinking Water: Occurrence, Toxicity and Abatement. Environ. Pollut. 2020, 267, 115474. [Google Scholar] [CrossRef]
- Bharti, B.; Li, H.; Ren, Z.; Zhu, R.; Zhu, Z. Recent Advances in Sterilization and Disinfection Technology: A Review. Chemosphere 2022, 308, 136404. [Google Scholar] [CrossRef] [PubMed]
- Rutala, W.A.; Weber, D.J. Disinfection, Sterilization, and Antisepsis: Principles, Practices, Current Issues, New Research, and New Technologies. Am. J. Infect. Control 2019, 47, A1–A2. [Google Scholar] [CrossRef] [PubMed]
- Karaböce, B.; Saban, E.; Böyük, A.A.; Durmuş, H.O.; Hamid, R.; Baş, A. Inactivation of Viruses on Surfaces by Infrared Techniques. Int. J. Therm. Sci. 2022, 179, 107595. [Google Scholar] [CrossRef]
- Ölmez, H.; Kretzschmar, U. Potential Alternative Disinfection Methods for Organic Fresh-Cut Industry for Minimizing Water Consumption and Environmental Impact. LWT 2009, 42, 686–693. [Google Scholar] [CrossRef]
- Fraise, A.P. Choosing Disinfectants. J. Hosp. Infect. 1999, 43, 255–264. [Google Scholar] [CrossRef]
- Rudhart, S.A.; Günther, F.; Dapper, L.; Stuck, B.A.; Hoch, S. UV-C Light-Based Surface Disinfection: Analysis of Its Virucidal Efficacy Using a Bacteriophage Model. Int. J. Environ. Res. Public Health 2022, 19, 3246. [Google Scholar] [CrossRef]
- Song, K.; Mohseni, M.; Taghipour, F. Application of Ultraviolet Light-Emitting Diodes (UV-LEDs) for Water Disinfection: A Review. Water Res. 2016, 94, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Ploydaeng, M.; Rajatanavin, N.; Rattanakaemakorn, P. UV-C Light: A Powerful Technique for Inactivating Microorganisms and the Related Side Effects to the Skin. Photodermatol. Photoimmunol. Photomed. 2021, 37, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, E.; Rodríguez, C.; Ródenas, J.; de Rozas, A.P.; Segalés, J.; Pujols, J.; Polo, J. Ultraviolet (UV-C) Inactivation of Enterococcus Faecium, Salmonella Choleraesuis and Salmonella Typhimurium in Porcine Plasma. PLoS ONE 2017, 12, e0175289. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.P. UV-Induced DNA Damage and Repair: A Review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Häder, D.P. Physiological Aspects of UV-Excitation of DNA. In Photoinduced Phenomena in Nucleic Acids II; Barbatti, M., Borin, A., Ullrich, S., Eds.; Topics in Current Chemistry; Springer: Cham, Switzerland, 2014; Volume 356, pp. 203–248. [Google Scholar] [CrossRef]
- Bae, J.M.; Jung, H.M.; Hong, B.Y.; Lee, J.H.; Choi, W.J.; Lee, J.H.; Kim, G.M. Phototherapy for Vitiligo: A Systematic Review and Meta-Analysis Supplemental Content. JAMA Dermatol. 2017, 153, 666–674. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, M.X. A Clinical Review of Phototherapy for Psoriasis. Lasers Med. Sci. 2017, 33, 173–180. [Google Scholar] [CrossRef]
- Smith, H.L.; Howland, M.C.; Szmodis, A.W.; Li, Q.; Daemen, L.L.; Parikh, A.N.; Majewski, J. Early Stages of Oxidative Stress-Induced Membrane Permeabilization: A Neutron Reflectometry Study. J. Am. Chem. Soc. 2009, 131, 3631–3638. [Google Scholar] [CrossRef]
- Totonchy, M.B.; Chiu, M.W. UV-Based Therapy. Dermatol. Clin. 2014, 32, 399–413. [Google Scholar] [CrossRef]
- Kvam, E.; Davis, B.; Benner, K. Comparative Assessment of Pulsed and Continuous LED UV-A Lighting for Disinfection of Contaminated Surfaces. Life 2022, 12, 1747. [Google Scholar] [CrossRef]
- Allahyari, E.; Carraturo, F.; De Risi, A.; Nappo, A.; Morelli, M.; Cajora, A.; Guida, M. A Sequential Utilization of the UV-A (365 Nm) Fluence Rate for Disinfection of Water, Contaminated with Legionella Pneumophila and Legionella Dumoffii. Environ. Pollut. 2022, 304, 119224. [Google Scholar] [CrossRef]
- Kebbi, Y.; Muhammad, A.I.; Sant’Ana, A.S.; do Prado-Silva, L.; Liu, D.; Ding, T. Recent Advances on the Application of UV-LED Technology for Microbial Inactivation: Progress and Mechanism. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3501–3527. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.R.; Cotton, C.A. The Ultraviolet Disinfection Handbook; American Water Works Association: Denver, CO, USA, 2008. [Google Scholar]
- Chatterley, C.; Linden, K. Demonstration and Evaluation of Germicidal UV-LEDs for Point-of-Use Water Disinfection. J. Water Health 2010, 8, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Autin, O.; Romelot, C.; Rust, L.; Hart, J.; Jarvis, P.; MacAdam, J.; Parsons, S.A.; Jefferson, B. Evaluation of a UV-Light Emitting Diodes Unit for the Removal of Micropollutants in Water for Low Energy Advanced Oxidation Processes. Chemosphere 2013, 92, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Taghipour, F.; Mohseni, M. Microorganisms Inactivation by Wavelength Combinations of Ultraviolet Light-Emitting Diodes (UV-LEDs). Sci. Total Environ. 2019, 665, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Würtele, M.A.; Kolbe, T.; Lipsz, M.; Külberg, A.; Weyers, M.; Kneissl, M.; Jekel, M. Application of GaN-Based Ultraviolet-C Light Emitting Diodes—UV LEDs—For Water Disinfection. Water Res. 2011, 45, 1481–1489. [Google Scholar] [CrossRef]
- Jo, W.K.; Tayade, R.J. New Generation Energy-Efficient Light Source for Photocatalysis: LEDs for Environmental Applications. Ind. Eng. Chem. Res. 2014, 53, 2073–2084. [Google Scholar] [CrossRef]
- Beck, S.E.; Ryu, H.; Boczek, L.A.; Cashdollar, J.L.; Jeanis, K.M.; Rosenblum, J.S.; Lawal, O.R.; Linden, K.G. Evaluating UV-C LED Disinfection Performance and Investigating Potential Dual-Wavelength Synergy. Water Res. 2017, 109, 207–216. [Google Scholar] [CrossRef]
- Green, A.; Popović, V.; Pierscianowski, J.; Biancaniello, M.; Warriner, K.; Koutchma, T. Inactivation of Escherichia coli, Listeria and Salmonella by Single and Multiple Wavelength Ultraviolet-Light Emitting Diodes. Innov. Food Sci. Emerg. Technol. 2018, 47, 353–361. [Google Scholar] [CrossRef]
- Nakahashi, M.; Mawatari, K.; Hirata, A.; Maetani, M.; Shimohata, T.; Uebanso, T.; Hamada, Y.; Akutagawa, M.; Kinouchi, Y.; Takahashi, A. Simultaneous Irradiation with Different Wavelengths of Ultraviolet Light Has Synergistic Bactericidal Effect on Vibrio Parahaemolyticus. Photochem. Photobiol. 2014, 90, 1397–1403. [Google Scholar] [CrossRef]
- Oguma, K.; Katayama, H.; Ohgaki, S. Photoreactivation of Escherichia coli after Low- or Medium-Pressure UV Disinfection Determined by an Endonuclease Sensitive Site Assay. Appl. Environ. Microbiol. 2002, 68, 6029–6035. [Google Scholar] [CrossRef]
- Poepping, C.; Beck, S.E.; Wright, H.; Linden, K.G. Evaluation of DNA Damage Reversal during Medium-Pressure UV Disinfection. Water Res. 2014, 56, 181–189. [Google Scholar] [CrossRef]
- Lui, G.Y.; Roser, D.; Corkish, R.; Ashbolt, N.J.; Stuetz, R. Point-of-Use Water Disinfection Using Ultraviolet and Visible Light-Emitting Diodes. Sci. Total Environ. 2016, 553, 626–635. [Google Scholar] [CrossRef]
- Argyraki, A.; Markvart, M.; Stavnsbjerg, C.; Kragh, K.N.; Ou, Y.; Bjørndal, L.; Bjarnsholt, T.; Petersen, P.M. UV Light Assisted Antibiotics for Eradication of in Vitro Biofilms. Sci. Rep. 2018, 8, 16360. [Google Scholar] [CrossRef] [PubMed]
- Amar, A.; Castro, C.; Bernardelli, C.; Costa, C.S.; Donati, E. Influence of UVA Radiation on Growth, Biofilm Formation and Bioleaching Capacity of Leptospirillum Ferrooxidans. Hydrometallurgy 2021, 201, 105574. [Google Scholar] [CrossRef]
- Keyse, S.M.; Tyrrell, R.M. Heme Oxygenase Is the Major 32-KDa Stress Protein Induced in Human Skin Fibroblasts by UVA Radiation, Hydrogen Peroxide, and Sodium Arsenite. Proc. Nati. Acad. Sci. USA 1989, 86, 99–103. [Google Scholar] [CrossRef]
- Yu, H.; Kuang, M.; Wang, Y.; Rodeni, S.; Wei, Q.; Wang, W.; Mao, D. Sodium Arsenite Injection Induces Ovarian Oxidative Stress and Affects Steroidogenesis in Rats. Biol. Trace Elem. Res. 2019, 189, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Culurciello, R.; Bosso, A.; Troisi, R.; Barrella, V.; Di Nardo, I.; Borriello, M.; Gaglione, R.; Pistorio, V.; Aceto, S.; Cafaro, V.; et al. Protective Effects of Recombinant Human Angiogenin in Keratinocytes: New Insights on Oxidative Stress Response Mediated by RNases. Int. J. Mol. Sci. 2022, 23, 8781. [Google Scholar] [CrossRef]
- Moutaoufik, M.T.; El Fatimy, R.; Nassour, H.; Gareau, C.; Lang, J.; Tanguay, R.M.; Mazroui, R.; Khandjian, E.W. UVC-Induced Stress Granules in Mammalian Cells. PLoS ONE 2014, 9, e112742. [Google Scholar] [CrossRef]
- Fuentes-Villalobos, F.; Farkas, C.; Riquelme-Barrios, S.; Armijo, M.E.; Soto-Rifo, R.; Pincheira, R.; Castro, A.F. DISC1 Promotes Translation Maintenance during Sodium Arsenite-Induced Oxidative Stress. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 657–669. [Google Scholar] [CrossRef]
- Gullian, M.; Espinosa-Faller, F.J.; Núñez, A.; López-Barahona, N. Effect of Turbidity on the Ultraviolet Disinfection Performance in Recirculating Aquaculture Systems with Low Water Exchange. Aquac. Res. 2012, 43, 595–606. [Google Scholar] [CrossRef]
- Cutler, T.D.; Wang, C.; Hoff, S.J.; Zimmerman, J.J. Effect of Temperature and Relative Humidity on Ultraviolet (UV 254) Inactivation of Airborne Porcine Respiratory and Reproductive Syndrome Virus. Vet. Microbiol. 2012, 159, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, N.; Perumal, G.; Doble, M. Bacterial Resistance in Biofilm-Associated Bacteria. Future Microbiol. 2015, 10, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.C.R. Biofilms: Microbial Strategies for Surviving UV Exposure. Adv. Exp. Med. Biol. 2017, 996, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, R.K.; Baun, A.; Andersen, H.R. Acute Toxicity and Risk Evaluation of the CSO Disinfectants Performic Acid, Peracetic Acid, Chlorine Dioxide and Their by-Products Hydrogen Peroxide and Chlorite. Sci. Total Environ. 2019, 677, 1–8. [Google Scholar] [CrossRef]
- Hatanaka, N.; Xu, B.; Yasugi, M.; Morino, H.; Tagishi, H.; Miura, T.; Shibata, T.; Yamasaki, S. Chlorine Dioxide Is a More Potent Antiviral Agent against SARS-CoV-2 than Sodium Hypochlorite. J. Hosp. Infect. 2021, 118, 20–26. [Google Scholar] [CrossRef]
- Ahmad, I.; Malak, H.A.; Abulreesh, H.H. Environmental Antimicrobial Resistance and Its Drivers: A Potential Threat to Public Health. J. Glob. Antimicrob. Resist. 2021, 27, 101–111. [Google Scholar] [CrossRef]
- Balakrishnan, A.; Jacob, M.M.; Kumar, P.S.; Kapoor, A.; Ponnuchamy, M.; Sivaraman, P.; Sillanpää, M. Strategies for Safe Management of Hospital Wastewater during the COVID-19 Pandemic. Int. J. Environ. Sci. Technol. 2023. [Google Scholar] [CrossRef]
- Rutala, W.A.; Weber, D.J. Disinfection and Sterilization in Health Care Facilities: An Overview and Current Issues. Infect. Dis. Clin. N. Am. 2016, 30, 609–637. [Google Scholar] [CrossRef]
- Hamamoto, A.; Mori, M.; Takahashi, A.; Nakano, M.; Wakikawa, N.; Akutagawa, M.; Ikehara, T.; Nakaya, Y.; Kinouchi, Y. New Water Disinfection System Using UVA Light-Emitting Diodes. J. Appl. Microbiol. 2007, 103, 2291–2298. [Google Scholar] [CrossRef]
- Nyangaresi, P.O.; Qin, Y.; Chen, G.; Zhang, B.; Lu, Y.; Shen, L. Effects of Single and Combined UV-LEDs on Inactivation and Subsequent Reactivation of E. Coli in Water Disinfection. Water Res. 2018, 147, 331–341. [Google Scholar] [CrossRef]
- Schwinn, K.; Ferré, N.; Huix-Rotllant, M. UV-Visible Absorption Spectrum of FAD and Its Reduced Forms Embedded in a Cryptochrome Protein. Phys. Chem. Chem. Phys. 2020, 22, 12447–12455. [Google Scholar] [CrossRef] [PubMed]
- Rahman, G.; Lim, J.Y.; Yeon Lim, J.; Jung, K.-D.; Joo, O.-S. Electrodeposited Ru Nanoparticles for Electrochemical Reduction of NAD + to NADH Metal Sulfides as Electrocatalyst for Hydrogen Evolution Reaction View Project Conversion of CO2 to Formic Acid/Formate View Project Electrodeposited Ru Nanoparticles for Electrochemical Reduction of NAD + to NADH. Int. J. Electrochem. Sci. 2011, 6, 2789–2797. [Google Scholar]
- Bolton, J.R.; Beck, S.E.; Linden, K.G. Protocol for the Determination of Fluence (UV Dose) Using A Low-Pressure or Low-Pressure High-Output UV Lamp in Bench-Scale Collimated Beam Ultraviolet Experiments. IUVA News 2015, 17, 11–16. [Google Scholar]
- Takada, A.; Matsushita, K.; Horioka, S.; Furuichi, Y.; Sumi, Y. Bactericidal Effects of 310 Nm Ultraviolet Light-Emitting Diode Irradiation on Oral Bacteria. BMC Oral Health 2017, 17, 96. [Google Scholar] [CrossRef]
- Brancaccio, D.; Pizzo, E.; Cafaro, V.; Notomista, E.; De Lise, F.; Bosso, A.; Gaglione, R.; Merlino, F.; Novellino, E.; Ungaro, F.; et al. Antimicrobial Peptide Temporin-L Complexed with Anionic Cyclodextrins Results in a Potent and Safe Agent against Sessile Bacteria. Int. J. Pharm. 2020, 584, 119437. [Google Scholar] [CrossRef]
- Rezaie, A.; Leite, G.G.S.; Melmed, G.Y.; Mathur, R.; Villanueva-Millan, M.J.; Parodi, G.; Sin, J.; Germano, J.F.; Morales, W.; Weitsman, S.; et al. Ultraviolet a Light Effectively Reduces Bacteria and Viruses Including Coronavirus. PLoS ONE 2020, 15, e0236199. [Google Scholar] [CrossRef]
- Baykuş, G.; Akgün, M.P.; Unluturk, S. Effects of Ultraviolet-Light Emitting Diodes (UV-LEDs) on Microbial Inactivation and Quality Attributes of Mixed Beverage Made from Blend of Carrot, Carob, Ginger, Grape and Lemon Juice. Innov. Food Sci. Emerg. Technol. 2021, 67, 102572. [Google Scholar] [CrossRef]
- Pane, K.; Cafaro, V.; Avitabile, A.; Torres, M.D.T.; Vollaro, A.; De Gregorio, E.; Catania, M.R.; Di Maro, A.; Bosso, A.; Gallo, G.; et al. Identification of Novel Cryptic Multifunctional Antimicrobial Peptides from the Human Stomach Enabled by a Computational-Experimental Platform. ACS Synth. Biol. 2018, 7, 2105–2115. [Google Scholar] [CrossRef]
- Bosso, A.; Pirone, L.; Gaglione, R.; Pane, K.; Del Gatto, A.; Zaccaro, L.; Di Gaetano, S.; Diana, D.; Fattorusso, R.; Pedone, E.; et al. A New Cryptic Host Defense Peptide Identified in Human 11-Hydroxysteroid Dehydrogenase-1 β-like: From in Silico Identification to Experimental Evidence. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2342–2353. [Google Scholar] [CrossRef]
- Rangel, K.; Cabral, F.O.; Lechuga, G.C.; Villas-Bôas, M.H.S.; Midlej, V.; De-Simone, S.G. Effectiveness Evaluation of a UV-C-Photoinactivator against Selected ESKAPE-E Pathogens. Int. J. Environ. Res. Public Health 2022, 19, 16559. [Google Scholar] [CrossRef]
- Bosso, A.; Gaglione, R.; Di Girolamo, R.; Veldhuizen, E.J.A.; García-Vello, P.; Fusco, S.; Cafaro, V.; Monticelli, M.; Culurciello, R.; Notomista, E.; et al. Human Cryptic Host Defence Peptide GVF27 Exhibits Anti-Infective Properties against Biofilm Forming Members of the Burkholderia Cepacia Complex. Pharmaceuticals 2022, 15, 260. [Google Scholar] [CrossRef] [PubMed]
- Coorens, M.; Scheenstra, M.R.; Veldhuizen, E.J.A.; Haagsman, H.P. Interspecies Cathelicidin Comparison Reveals Divergence in Antimicrobial Activity, TLR Modulation, Chemokine Induction and Regulation of Phagocytosis. Sci. Rep. 2017, 7, 40874. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, N.L.; Banerjee, P. Inactivation of Escherichia Coli O157:H7 and Listeria monocytogenes in Biofilms by Pulsed Ultraviolet Light. BMC Res. Notes 2015, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Lampitella, E.; Landi, N.; Oliva, R.; Gaglione, R.; Bosso, A.; de Lise, F.; Ragucci, S.; Arciello, A.; Petraccone, L.; Pizzo, E.; et al. Toxicity and Membrane Perturbation Properties of the Ribotoxin-like Protein Ageritin. J. Biochem. 2021, 170, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, A.; Torres, M.D.T.; Gaglione, R.; Dell’olmo, E.; Di Girolamo, R.; Bosso, A.; Pizzo, E.; Haagsman, H.P.; Veldhuizen, E.J.A.; De La Fuente-Nunez, C.; et al. Synthetic Antibiotic Derived from Sequences Encrypted in a Protein from Human Plasma. ACS Nano 2022, 6, 1880–1895. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UV-C (2 × 120 W) | UV-B (60 W) | UV-A (60 W) | ||||
|---|---|---|---|---|---|---|
| Time (min) | Irradiance (mW/cm2) | Energy Dose (mJ/cm2) | Irradiance (mW/cm2) | Energy Dose (mJ/cm2) | Irradiance (mW/cm2) | Energy Dose (mJ/cm2) |
| 0 | 0.93 | 27.9 | 0.462 | 13.86 | 0.4 | 12 |
| 5 | 0.34 | 102 | 0.342 | 102.6 | 0.322 | 96.6 |
| 15 | 0.266 | 239.4 | 0.328 | 295.2 | 0.205 | 184.5 |
| 30 | 0.247 | 444.6 | 0.318 | 572.4 | 0.202 | 363.6 |
| 60 | 0.23 | 828 | 0.305 | 1098 | 0.19 | 684 |
| 180 | 0.21 | 2268 | 0.285 | 3078 | 0.175 | 1890 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosso, A.; Tortora, F.; Culurciello, R.; Di Nardo, I.; Pistorio, V.; Carraturo, F.; Colecchia, A.; Di Girolamo, R.; Cafaro, V.; Notomista, E.; et al. Simultaneous Irradiation with UV-A, -B, and -C Lights Promotes Effective Decontamination of Planktonic and Sessile Bacteria: A Pilot Study. Int. J. Mol. Sci. 2023, 24, 12951. https://doi.org/10.3390/ijms241612951
Bosso A, Tortora F, Culurciello R, Di Nardo I, Pistorio V, Carraturo F, Colecchia A, Di Girolamo R, Cafaro V, Notomista E, et al. Simultaneous Irradiation with UV-A, -B, and -C Lights Promotes Effective Decontamination of Planktonic and Sessile Bacteria: A Pilot Study. International Journal of Molecular Sciences. 2023; 24(16):12951. https://doi.org/10.3390/ijms241612951
Chicago/Turabian StyleBosso, Andrea, Francesca Tortora, Rosanna Culurciello, Ilaria Di Nardo, Valeria Pistorio, Federica Carraturo, Andrea Colecchia, Rocco Di Girolamo, Valeria Cafaro, Eugenio Notomista, and et al. 2023. "Simultaneous Irradiation with UV-A, -B, and -C Lights Promotes Effective Decontamination of Planktonic and Sessile Bacteria: A Pilot Study" International Journal of Molecular Sciences 24, no. 16: 12951. https://doi.org/10.3390/ijms241612951
APA StyleBosso, A., Tortora, F., Culurciello, R., Di Nardo, I., Pistorio, V., Carraturo, F., Colecchia, A., Di Girolamo, R., Cafaro, V., Notomista, E., Ingenito, R., & Pizzo, E. (2023). Simultaneous Irradiation with UV-A, -B, and -C Lights Promotes Effective Decontamination of Planktonic and Sessile Bacteria: A Pilot Study. International Journal of Molecular Sciences, 24(16), 12951. https://doi.org/10.3390/ijms241612951