
Deciphering the Action of Neuraminidase in Glioblastoma Models


 ,
, 
Abstract
1. Introduction
2. Results
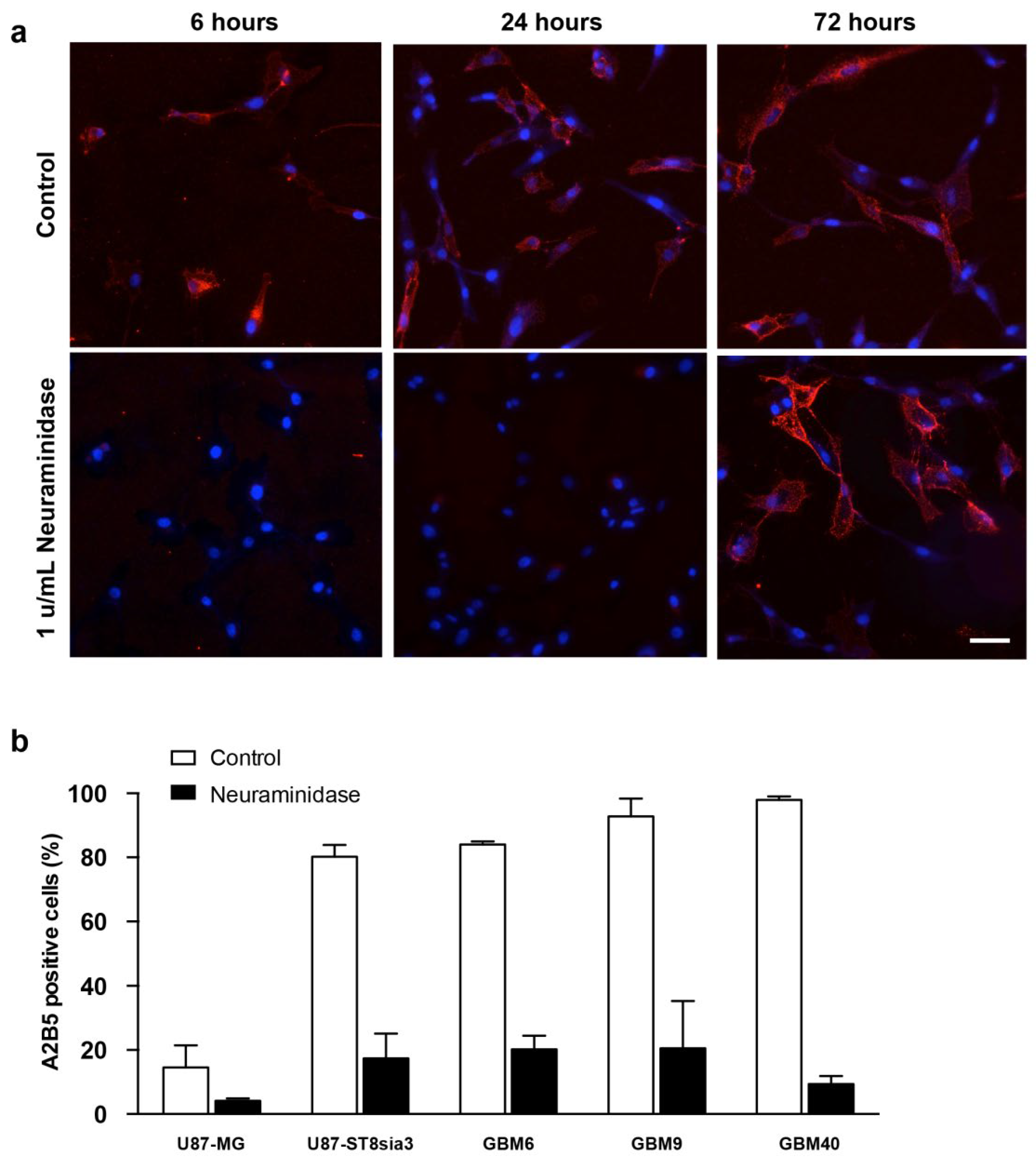
2.1. A2B5 Expression in Native Cell Lines and after Neuraminidase Administration
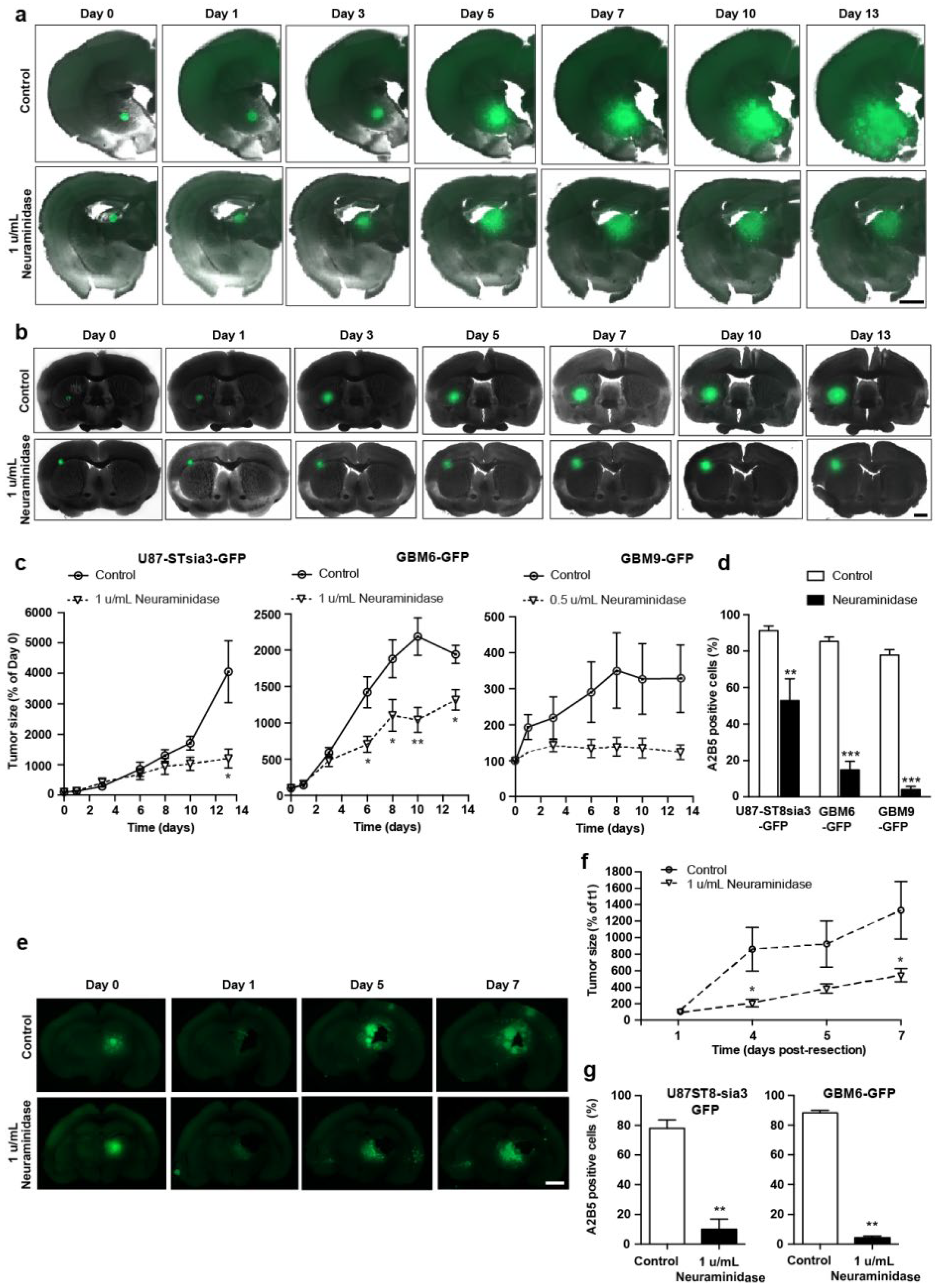
2.2. Neuraminidase Administration Strongly Decreased A2B5 Expression Tumor Size and Regrowth after Surgical Removal in a GBM Organotypic Slice Model
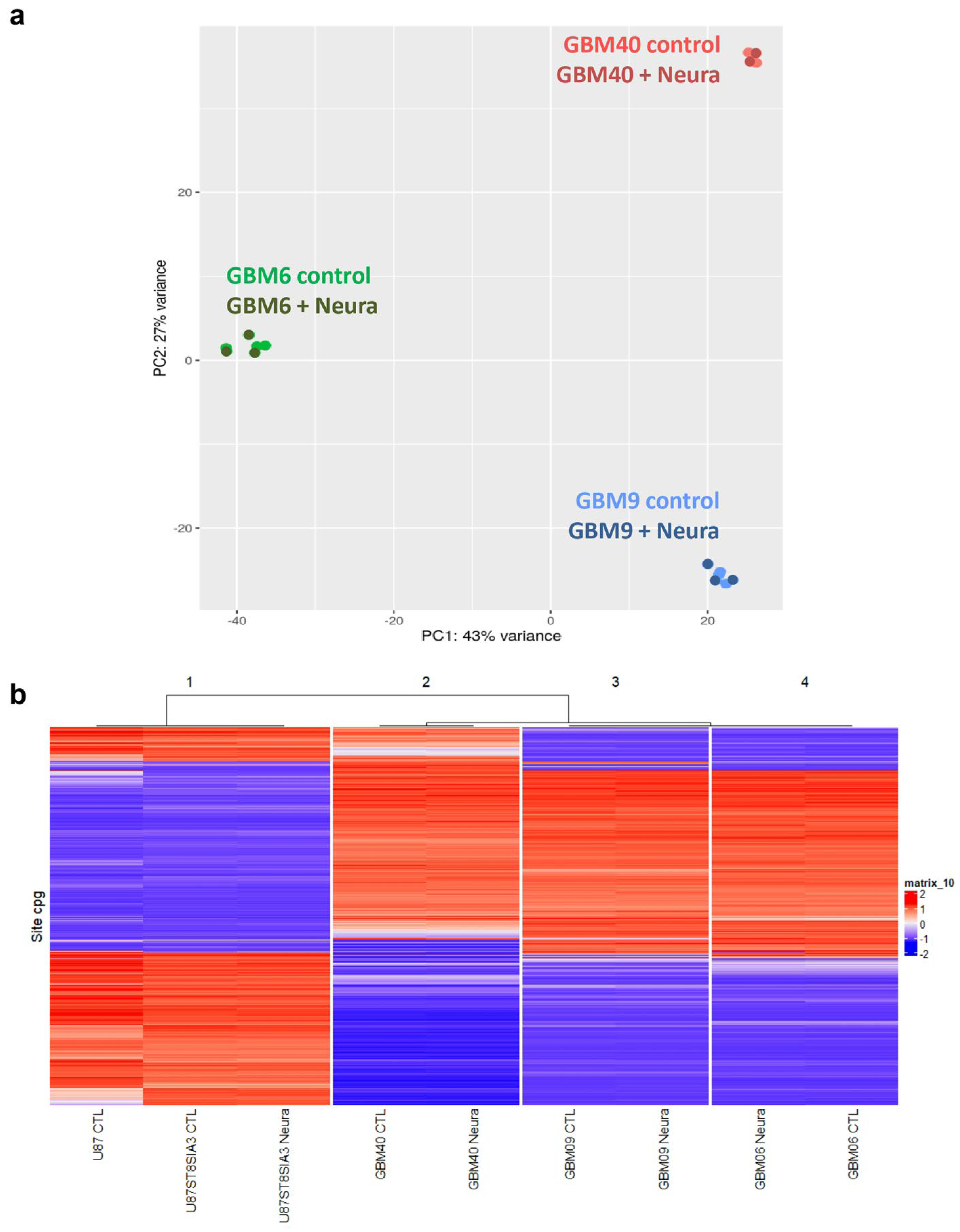
2.3. Neuraminidase Administration Did Not Induce a Distinct Transcriptomic Signature in CSC Lines
2.4. DNA Methylation-Profiling Demonstrated Strong Differences in Epigenetic Signature in U87 Cell Lines Versus CSC Lines but no Distinct Signature Was Induced by Neuraminidase
2.5. ST8Sia3 Overexpression in U87-MG Cell Line Induced a Specific Transcriptomic Signature
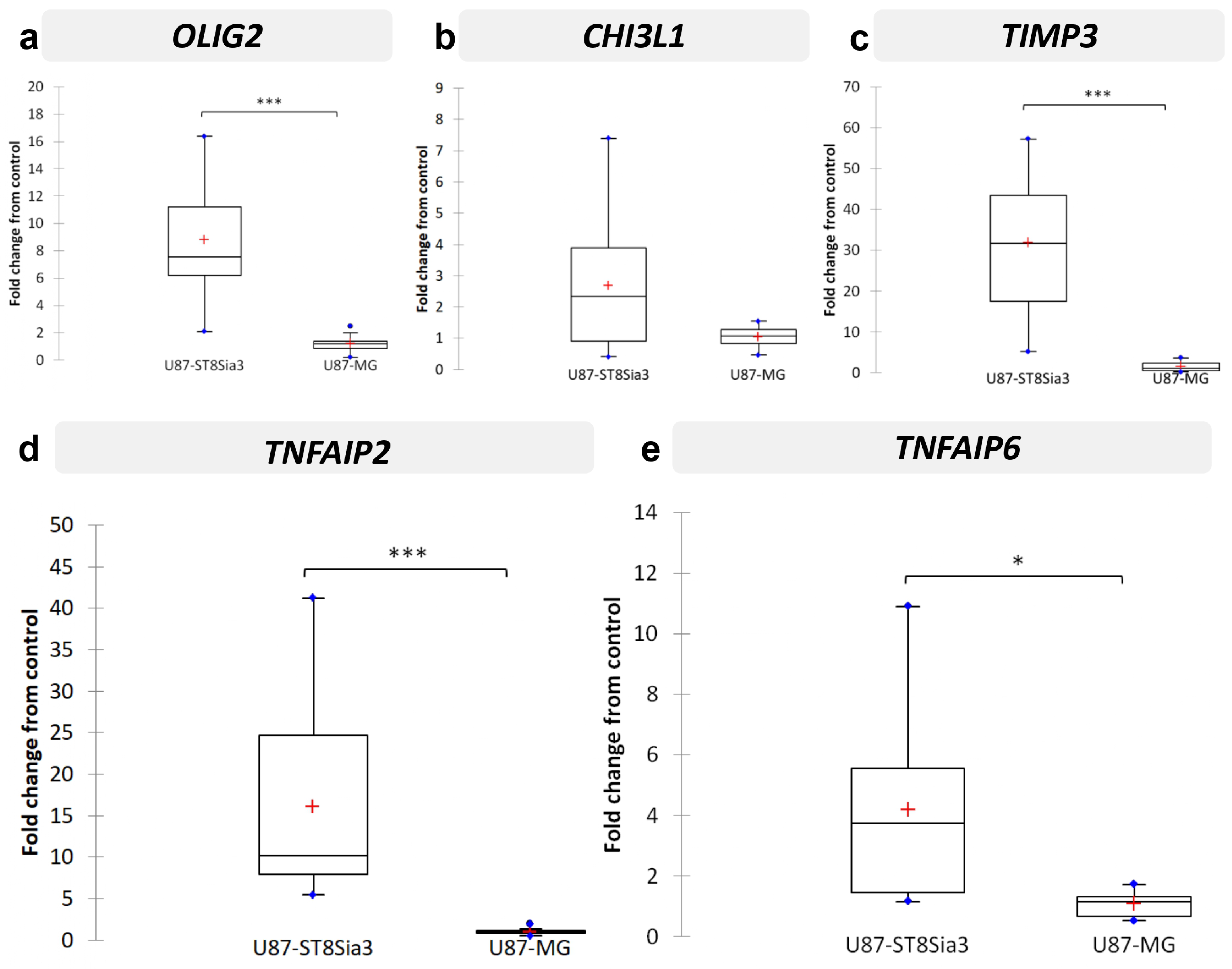
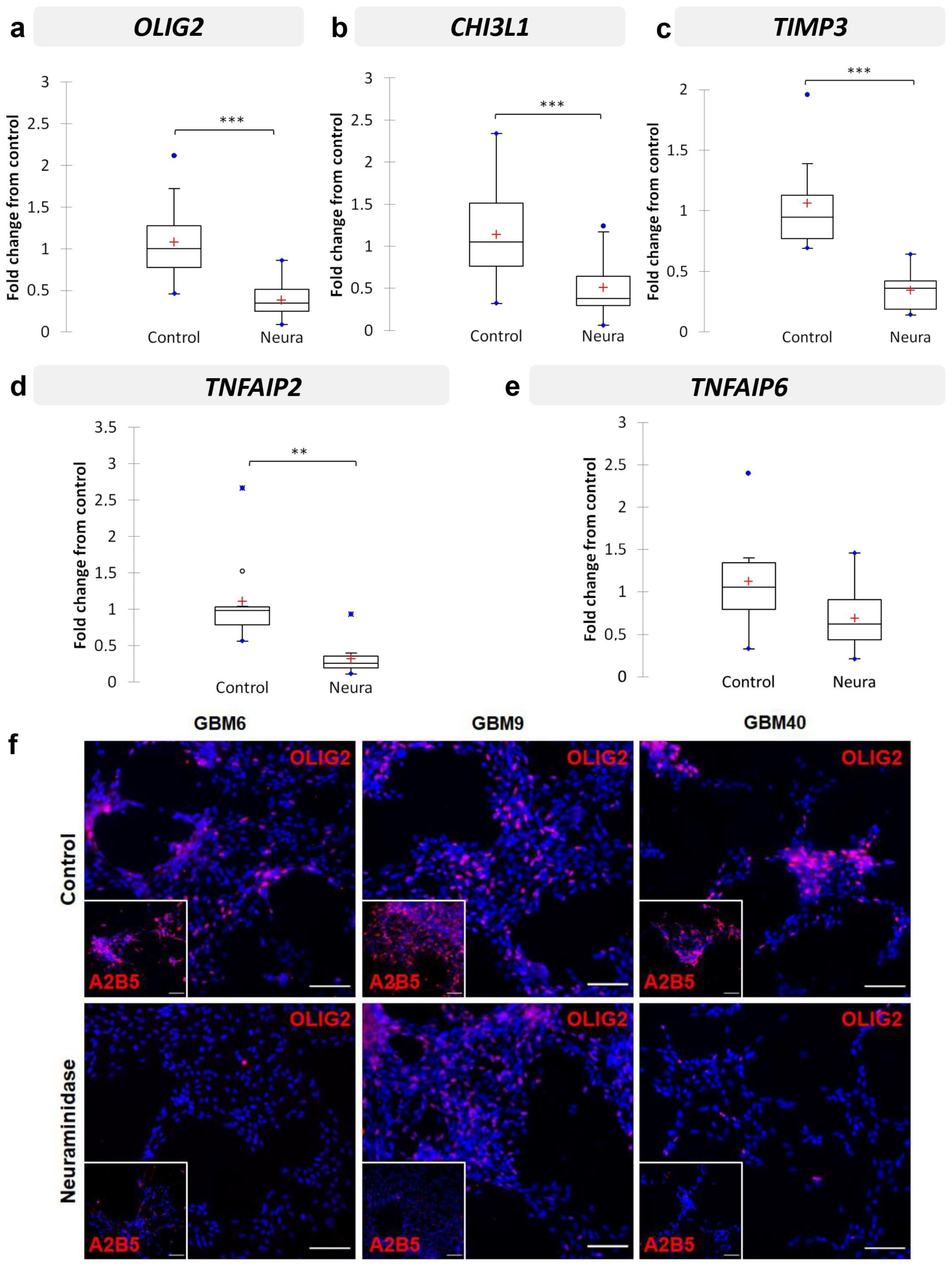
2.6. Neuraminidase Administration down Regulated OLIG2, CHI3L1, TIMP3, TNFAIP2 and TNFAIP6 Expression in CSC Lines
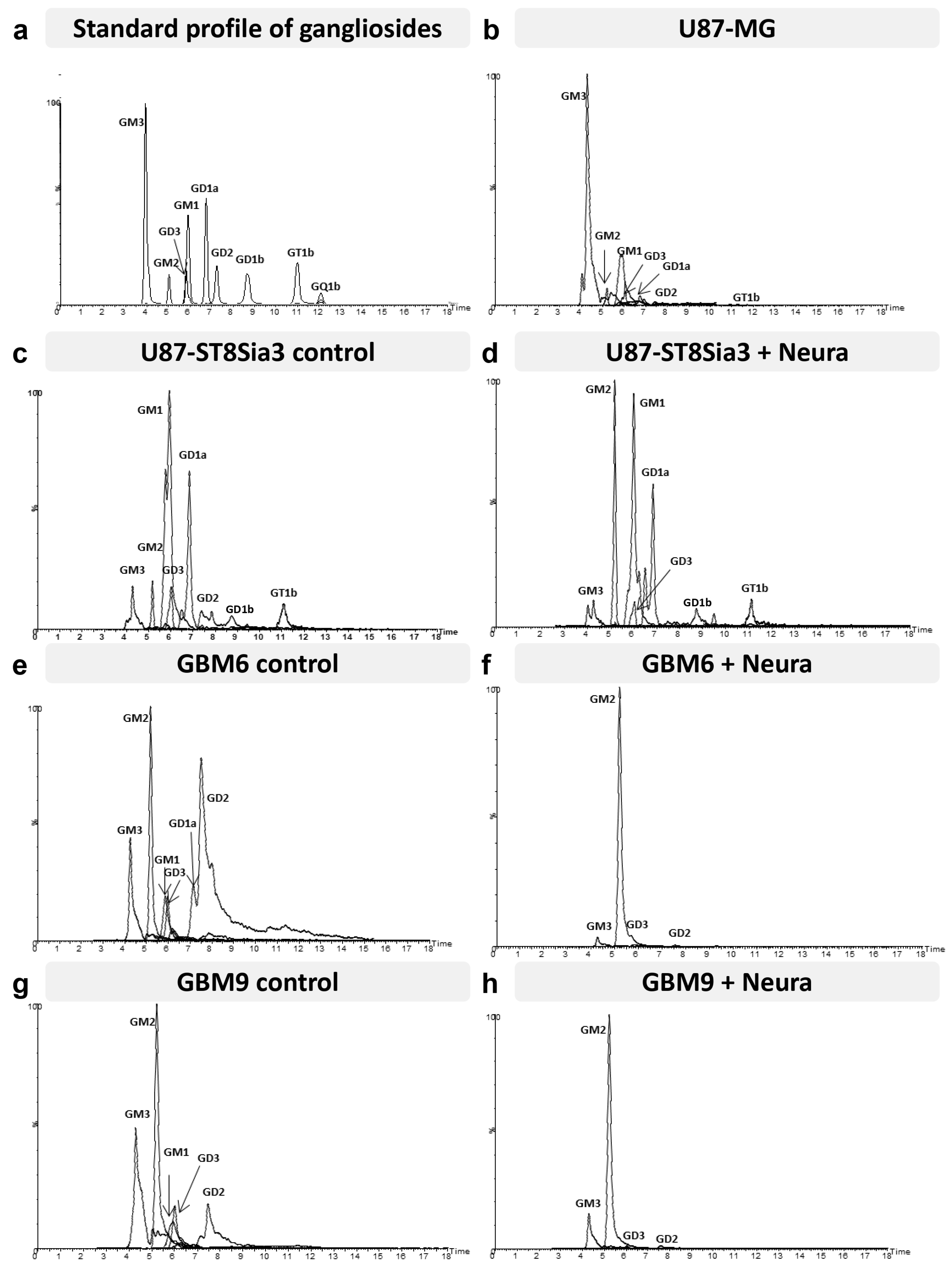
2.7. Neuraminidase Administration Induced Major Changes in Cell Surface Ganglioside Expression
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Culture Conditions and Reagents Used
4.2. Stainings
4.3. GBM Organotypic Slice Culture Model
4.4. DNA Methylation Analysis
4.4.1. DNA Extraction and Quantification
4.4.2. DNA Methylation Data Processing
4.5. Transcriptomic Analysis by RNA-Sequencing and Differential Gene Expression Analysis
4.5.1. Transcriptomic Analyses by RNA-Sequencing
4.5.2. Differential Gene Expression Analyses
4.6. Real-Time Quantitative PCR Analysis (RT-qPCR)
4.7. Ganglioside Expression in GBM Cell Lines
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Wen, P.Y.; Weller, M.; Lee, E.Q.; Alexander, B.M.; Barnholtz-Sloan, J.S.; Barthel, F.P.; Batchelor, T.T.; Bindra, R.S.; Chang, S.M.; Chiocca, E.A.; et al. Glioblastoma in adults: A Society for Neuro-Oncology (SNO) and European Society of Neuro-Oncology (EANO) consensus review on current management and future directions. Neuro Oncol. 2020, 22, 1073–1113. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Erhart, F.; Blauensteiner, B.; Zirkovits, G.; Printz, D.; Soukup, K.; Klingenbrunner, S.; Fischhuber, K.; Reitermaier, R.; Halfmann, A.; Lotsch, D.; et al. Gliomasphere marker combinatorics: Multidimensional flow cytometry detects CD44+/CD133+/ITGA6+/CD36+ signature. J. Cell. Mol. Med. 2019, 23, 281–292. [Google Scholar] [CrossRef]
- Bao, S.; Wu, Q.; Li, Z.; Sathornsumetee, S.; Wang, H.; McLendon, R.E.; Hjelmeland, A.B.; Rich, J.N. Targeting cancer stem cells through L1CAM suppresses glioma growth. Cancer Res. 2008, 68, 6043–6048. [Google Scholar] [CrossRef]
- Ogden, A.T.; Waziri, A.E.; Lochhead, R.A.; Fusco, D.; Lopez, K.; Ellis, J.A.; Kang, J.; Assanah, M.; McKhann, G.M.; Sisti, M.B.; et al. Identification of A2B5+CD133- tumor-initiating cells in adult human gliomas. Neurosurgery 2008, 62, 505–514. [Google Scholar] [CrossRef]
- Tchoghandjian, A.; Baeza, N.; Colin, C.; Cayre, M.; Metellus, P.; Beclin, C.; Ouafik, L.; Figarella-Branger, D. A2B5 cells from human glioblastoma have cancer stem cell properties. Brain Pathol. 2010, 20, 211–221. [Google Scholar] [CrossRef]
- Auvergne, R.M.; Sim, F.J.; Wang, S.; Chandler-Militello, D.; Burch, J.; Al Fanek, Y.; Davis, D.; Benraiss, A.; Walter, K.; Achanta, P.; et al. Transcriptional differences between normal and glioma-derived glial progenitor cells identify a core set of dysregulated genes. Cell Rep. 2013, 3, 2127–2141. [Google Scholar] [CrossRef]
- Eisenbarth, G.S.; Walsh, F.S.; Nirenberg, M. Monoclonal antibody to a plasma membrane antigen of neurons. Proc. Natl. Acad. Sci. USA 1979, 76, 4913–4917. [Google Scholar] [CrossRef]
- Kasai, N.; Yu, R.K. The monoclonal antibody A2B5 is specific to ganglioside GQ1c. Brain Res. 1983, 277, 155–158. [Google Scholar] [CrossRef]
- Saito, M.; Kitamura, H.; Sugiyama, K. The specificity of monoclonal antibody A2B5 to c-series gangliosides. J. Neurochem. 2001, 78, 64–74. [Google Scholar] [CrossRef]
- Dubois, C.; Manuguerra, J.C.; Hauttecoeur, B.; Maze, J. Monoclonal antibody A2B5, which detects cell surface antigens, binds to ganglioside GT3 (II3 (NeuAc)3LacCer) and to its 9-O-acetylated derivative. J. Biol. Chem. 1990, 265, 2797–2803. [Google Scholar] [CrossRef]
- Svennerholm, L. Chromatographic Separation of Human Brain Gangliosides. J. Neurochem. 1963, 10, 613–623. [Google Scholar] [CrossRef]
- Figarella-Branger, D.; Colin, C.; Baeza-Kallee, N.; Tchoghandjian, A. A2B5 Expression in Central Nervous System and Gliomas. Int. J. Mol. Sci. 2022, 23, 4670. [Google Scholar] [CrossRef]
- Crespo, P.M.; Demichelis, V.T.; Daniotti, J.L. Neobiosynthesis of glycosphingolipids by plasma membrane-associated glycosyltransferases. J. Biol. Chem. 2010, 285, 29179–29190. [Google Scholar] [CrossRef]
- Inoko, E.; Nishiura, Y.; Tanaka, H.; Takahashi, T.; Furukawa, K.; Kitajima, K.; Sato, C. Developmental stage-dependent expression of an alpha2,8-trisialic acid unit on glycoproteins in mouse brain. Glycobiology 2010, 20, 916–928. [Google Scholar] [CrossRef]
- Baeza-Kallee, N.; Berges, R.; Souberan, A.; Colin, C.; Denicolai, E.; Appay, R.; Tchoghandjian, A.; Figarella-Branger, D. Glycolipids Recognized by A2B5 Antibody Promote Proliferation, Migration, and Clonogenicity in Glioblastoma Cells. Cancers 2019, 11, 1267. [Google Scholar] [CrossRef]
- Tchoghandjian, A.; Baeza-Kallee, N.; Beclin, C.; Metellus, P.; Colin, C.; Ducray, F.; Adelaide, J.; Rougon, G.; Figarella-Branger, D. Cortical and subventricular zone glioblastoma-derived stem-like cells display different molecular profiles and differential in vitro and in vivo properties. Ann. Surg. Oncol. 2012, 19 (Suppl. S3), S608–S619. [Google Scholar] [CrossRef]
- Souberan, A.; Cappai, J.; Chocry, M.; Nuccio, C.; Raujol, J.; Colin, C.; Lafitte, D.; Kovacic, H.; Quillien, V.; Baeza-Kallee, N.; et al. Inhibitor of Apoptosis Proteins Determines Glioblastoma Stem-Like Cell Fate in an Oxygen-Dependent Manner. Stem Cells 2019, 37, 731–742. [Google Scholar] [CrossRef]
- Sibille, E.; Berdeaux, O.; Martine, L.; Bron, A.M.; Creuzot-Garcher, C.P.; He, Z.; Thuret, G.; Bretillon, L.; Masson, E.A. Ganglioside Profiling of the Human Retina: Comparison with Other Ocular Structures, Brain and Plasma Reveals Tissue Specificities. PLoS ONE 2016, 11, e0168794. [Google Scholar] [CrossRef] [PubMed]
- Masson, E.A.; Sibille, E.; Martine, L.; Chaux-Picquet, F.; Bretillon, L.; Berdeaux, O. Apprehending ganglioside diversity: A comprehensive methodological approach. J. Lipid Res. 2015, 56, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Yalamarty, S.S.K.; Filipczak, N.; Li, X.; Subhan, M.A.; Parveen, F.; Ataide, J.A.; Rajmalani, B.A.; Torchilin, V.P. Mechanisms of Resistance and Current Treatment Options for Glioblastoma Multiforme (GBM). Cancers 2023, 15, 2116. [Google Scholar] [CrossRef] [PubMed]
- Osuka, S.; Van Meir, E.G. Overcoming therapeutic resistance in glioblastoma: The way forward. J. Clin. Investig. 2017, 127, 415–426. [Google Scholar] [CrossRef]
- Fleurence, J.; Bahri, M.; Fougeray, S.; Faraj, S.; Vermeulen, S.; Pinault, E.; Geraldo, F.; Oliver, L.; Veziers, J.; Marquet, P.; et al. Impairing temozolomide resistance driven by glioma stem-like cells with adjuvant immunotherapy targeting O-acetyl GD2 ganglioside. Int. J. Cancer 2020, 146, 424–438. [Google Scholar] [CrossRef]
- Capper, D.; Jones, D.T.W.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chavez, L.; Reuss, D.E.; et al. DNA methylation-based classification of central nervous system tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef]
- Allen, M.; Bjerke, M.; Edlund, H.; Nelander, S.; Westermark, B. Origin of the U87MG glioma cell line: Good news and bad news. Sci. Transl. Med. 2016, 8, 354re3. [Google Scholar] [CrossRef]
- Bigner, D.D.; Bigner, S.H.; Ponten, J.; Westermark, B.; Mahaley, M.S.; Ruoslahti, E.; Herschman, H.; Eng, L.F.; Wikstrand, C.J. Heterogeneity of Genotypic and phenotypic characteristics of fifteen permanent cell lines derived from human gliomas. J. Neuropathol. Exp. Neurol. 1981, 40, 201–229. [Google Scholar] [CrossRef]
- Sauvageot, C.M.; Stiles, C.D. Molecular mechanisms controlling cortical gliogenesis. Curr. Opin. Neurobiol. 2002, 12, 244–249. [Google Scholar] [CrossRef]
- Ligon, K.L.; Huillard, E.; Mehta, S.; Kesari, S.; Liu, H.; Alberta, J.A.; Bachoo, R.M.; Kane, M.; Louis, D.N.; Depinho, R.A.; et al. Olig2-regulated lineage-restricted pathway controls replication competence in neural stem cells and malignant glioma. Neuron 2007, 53, 503–517. [Google Scholar] [CrossRef]
- Cheng, L.; Bao, S.; Rich, J.N. Potential therapeutic implications of cancer stem cells in glioblastoma. Biochem. Pharmacol. 2010, 80, 654–665. [Google Scholar] [CrossRef]
- Kupp, R.; Shtayer, L.; Tien, A.C.; Szeto, E.; Sanai, N.; Rowitch, D.H.; Mehta, S. Lineage-Restricted OLIG2-RTK Signaling Governs the Molecular Subtype of Glioma Stem-like Cells. Cell Rep. 2016, 16, 2838–2845. [Google Scholar] [CrossRef]
- Jia, L.; Zhou, Z.; Liang, H.; Wu, J.; Shi, P.; Li, F.; Wang, Z.; Wang, C.; Chen, W.; Zhang, H.; et al. KLF5 promotes breast cancer proliferation, migration and invasion in part by upregulating the transcription of TNFAIP2. Oncogene 2016, 35, 2040–2051. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, B. Downregulation of TNFAIP2 suppresses proliferation and metastasis in esophageal squamous cell carcinoma through activation of the Wnt/beta-catenin signaling pathway. Oncol. Rep. 2017, 37, 2920–2928. [Google Scholar] [CrossRef]
- Cheng, Z.; Wang, H.Z.; Li, X.; Wu, Z.; Han, Y.; Li, Y.; Chen, G.; Xie, X.; Huang, Y.; Du, Z.; et al. MicroRNA-184 inhibits cell proliferation and invasion, and specifically targets TNFAIP2 in Glioma. J. Exp. Clin. Cancer Res. 2015, 34, 27. [Google Scholar] [CrossRef]
- Tian, B.; Nowak, D.E.; Jamaluddin, M.; Wang, S.; Brasier, A.R. Identification of direct genomic targets downstream of the nuclear factor-kappaB transcription factor mediating tumor necrosis factor signaling. J. Biol. Chem. 2005, 280, 17435–17448. [Google Scholar] [CrossRef]
- Colin, C.; Baeza, N.; Bartoli, C.; Fina, F.; Eudes, N.; Nanni, I.; Martin, P.M.; Ouafik, L.; Figarella-Branger, D. Identification of genes differentially expressed in glioblastoma versus pilocytic astrocytoma using Suppression Subtractive Hybridization. Oncogene 2006, 25, 2818–2826. [Google Scholar] [CrossRef]
- Serres, E.; Debarbieux, F.; Stanchi, F.; Maggiorella, L.; Grall, D.; Turchi, L.; Burel-Vandenbos, F.; Figarella-Branger, D.; Virolle, T.; Rougon, G.; et al. Fibronectin expression in glioblastomas promotes cell cohesion, collective invasion of basement membrane in vitro and orthotopic tumor growth in mice. Oncogene 2014, 33, 3451–3462. [Google Scholar] [CrossRef]
- Lin, D.; Li, W.; Zhang, N.; Cai, M. Identification of TNFAIP6 as a hub gene associated with the progression of glioblastoma by weighted gene co-expression network analysis. IET Syst. Biol. 2022, 16, 145–156. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, X.; Xie, Y.; Liu, Y.; Ning, C.; Ai, Y.; Lv, C.; Wei, H.; Ge, X.; Yi, T.; et al. Identification of Neural Progenitor Cell-associated Chemoradiotherapy Resistance Gene Set (ARL4C, MSN, TNFAIP6) for Prognosis of Glioma. Curr. Pharm. Des. 2022, 28, 2189–2202. [Google Scholar] [CrossRef]
- Lan, G.; Yu, X.; Sun, X.; Li, W.; Zhao, Y.; Lan, J.; Wu, X.; Gao, R. Comprehensive analysis of the expression and prognosis for TNFAIPs in head and neck cancer. Sci. Rep. 2021, 11, 15696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xue, J.; Yang, H.; Zhou, T.; Zu, G. TNFAIP6 promotes invasion and metastasis of gastric cancer and indicates poor prognosis of patients. Tissue Cell 2021, 68, 101455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef] [PubMed]
- Recklies, A.D.; White, C.; Ling, H. The chitinase 3-like protein human cartilage glycoprotein 39 (HC-gp39) stimulates proliferation of human connective-tissue cells and activates both extracellular signal-regulated kinase- and protein kinase B-mediated signalling pathways. Biochem. J. 2002, 365 Pt 1, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Junker, N.; Johansen, J.S.; Hansen, L.T.; Lund, E.L.; Kristjansen, P.E. Regulation of YKL-40 expression during genotoxic or microenvironmental stress in human glioblastoma cells. Cancer Sci. 2005, 96, 183–190. [Google Scholar] [CrossRef]
- Zhao, T.; Zeng, J.; Xu, Y.; Su, Z.; Chong, Y.; Ling, T.; Xu, H.; Shi, H.; Zhu, M.; Mo, Q.; et al. Chitinase-3 like-protein-1 promotes glioma progression via the NF-kappaB signaling pathway and tumor microenvironment reprogramming. Theranostics 2022, 12, 6989–7008. [Google Scholar] [CrossRef]
- Jackson, H.W.; Defamie, V.; Waterhouse, P.; Khokha, R. TIMPs: Versatile extracellular regulators in cancer. Nat. Rev. Cancer 2017, 17, 38–53. [Google Scholar] [CrossRef]
- Hu, A.; Hong, F.; Li, D.; Jin, Y.; Kon, L.; Xu, Z.; He, H.; Xie, Q. Long non-coding RNA ROR recruits histone transmethylase MLL1 to up-regulate TIMP3 expression and promote breast cancer progression. J. Transl. Med. 2021, 19, 95. [Google Scholar] [CrossRef]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuna, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Chinot, O.L.; Wick, W.; Mason, W.; Henriksson, R.; Saran, F.; Nishikawa, R.; Carpentier, A.F.; Hoang-Xuan, K.; Kavan, P.; Cernea, D.; et al. Bevacizumab plus radiotherapy-temozolomide for newly diagnosed glioblastoma. N. Engl. J. Med. 2014, 370, 709–722. [Google Scholar] [CrossRef]
- Jiguet-Jiglaire, C.; Boissonneau, S.; Denicolai, E.; Hein, V.; Lasseur, R.; Garcia, J.; Romain, S.; Appay, R.; Graillon, T.; Mason, W.; et al. Plasmatic MMP9 released from tumor-infiltrating neutrophils is predictive for bevacizumab efficacy in glioblastoma patients: An AVAglio ancillary study. Acta Neuropathol. Commun. 2022, 10, 1. [Google Scholar] [CrossRef]
- Eckfeld, C.; Haussler, D.; Schoeps, B.; Hermann, C.D.; Kruger, A. Functional disparities within the TIMP family in cancer: Hints from molecular divergence. Cancer Metastasis Rev. 2019, 38, 469–481. [Google Scholar] [CrossRef]
- He, H.; Nilsson, C.L.; Emmett, M.R.; Ji, Y.; Marshall, A.G.; Kroes, R.A.; Moskal, J.R.; Colman, H.; Lang, F.F.; Conrad, C.A. Polar lipid remodeling and increased sulfatide expression are associated with the glioma therapeutic candidates, wild type p53 elevation and the topoisomerase-1 inhibitor, irinotecan. Glycoconj. J. 2010, 27, 27–38. [Google Scholar] [CrossRef]
- Kuhn, R.; Wiegandt, H. The Constitution of Gangliosides G-Ii, G-Iii and G-Iv. Z. Naturforschung B 1963, 18, 541–543. [Google Scholar] [CrossRef]
- Ledeen, R. New developments in the study of ganglioside structures. Chem. Phys. Lipids 1970, 5, 205–219. [Google Scholar] [CrossRef]
- Wikstrand, C.J.; Fredman, P.; Svennerholm, L.; Bigner, D.D. Detection of glioma-associated gangliosides GM2, GD2, GD3, 3′-isoLM1 3′,6′-isoLD1 in central nervous system tumors in vitro and in vivo using epitope-defined monoclonal antibodies. Prog. Brain Res. 1994, 101, 213–223. [Google Scholar] [CrossRef]
- Yeh, S.C.; Wang, P.Y.; Lou, Y.W.; Khoo, K.H.; Hsiao, M.; Hsu, T.L.; Wong, C.H. Glycolipid GD3 and GD3 synthase are key drivers for glioblastoma stem cells and tumorigenicity. Proc. Natl. Acad. Sci. USA 2016, 113, 5592–5597. [Google Scholar] [CrossRef]
- Fleurence, J.; Cochonneau, D.; Fougeray, S.; Oliver, L.; Geraldo, F.; Terme, M.; Dorvillius, M.; Loussouarn, D.; Vallette, F.; Paris, F.; et al. Targeting and killing glioblastoma with monoclonal antibody to O-acetyl GD2 ganglioside. Oncotarget 2016, 7, 41172–41185. [Google Scholar] [CrossRef]
- Mereiter, S.; Balmana, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef]
- Klaus, C.; Liao, H.; Allendorf, D.H.; Brown, G.C.; Neumann, H. Sialylation acts as a checkpoint for innate immune responses in the central nervous system. Glia 2021, 69, 1619–1636. [Google Scholar] [CrossRef]
- Bull, C.; Boltje, T.J.; Balneger, N.; Weischer, S.M.; Wassink, M.; van Gemst, J.J.; Bloemendal, V.R.; Boon, L.; van der Vlag, J.; Heise, T.; et al. Sialic Acid Blockade Suppresses Tumor Growth by Enhancing T-cell-Mediated Tumor Immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef]
- Appay, R.; Bielle, F.; Sievers, P.; Barets, D.; Fina, F.; Boutonnat, J.; Adam, C.; Gauchotte, G.; Godfraind, C.; Lhermitte, B.; et al. Rosette-forming glioneuronal tumours are midline, FGFR1-mutated tumours. Neuropathol. Appl. Neurobiol. 2022, 48, e12813. [Google Scholar] [CrossRef] [PubMed]
- Capper, D.; Stichel, D.; Sahm, F.; Jones, D.T.W.; Schrimpf, D.; Sill, M.; Schmid, S.; Hovestadt, V.; Reuss, D.E.; Koelsche, C.; et al. Practical implementation of DNA methylation and copy-number-based CNS tumor diagnostics: The Heidelberg experience. Acta Neuropathol. 2018, 136, 181–210. [Google Scholar] [CrossRef] [PubMed]
- Stichel, D.; Schrimpf, D.; Sievers, P.; Reinhardt, A.; Suwala, A.K.; Sill, M.; Reuss, D.E.; Korshunov, A.; Casalini, B.M.; Sommerkamp, A.C.; et al. Accurate calling of KIAA1549-BRAF fusions from DNA of human brain tumours using methylation array-based copy number and gene panel sequencing data. Neuropathol. Appl. Neurobiol. 2021, 47, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 16 July 2023).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Denicolai, E.; Baeza-Kallee, N.; Tchoghandjian, A.; Carre, M.; Colin, C.; Jiglaire, C.J.; Mercurio, S.; Beclin, C.; Figarella-Branger, D. Proscillaridin A is cytotoxic for glioblastoma cell lines and controls tumor xenograft growth in vivo. Oncotarget 2014, 5, 10934–10948. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Khoury, S.; Masson, E.; Sibille, E.; Cabaret, S.; Berdeaux, O. Rapid sample preparation for ganglioside analysis by liquid chromatography mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2020, 1137, 121956. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Gene Symbol | Fold Change | p Value | Description |
|---|---|---|---|---|
| Signal transduction | TNFAIP6 | 6.23 | 7.88 × 10−14 | TNF-Alpha-Induced Protein 6 |
| Cell signalling | PLEKHA1 | 5.58 | 4.97 × 10−12 | Pleckstrin Homology Domain Containing A1 |
| Receptors | WDFY1 | 5.28 | 1.76 × 10−11 | WD Repeat and FYVE Domain Containing 1 |
| RNF139 | 5.05 | 1.19 × 10−10 | Ring Finger Protein 139 | |
| CSNK2A1 | 4.99 | 1.21 × 10−10 | Casein Kinase 2 Alpha 1 | |
| IL1B | 4.52 | 2.54 × 10−09 | Interleukin 1 Alpha | |
| TOLLIP | 4.10 | 3.63 × 10−08 | Toll Interacting Protein | |
| ADGRE1 | 3.86 | 2.75 × 10−07 | Adhesion G Protein-Coupled Receptor E1 | |
| TMEM14C | 3.55 | 1.10 × 10−06 | Transmembrane Protein 14C | |
| CIR1 | 3.53 | 1.23 × 10−06 | Corepressor Interacting With RBPJ, 1 | |
| RASA2 | 3.52 | 1.56 × 10−06 | RAS P21 Protein Activator 2 | |
| ROS1 | 3.50 | 1.38 × 10−06 | ROS Proto-Oncogene 1, Receptor Tyrosine Kinase | |
| IGFBP4 | 3.16 | 8.39 × 10−06 | Insulin-Like Growth Factor Binding Protein 4 | |
| PTBP3 | 3.09 | 1.34 × 10−05 | Polypyrimidine Tract Binding Protein 3 | |
| TNFAIP2 | 3.01 | 1.90 × 10−05 | TNF-Alpha-Induced Protein 2 | |
| Transcription | MAFF | 5.28 | 3.87 × 10−11 | MAF BZIP Transcription Factor F |
| Translation | MRTO4 | 4.57 | 2.86 × 10−09 | MRT4 Homolog, Ribosome Maturation Factor |
| MED21 | 3.74 | 4.11 × 10−07 | Mediator Complex Subunit 21 | |
| CHRAC1 | 3.70 | 4.96 × 10−07 | Chromatin Accessibility Complex 1 | |
| POLR2J | 3.57 | 1.67 × 10−06 | RNA Polymerase II Subunit J | |
| RPS17L | 3.48 | 1.42 × 10−06 | Ribosomal Protein S17 | |
| ZNF721 | 3.42 | 7.16 × 10−06 | Zinc Finger Protein 721 | |
| KANSL3 | 3.29 | 4.50 × 10−06 | KAT8 Regulatory NSL Complex Subunit 3 | |
| RPS7 | 3.22 | 6.15 × 10−06 | Ribosomal Protein S7 | |
| TSC22D1 | 3.10 | 1.14 × 10−05 | TSC22 Domain Family Member 1 | |
| OLIG2 | 3.05 | 3.27 × 10−05 | Oligodendrocyte Transcription Factor 2 | |
| TBX3 | 3.04 | 1.85 × 10−05 | T-BOX 3 | |
| Cell cycle | G0S2 | 5.56 | 2.52 × 10−12 | G0/G1 Switch 2 |
| Mitosis | PCNP | 4.42 | 4.42 × 10−09 | PEST Proteolytic Signal Containing Nuclear Protein |
| INSIG1 | 4.24 | 1.22 × 10−08 | Insulin-Induced Gene 1 | |
| CSNK1G1 | 3.35 | 3.31 × 10−06 | Casein Kinase 1 Gamma 1 | |
| Cell structure | MAP1LC3B | 7.46 | 8.77 × 10−18 | Microtubule-Associated Protein 1 Light Chain 3 Beta |
| Motility | IGFN1 | 5.66 | 6.48 × 10−12 | Immunoglobulin-Like Fibronectin Type III Domain Containing 1 |
| PDLIM2 | 5.54 | 1.45 × 10−11 | PDZ And LIM Domain 2 | |
| FIBCD1 | 5.36 | 2.05 × 10−11 | Fibrinogen C Domain Containing 1 | |
| LASP1 | 4.90 | 2.03 × 10−10 | LIM And SH3 Protein 1 | |
| CHI3L1 | 4.71 | 6.31 × 10−10 | Chitinase 3 Like 1 | |
| TWF1 | 3.62 | 6.12 × 10−07 | Twinfilin Actin Binding Protein 1 | |
| P4HA1 | 3.27 | 4.46 × 10−06 | Prolyl 4-Hydroxylase Subunit Alpha 1 | |
| TIMP3 | 3.26 | 1.11 × 10−05 | TIMP Metallopeptidase Inhibitor 3 | |
| KRT8 | 3.04 | 1.76 × 10−05 | Keratin 8 | |
| Mitochondria | ISCA2 | 3.61 | 8.91 × 10−07 | Iron-Sulfur Cluster Assembly 2 |
| Metabolism | PRDX1 | 6.69 | 3.03 × 10−15 | Peroxiredoxin 1 |
| CHST1 | 5.64 | 7.10 × 10−12 | Carbohydrate Sulfotransferase 1 | |
| PDXK | 5.36 | 1.00 × 10−11 | Pyridoxal Kinase | |
| HSD11B1 | 4.88 | 8.27 × 10−10 | Hydroxysteroid 11-Beta Dehydrogenase 1 | |
| HS3ST2 | 4.65 | 2.33 × 10−09 | Heparan Sulfate-Glucosamine 3-Sulfotransferase 2 | |
| COLGALT1 | 4.49 | 2.71 × 10−09 | Collagen Beta(1-O)Galactosyltransferase 1 | |
| PSMB7 | 4.04 | 1.03 × 10−07 | Proteasome Subunit Beta 7 | |
| NHLRC2 | 3.97 | 8.57 × 10−08 | NHL Repeat Containing 2 | |
| SLC44A1 | 3.92 | 1.01 × 10−07 | Solute Carrier Family 44 Member 1 | |
| DNAJB2 | 3.90 | 1.20 × 10−07 | DnaJ Heat Shock Protein Family (Hsp40) Member B2 | |
| AOC2 | 3.69 | 1.37 × 10−06 | Amine Oxidase, Copper Containing 2 | |
| ACSS2 | 3.49 | 1.34 × 10−06 | Acyl-CoA Synthetase Short Chain Family Member 2 | |
| AKR1C1 | 3.44 | 1.92 × 10−06 | Aldo-Keto Reductase Family 1 Member C1 | |
| Transport | SCOC | 7.50 | 1.83 × 10−17 | Short Coiled-Coil Protein |
| SLC25A42 | 4.23 | 2.81 × 10−08 | Solute Carrier Family 25 Member 42 | |
| SLC39A1 | 3.66 | 4.84 × 10−07 | Solute Carrier Family 39 Member 1 | |
| RAB5C | 3.65 | 1.11 × 10−06 | RAB5C, Member RAS Oncogene Family | |
| Others | CCDC117 | 4.24 | 1.74 × 10−08 | Coiled-Coil Domain Containing 117 |
| KIAA1755 | 3.72 | 8.03 × 10−07 | Unknown | |
| ICOSLG | 3.48 | 1.91 × 10−06 | Inducible T-Cell Costimulator Ligand | |
| BST2 | 3.46 | 3.74 × 10−06 | Bone Marrow Stromal Cell Antigen 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baeza-Kallee, N.; Bergès, R.; Hein, V.; Cabaret, S.; Garcia, J.; Gros, A.; Tabouret, E.; Tchoghandjian, A.; Colin, C.; Figarella-Branger, D. Deciphering the Action of Neuraminidase in Glioblastoma Models. Int. J. Mol. Sci. 2023, 24, 11645. https://doi.org/10.3390/ijms241411645
Baeza-Kallee N, Bergès R, Hein V, Cabaret S, Garcia J, Gros A, Tabouret E, Tchoghandjian A, Colin C, Figarella-Branger D. Deciphering the Action of Neuraminidase in Glioblastoma Models. International Journal of Molecular Sciences. 2023; 24(14):11645. https://doi.org/10.3390/ijms241411645
Chicago/Turabian StyleBaeza-Kallee, Nathalie, Raphaël Bergès, Victoria Hein, Stéphanie Cabaret, Jeremy Garcia, Abigaëlle Gros, Emeline Tabouret, Aurélie Tchoghandjian, Carole Colin, and Dominique Figarella-Branger. 2023. "Deciphering the Action of Neuraminidase in Glioblastoma Models" International Journal of Molecular Sciences 24, no. 14: 11645. https://doi.org/10.3390/ijms241411645
APA StyleBaeza-Kallee, N., Bergès, R., Hein, V., Cabaret, S., Garcia, J., Gros, A., Tabouret, E., Tchoghandjian, A., Colin, C., & Figarella-Branger, D. (2023). Deciphering the Action of Neuraminidase in Glioblastoma Models. International Journal of Molecular Sciences, 24(14), 11645. https://doi.org/10.3390/ijms241411645