
The Biostimulant, Potassium Humate Ameliorates Abiotic Stress in Arabidopsis thaliana by Increasing Starch Availability




Abstract
1. Introduction
2. Results
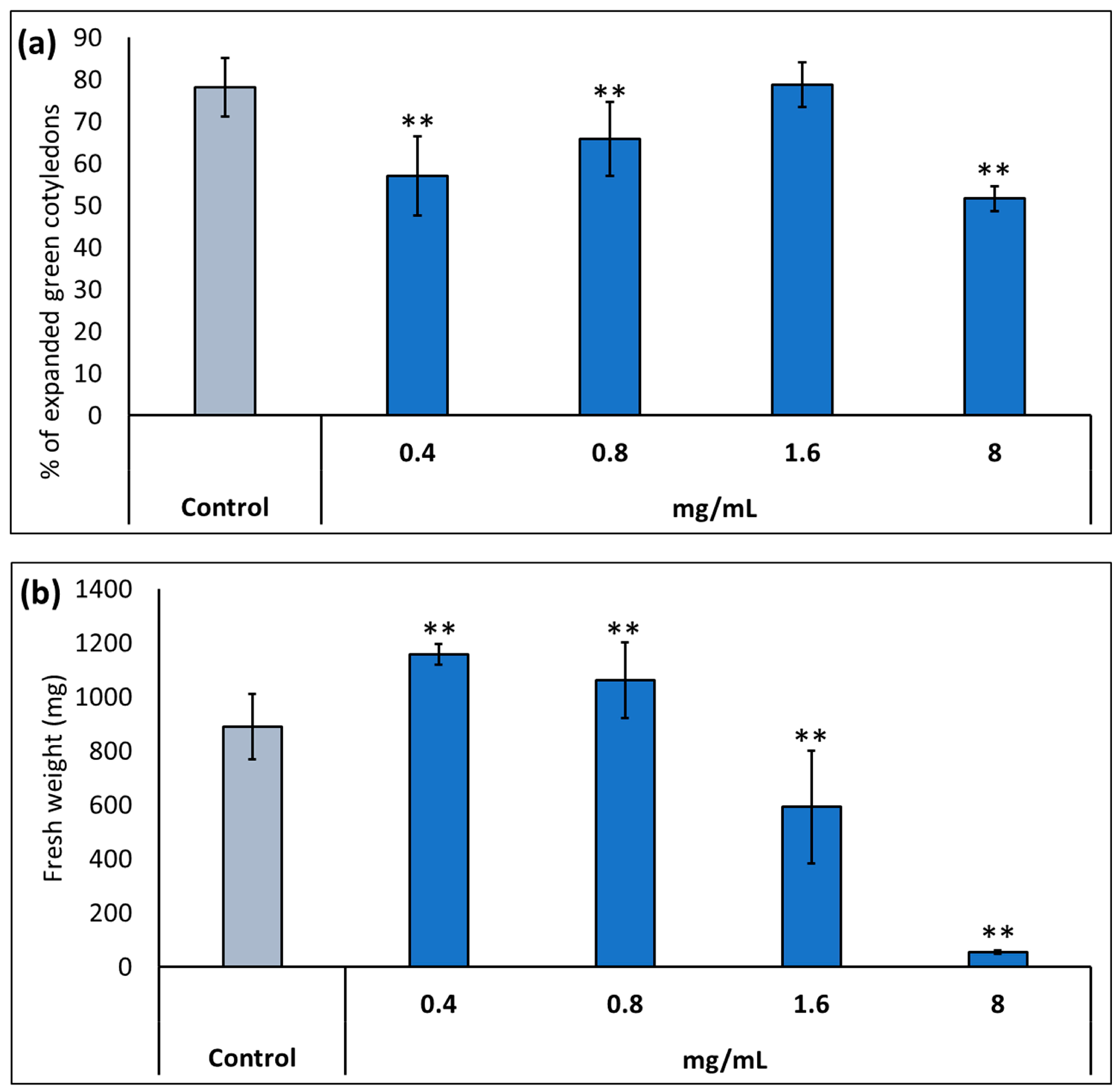
2.1. Effect of Potassium Humate (KH60) on the Growth of Arabidopsis thaliana in the Presence of Abiotic Stress
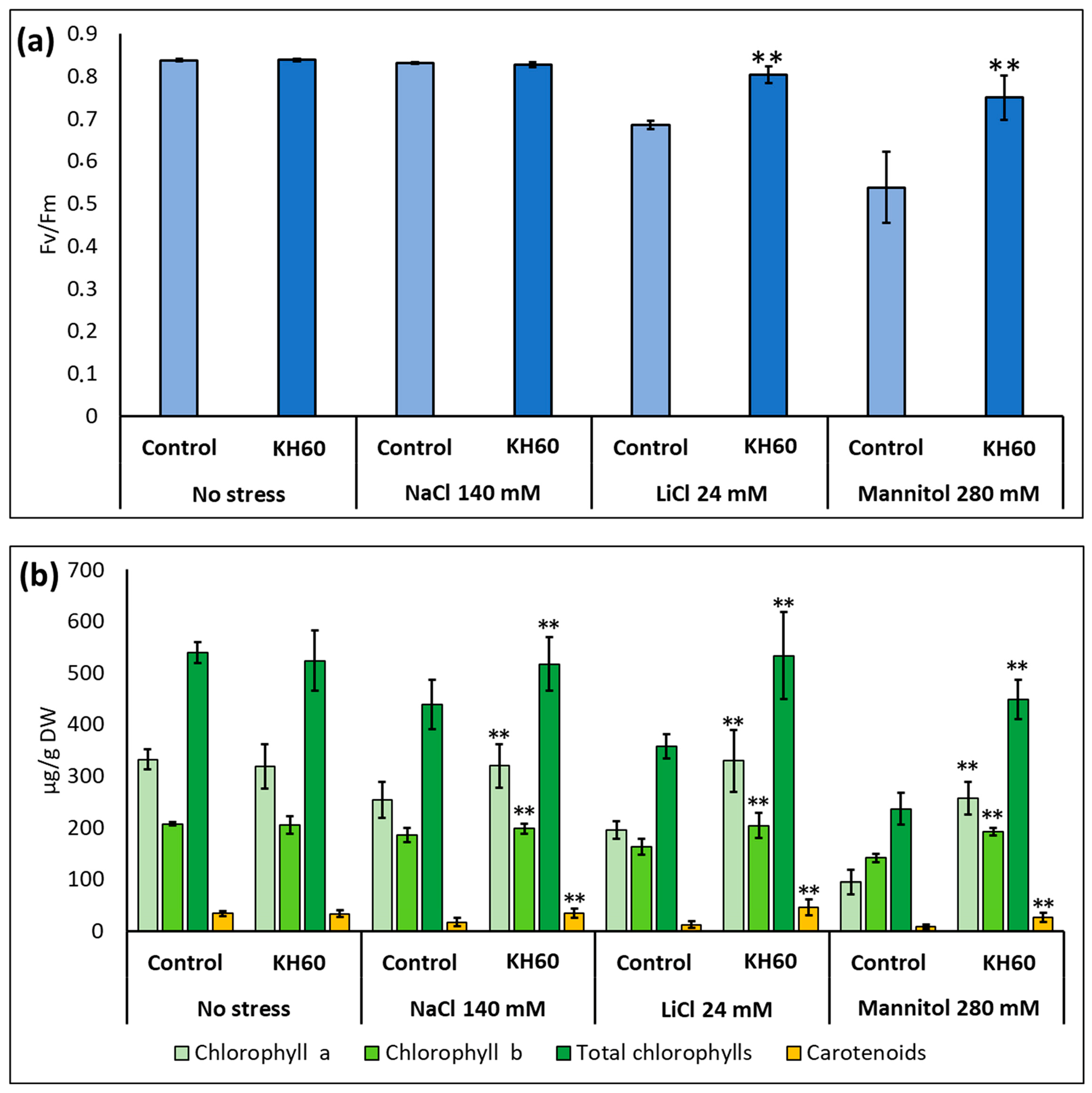
2.2. Photosynthetic Pigment Content and Photosystem II Yield Index of Arabidopsis Leaves
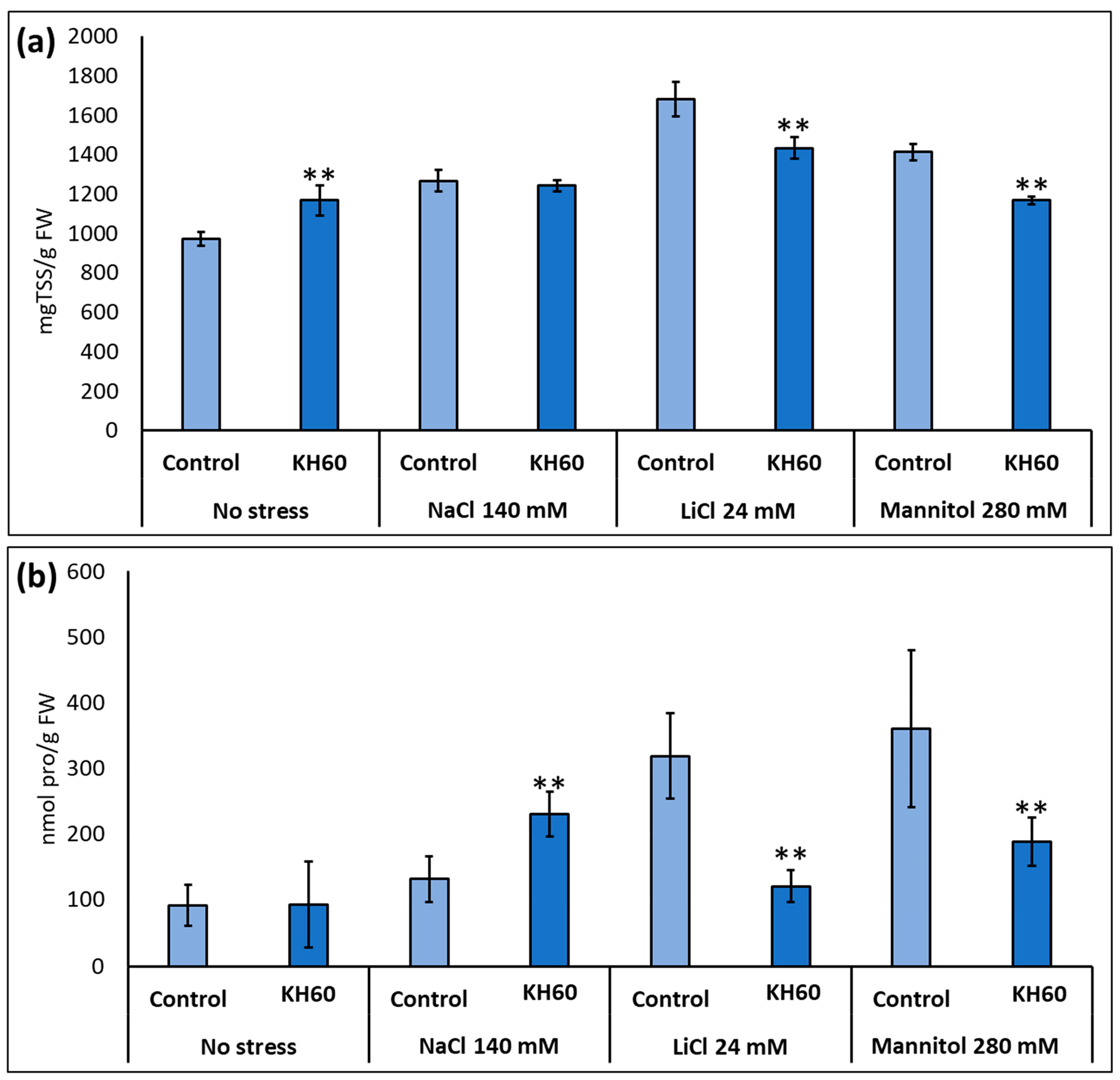
2.3. Effect of KH60 on the Content of Osmolytes and Non-Enzymatic Antioxidants under Abiotic Stress Conditions
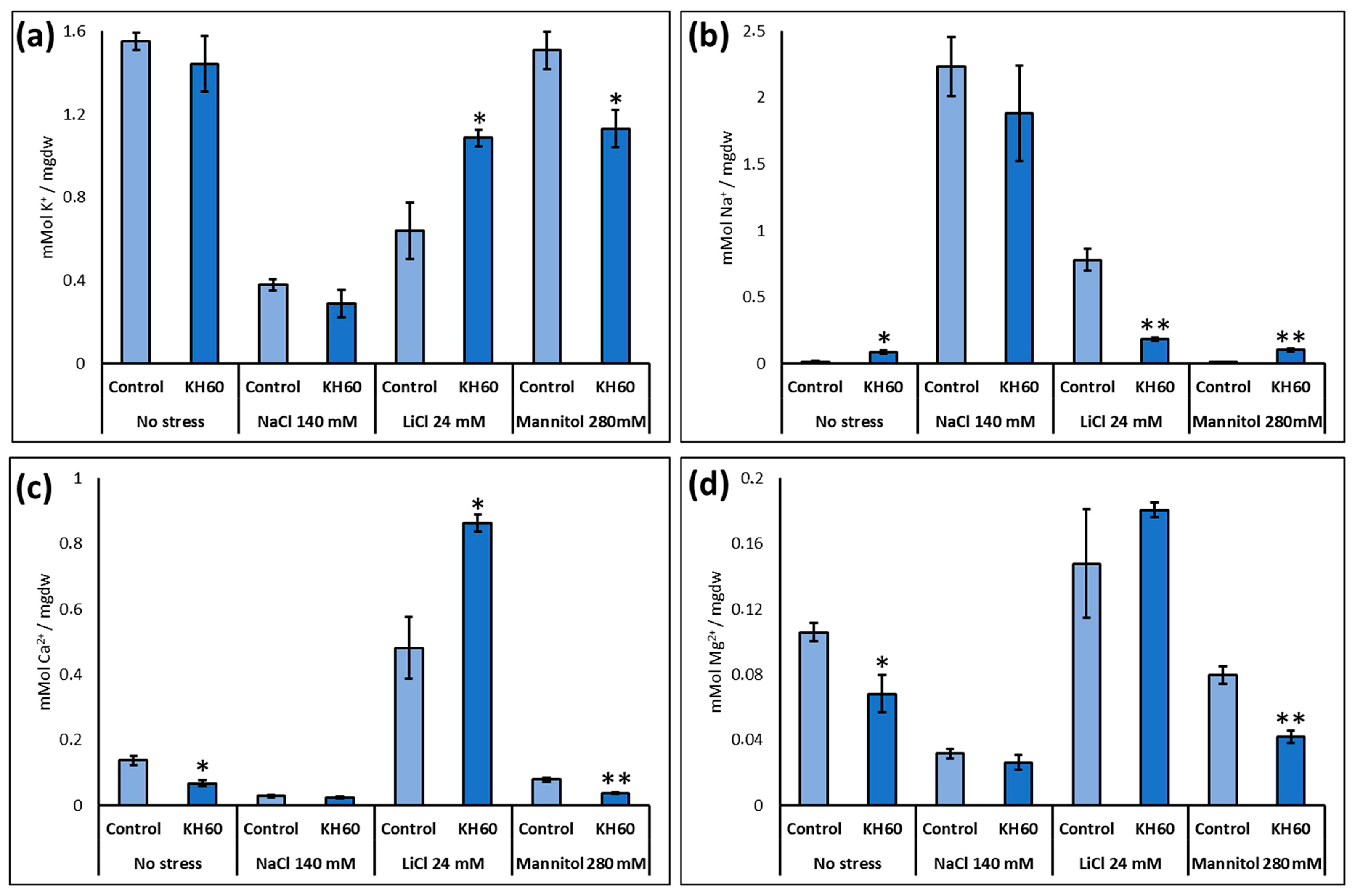
2.4. Effect of Stress and KH60 on Ion Content
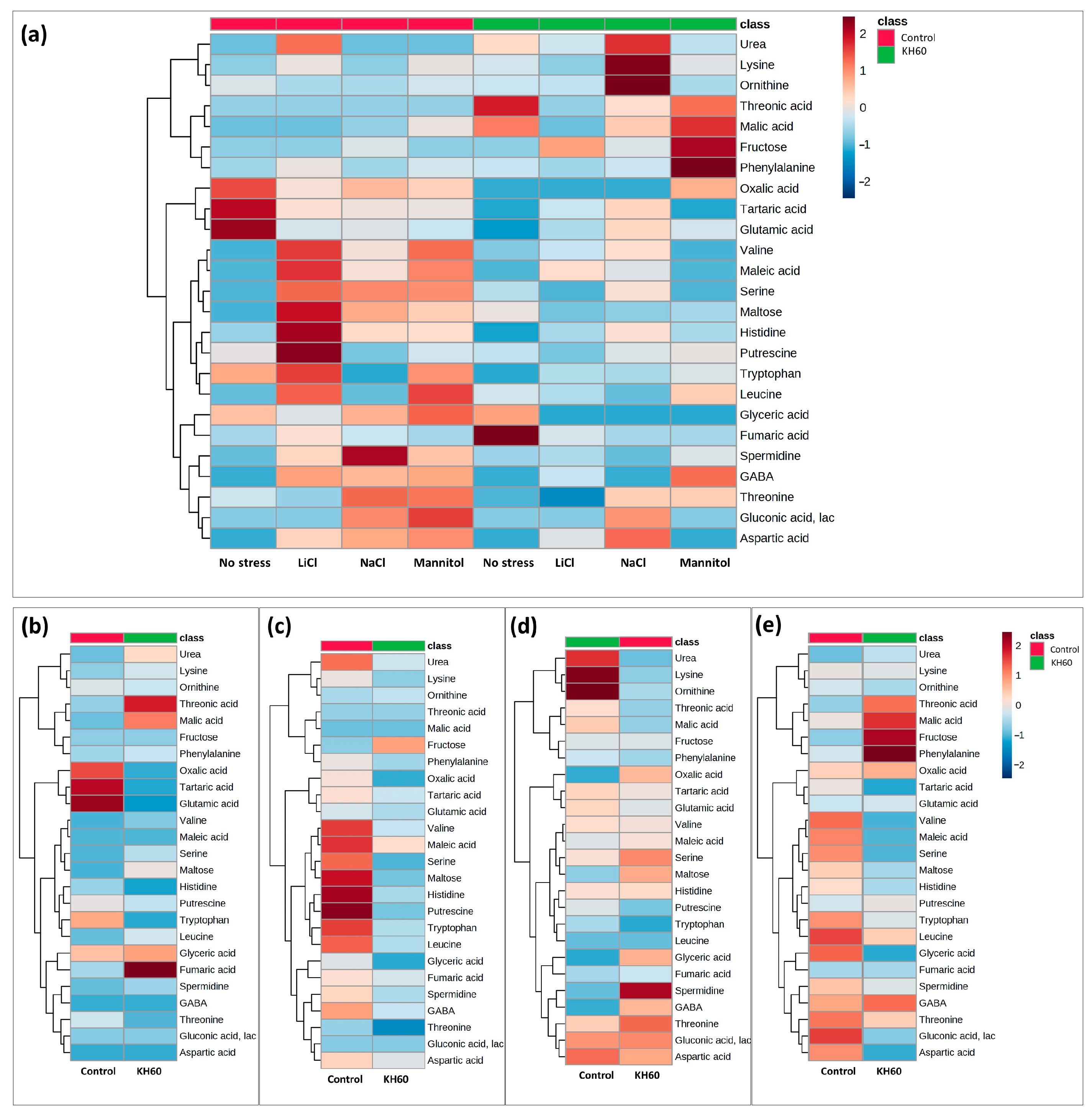
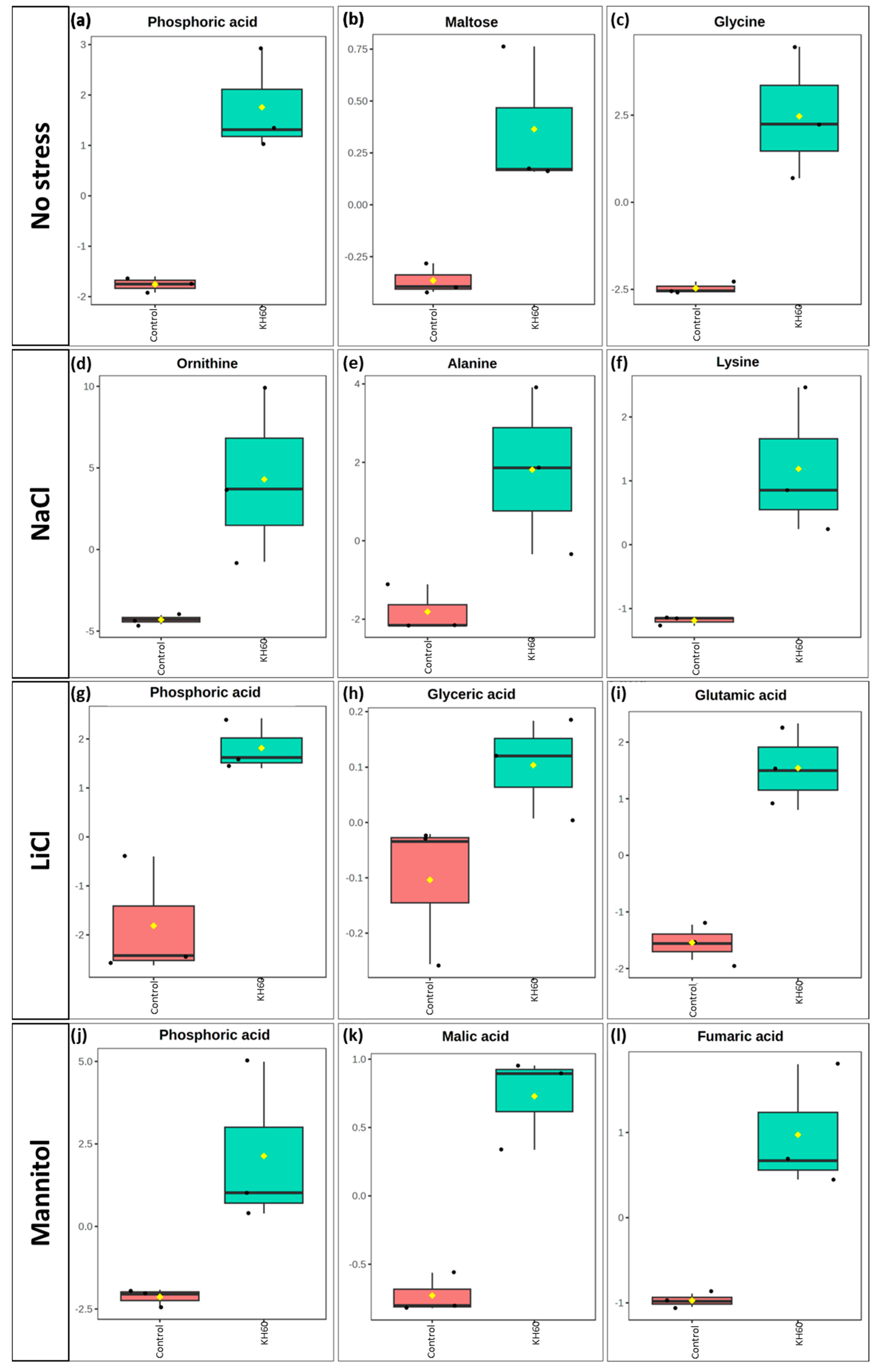
2.5. Effect of KH60 on the Arabidopsis Metabolome under Different Abiotic Stress Conditions
2.6. Effect of KH60 on Starch Levels
3. Discussion
4. Materials and Methods
4.1. Biostimulant Product, Plant Media, and Growth Conditions
4.2. Arabidopsis Germination and Early Development under Abiotic Stress
4.3. Leaf Photosynthetic Pigments and Photosystem II Yield
4.4. Proline and Total Sugars Determination
4.5. Total Phenols and Flavonoid Determination
4.6. Ion Content Determination
4.7. Staining of Arabidopsis Leaves with Lugol
4.8. Metabolomic Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osman, H.S.; Gowayed, S.M.; Elbagory, M.; Omara, A.E.D.; Abd El-Monem, A.M.; Abd El-Razek, U.A.; Hafez, E.M. Interactive Impacts of Beneficial Microbes and Si-Zn Nanocomposite on Growth and Productivity of Soybean Subjected to Water Deficit under Salt-Affected Soil Conditions. Plants 2021, 10, 1396. [Google Scholar] [CrossRef] [PubMed]
- Taïbi, K.; del Campo, A.D.; Aguado, A.; Mulet, J.M. The Effect of Genotype by Environment Interaction, Phenotypic Plasticity and Adaptation on Pinus Halepensis Reforestation Establishment under Expected Climate Drifts. Ecol. Eng. 2015, 84, 218–228. [Google Scholar] [CrossRef]
- Rossi, R. Desertification and Agriculture. EPRS | European Parliamentary Research Service, Members’ Research Service, PE 646.171–February 2020. 2020. Available online: https://www.europarl.europa.eu/RegData/etudes/BRIE/2020/646171/EPRS_BRI(2020)646171_EN.pdf (accessed on 25 July 2023).
- Reynolds, J.F.; Smith, D.M.S.; Lambin, E.F.; Turner, B.L.; Mortimore, M.; Batterbury, S.P.J.; Downing, T.E.; Dowlatabadi, H.; Fernández, R.J.; Herrick, J.E.; et al. Global Desertification: Building a Science for Dryland Development. Science 2007, 316, 847–851. [Google Scholar] [CrossRef]
- UNCCD. Drought in Numbers 2022: Restoration for Readiness and Resilience; United Nations Expert Group Meeting on Population Distribution, Urbanization, Internal Migration and Development: New York, NY, USA, 2022; p. 50. [Google Scholar]
- Kauffman, G.L.; Kneivel, D.P.; Watschke, T.L. Effects of a Biostimulant on the Heat Tolerance Associated with Photosynthetic Capacity, Membrane Thermostability, and Polyphenol Production of Perennial Ryegrass. Crop Sci. 2007, 47, 261–267. [Google Scholar] [CrossRef]
- Chevilly, S.; Dolz-Edo, L.; Morcillo, L.; Vilagrosa, A.; López-Nicolás, J.M.; Yenush, L.; Mulet, J.M. Identification of Distinctive Physiological and Molecular Responses to Salt Stress among Tolerant and Sensitive Cultivars of Broccoli (Brassica oleracea Var. Italica). BMC Plant Biol. 2021, 21, 488. [Google Scholar] [CrossRef] [PubMed]
- Chevilly, S.; Dolz-Edo, L.; López-Nicolás, J.M.; Morcillo, L.; Vilagrosa, A.; Yenush, L.; Mulet, J.M. Physiological and Molecular Characterization of the Differential Response of Broccoli (Brassica oleracea Var. Italica) Cultivars Reveals Limiting Factors for Broccoli Tolerance to Drought Stress. J. Agric. Food Chem. 2021, 69, 10394–10404. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil. 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. Chapter Two—The Use of Biostimulants for Enhancing Nutrient Uptake. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 130, pp. 141–174. ISBN 0065-2113. [Google Scholar]
- Kumari, M.; Swarupa, P.; Kesari, K.K.; Kumar, A. Microbial Inoculants as Plant Biostimulants: A Review on Risk Status. Life 2023, 13, 12. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The Effect of a Plant-Derived Biostimulant on Metabolic Profiling and Crop Performance of Lettuce Grown under Saline Conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Nardi, S.; Muscolo, A.; Vaccaro, S.; Baiano, S.; Spaccini, R.; Piccolo, A. Relationship between Molecular Characteristics of Soil Humic Fractions and Glycolytic Pathway and Krebs Cycle in Maize Seedlings. Soil. Biol. Biochem. 2007, 39, 3138–3146. [Google Scholar] [CrossRef]
- Sutton, R.; Sposito, G. Molecular Structure in Soil Humic Substances: The New View. Environ. Sci. Technol. 2005, 39, 9009–9015. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The Contentious Nature of Soil Organic Matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Regelink, I.C.; Stoof, C.R.; Rousseva, S.; Weng, L.; Lair, G.J.; Kram, P.; Nikolaidis, N.P.; Kercheva, M.; Banwart, S.; Comans, R.N.J. Linkages between Aggregate Formation, Porosity and Soil Chemical Properties. Geoderma 2015, 247–248, 24–37. [Google Scholar] [CrossRef]
- Yang, F.; Tang, C.; Antonietti, M. Natural and Artificial Humic Substances to Manage Minerals, Ions, Water, and Soil Microorganisms. Chem. Soc. Rev. 2021, 50, 6221–6239. [Google Scholar] [CrossRef]
- Huang, P.M.; Hardie, A.G. Formation Mechanisms of Humic Substances in the Environment. In Biophysico-Chemical Processes Involving Natural Nonliving Organic Matter in Environmental Systems; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 41–109. ISBN 9780470494950. [Google Scholar]
- Stevenson, F.J.; Olsen, R.A. A Simplified Representation of the Chemical Nature and Reactions of Soil Humus. J. Agron. Educ. 1989, 18, 84–88. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous Potassium Treatments Elevate Salt Tolerance and Performances of Glycine max L. By Boosting Antioxidant Defense System under Actual Saline Field Conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Idrees, M.; Anjum, M.A.; Mirza, J.I. Potassium Humate and NPK Application Rates Influence Yield and Economic Performance of Potato Crops Grown in Clayey Loam Soils. Soil. Environ. 2018, 37, 53–61. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, A.P.; Raha, P.; Rakshit, A.; Singh, C.M.; Kishor, P. Potassium Humate: A Potential Soil Conditioner and Plant Growth Promoter. Int. J. Agric. Environ. Biotechnol. 2013, 6, 441. [Google Scholar] [CrossRef]
- Serrano, R.; Mulet, J.M.; Rios, G.; Marquez, J.A.; Larrinoa, I.I.F.d.; Leube, M.P.; Mendizabal, I.; Pascual-Ahuir, A.; Proft, M.; Ros, R.; et al. A Glimpse of the Mechanisms of Ion Homeostasis during Salt Stress. J. Exp. Bot. 1999, 50, 1023–1036. [Google Scholar] [CrossRef]
- Canellas, L.P.; Canellas, N.O.A.; da S Irineu, L.E.S.; Olivares, F.L.; Piccolo, A. Plant Chemical Priming by Humic Acids. Chem. Biol. Technol. Agric. 2020, 7, 12. [Google Scholar] [CrossRef]
- Kanojia, A.; Dijkwel, P.P. Abiotic Stress Responses Are Governed by Reactive Oxygen Species and Age. Annu. Plant Rev. 2018, 1, 295–326. [Google Scholar]
- Maggioni, A.; Varanini, Z.; Nardi, S.; Pinton, R. Action of Soil Humic Matter on Plant Roots: Stimulation of Ion Uptake and Effects on(Mg2+ + K+) ATPase Activity. Sci. Total Environ. 1987, 62, 355–363. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Muscolo, A.; Vianello, A. Physiological Effects of Humic Substances on Higher Plants. Soil. Biol. Biochem. 2002, 34, 1527–1536. [Google Scholar] [CrossRef]
- Trevisan, S.; Botton, A.; Vaccaro, S.; Vezzaro, A.; Quaggiotti, S.; Nardi, S. Humic Substances Affect Arabidopsis Physiology by Altering the Expression of Genes Involved in Primary Metabolism, Growth and Development. Environ. Exp. Bot. 2011, 74, 45–55. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Microarray Analysis of Humic Acid Effects on Brassica Napus Growth: Involvement of N, C and S Metabolisms. Plant Soil. 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Roomi, S.; Masi, A.; Conselvan, G.B.; Trevisan, S.; Quaggiotti, S.; Pivato, M.; Arrigoni, G.; Yasmin, T.; Carletti, P. Protein Profiling of Arabidopsis Roots Treated With Humic Substances: Insights Into the Metabolic and Interactome Networks. Front. Plant Sci. 2018, 9, 1812. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, N.O.; Olivares, F.L.; Novotny, E.H.; Canellas, L.P. Changes in Metabolic Profiling of Sugarcane Leaves Induced by Endophytic Diazotrophic Bacteria and Humic Acids. PeerJ 2018, 2018, e5445. [Google Scholar] [CrossRef]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Cavagnaro, T.R. Chapter Two—A Meta-Analysis and Review of Plant-Growth Response to Humic Substances: Practical Implications for Agriculture. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 124, pp. 37–89. ISBN 0065-2113. [Google Scholar]
- Benito, P.; Ligorio, D.; Bellón, J.; Yenush, L.; Mulet, J.M. A Fast Method to Evaluate in a Combinatorial Manner the Synergistic Effect of Different Biostimulants for Promoting Growth or Tolerance against Abiotic Stress. Plant Methods 2022, 18, 111. [Google Scholar] [CrossRef]
- Benito, P.; Ligorio, D.; Bellón, J.; Yenush, L.; Mulet, J.M. Use of Yucca (Yucca schidigera) Extracts as Biostimulants to Promote Germination and Early Vigor and as Natural Fungicides. Plants 2023, 12, 274. [Google Scholar] [CrossRef] [PubMed]
- Saporta, R.; Bou, C.; Frías, V.; Mulet, J.M. A Method for a Fast Evaluation of the Biostimulant Potential of Different Natural Extracts for Promoting Growth or Tolerance against Abiotic Stress. Agronomy 2019, 9, 143. [Google Scholar] [CrossRef]
- Gisbert, C.; Timoneda, A.; Porcel, R.; Ros, R.; Mulet, J.M. Overexpression of Bvhb2, a Class 2 Non-Symbiotic Hemoglobin from Sugar Beet, Confers Drought-Induced Withering Resistance and Alters Iron Content in Tomato. Agronomy 2020, 10, 1754. [Google Scholar] [CrossRef]
- Ali, L.; Elbordiny, M. Response of Wheat Plants to Potassium Humate Application. J. Appl. Sci. Res. 2009, 5, 1202–1209. [Google Scholar]
- Said-Al Ahl, H.; Ayad, H.S.; Hendawy, S. Effect of Potassium Humate and Nitrogen Fertilizer on Herb and Essential Oil of Oregano under Different Irrigation Intervals. J. Appl. Sci. 2009, 2, 319–323. [Google Scholar]
- Ullah, A.; Ali, M.; Shahzad, K.; Ahmad, F.; Iqbal, S.; Habib Ur Rahman, M.; Ahmad, S.; Mazhar Iqbal, M.; Danish, S.; Fahad, S.; et al. Impact of Seed Dressing and Soil Application of Potassium Humate on Cotton Plants Productivity and Fiber Quality. Plants 2020, 9, 1444. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Al-Otaibi, H.H.; Parmar, A.; Ramadan, K.M.A.; Lobato, A.K.d.S.; El-Mogy, M.M. Application of Potassium Humate and Salicylic Acid to Mitigate Salinity Stress of Common Bean. Life 2023, 13, 448. [Google Scholar] [CrossRef]
- Alharbi, K.; Rashwan, E.; Hafez, E.; Omara, A.E.D.; Mohamed, H.H.; Alshaal, T. Potassium Humate and Plant Growth-Promoting Microbes Jointly Mitigate Water Deficit Stress in Soybean Cultivated in Salt-Affected Soil. Plants 2022, 11, 3016. [Google Scholar] [CrossRef]
- Ray, I.; Mridha, D.; Sarkar, J.; Joardar, M.; Das, A.; Chowdhury, N.R.; De, A.; Acharya, K.; Roychowdhury, T. Application of Potassium Humate to Reduce Arsenic Bioavailability and Toxicity in Rice Plants (Oryza sativa L.) during Its Course of Germination and Seedling Growth. Environ. Pollut. 2022, 313, 120066. [Google Scholar] [CrossRef]
- Mulet, J.M.; Llopis-Torregrosa, V.; Primo, C.; Marqués, M.C.; Yenush, L. Endocytic Regulation of Alkali Metal Transport Proteins in Mammals, Yeast and Plants. Curr. Genet. 2013, 59, 207–230. [Google Scholar] [CrossRef]
- LU, Y.A.N.; SHARKEY, T.D. The Importance of Maltose in Transitory Starch Breakdown. Plant Cell Environ. 2006, 29, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.-H.; Foster, K.J.; et al. Energy Costs of Salt Tolerance in Crop Plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, M.; Santelia, D. Starch as a Determinant of Plant Fitness under Abiotic Stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Streb, S.; Zeeman, S.C. Starch Metabolism in Arabidopsis. Arab. Book 2012, 10, e0160. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, Salt, and Temperature Stress-Induced Metabolic Rearrangements and Regulatory Networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- González-Cruz, J.; Pastenes, C. Water-Stress-Induced Thermotolerance of Photosynthesis in Bean (Phaseolus Vulgaris L.) Plants: The Possible Involvement of Lipid Composition and Xanthophyll Cycle Pigments. Environ. Exp. Bot. 2012, 77, 127–140. [Google Scholar] [CrossRef]
- Taïbi, K.; Ait Abderrahim, L.; Boussaid, M.; Bissoli, G.; Taïbi, F.; Achir, M.; Souana, K.; Mulet, J.M. Salt-Tolerance of Phaseolus Vulgaris L. Is a Function of the Potentiation Extent of Antioxidant Enzymes and the Expression Profiles of Polyamine Encoding Genes. South Afr. J. Bot. 2021, 140, 114–122. [Google Scholar] [CrossRef]
- Cuellar-Ortiz, S.M.; De La Paz Arrieta-Montiel, M.; Acosta-Gallegos, J.; Covarrubias, A.A. Relationship between Carbohydrate Partitioning and Drought Resistance in Common Bean. Plant Cell Environ. 2008, 31, 1399–1409. [Google Scholar] [CrossRef]
- Wu, W.; Zhong, Y.; Liu, Y.; Xu, R.; Zhang, X.; Liu, N.; Guo, D. A New Insight into the Biosynthesis, Structure, and Functionality of Waxy Maize Starch under Drought Stress. J. Sci. Food Agric. 2023, 103, 5270–5276. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. ISBN 0076-6879. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A Dyer Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1958, 37, 911–917. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water Stress Induced Changes in Concentrations of Proline and Total Soluble Sugars in Nodulated Alfalfa (Medicago sativa) Plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; De Mello, J.C.P. Application and Analysis of the Folin Ciocalteu Method for the Determination of the Total Phenolic Content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Ríos, G.; Cabedo, M.; Rull, B.; Yenush, L.; Serrano, R.; Mulet, J.M. Role of the Yeast Multidrug Transporter Qdr2 in Cation Homeostasis and the Oxidative Stress Response. FEMS Yeast Res. 2013, 13, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Caspar, T.; Lin, T.-P.; Kakefuda, G.; Benbow, L.; Preiss, J.; Somerville, C. Mutants of Arabidopsis with Altered Regulation of Starch Degradation. Plant Physiol. 1991, 95, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Caspar, T.; Huber, S.C.; Somerville, C. Alterations in Growth, Photosynthesis, and Respiration in a Starchless Mutant of Arabidopsis Thaliana (L.) Deficient in Chloroplast Phosphoglucomutase Activity. Plant Physiol. 1985, 79, 11–17. [Google Scholar] [CrossRef]
- Roessner, U.; Wagner, C.; Kopka, J.; Trethewey, R.N.; Willmitzer, L. Simultaneous Analysis of Metabolites in Potato Tuber by Gas Chromatography–Mass Spectrometry. Plant J. 2000, 23, 131–142. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Web-Based Inference of Biological Patterns, Functions and Pathways from Metabolomic Data Using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Common Up-Accumulated Metabolites (No KH60) | Conditions | Common Up-Accumulated Metabolites (In the Presence of KH60) |
|---|---|---|---|
| [LiCl vs. Man] | Sucrose | [LiCl vs. NaCl] | Histidine |
| [NaCl vs. Man] | Serine | [LiCl vs. Man] | Pyroglutamic acid |
| [LiCl vs. NaCl] [LiCl vs. Man] [NaCl vs. Man] | Phosphoric acid, Maltose | [LiCl vs. NaCl] [LiCl vs. Man] [NaCl vs. Man] | Glutamine |
| Conditions | Common Down-Accumulated Metabolites (No KH60) | Conditions | Common Down-Accumulated Metabolites (In the Presence of KH60) |
| [LiCl vs. NaCl] | Succinic acid, Asparagine | [LiCl vs. NaCl] | Glycine, Glycerol, Maleic acid, Malic acid |
| [LiCl vs. Man] | Lysine, Oxalic acid, Arginine | [LiCl vs. Man] | Succinic acid, Glutamic acid, Proline |
| [LiCl vs. NaCl] [LiCl vs. Man] [NaCl vs. Man] | Fumaric acid, Malic acid, Threonic acid, Aspartic acid, Glutamic acid, Tartaric acid, Fructose | [LiCl vs. NaCl] [LiCl vs. Man] [NaCl vs. Man] | Maltose, Threonic acid, Glucose, Fructose |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito, P.; Bellón, J.; Porcel, R.; Yenush, L.; Mulet, J.M. The Biostimulant, Potassium Humate Ameliorates Abiotic Stress in Arabidopsis thaliana by Increasing Starch Availability. Int. J. Mol. Sci. 2023, 24, 12140. https://doi.org/10.3390/ijms241512140
Benito P, Bellón J, Porcel R, Yenush L, Mulet JM. The Biostimulant, Potassium Humate Ameliorates Abiotic Stress in Arabidopsis thaliana by Increasing Starch Availability. International Journal of Molecular Sciences. 2023; 24(15):12140. https://doi.org/10.3390/ijms241512140
Chicago/Turabian StyleBenito, Patricia, Javier Bellón, Rosa Porcel, Lynne Yenush, and José M. Mulet. 2023. "The Biostimulant, Potassium Humate Ameliorates Abiotic Stress in Arabidopsis thaliana by Increasing Starch Availability" International Journal of Molecular Sciences 24, no. 15: 12140. https://doi.org/10.3390/ijms241512140
APA StyleBenito, P., Bellón, J., Porcel, R., Yenush, L., & Mulet, J. M. (2023). The Biostimulant, Potassium Humate Ameliorates Abiotic Stress in Arabidopsis thaliana by Increasing Starch Availability. International Journal of Molecular Sciences, 24(15), 12140. https://doi.org/10.3390/ijms241512140