
The Effects of DDI1 on Inducing Differentiation in Ovine Preadipocytes via Oar-miR-432



Abstract
1. Introduction
2. Results
2.1. Oar-miR-432 Is Down-Regulated and Differentially Expressed in Sheep Tail Fat
2.2. Oar-miR-432 Inhibits Fat Deposition and Directly Targets DDI1
2.3. Oar-miR-432 Negatively Regulates the Expression of DDI1 in Ovine Preadipocytes
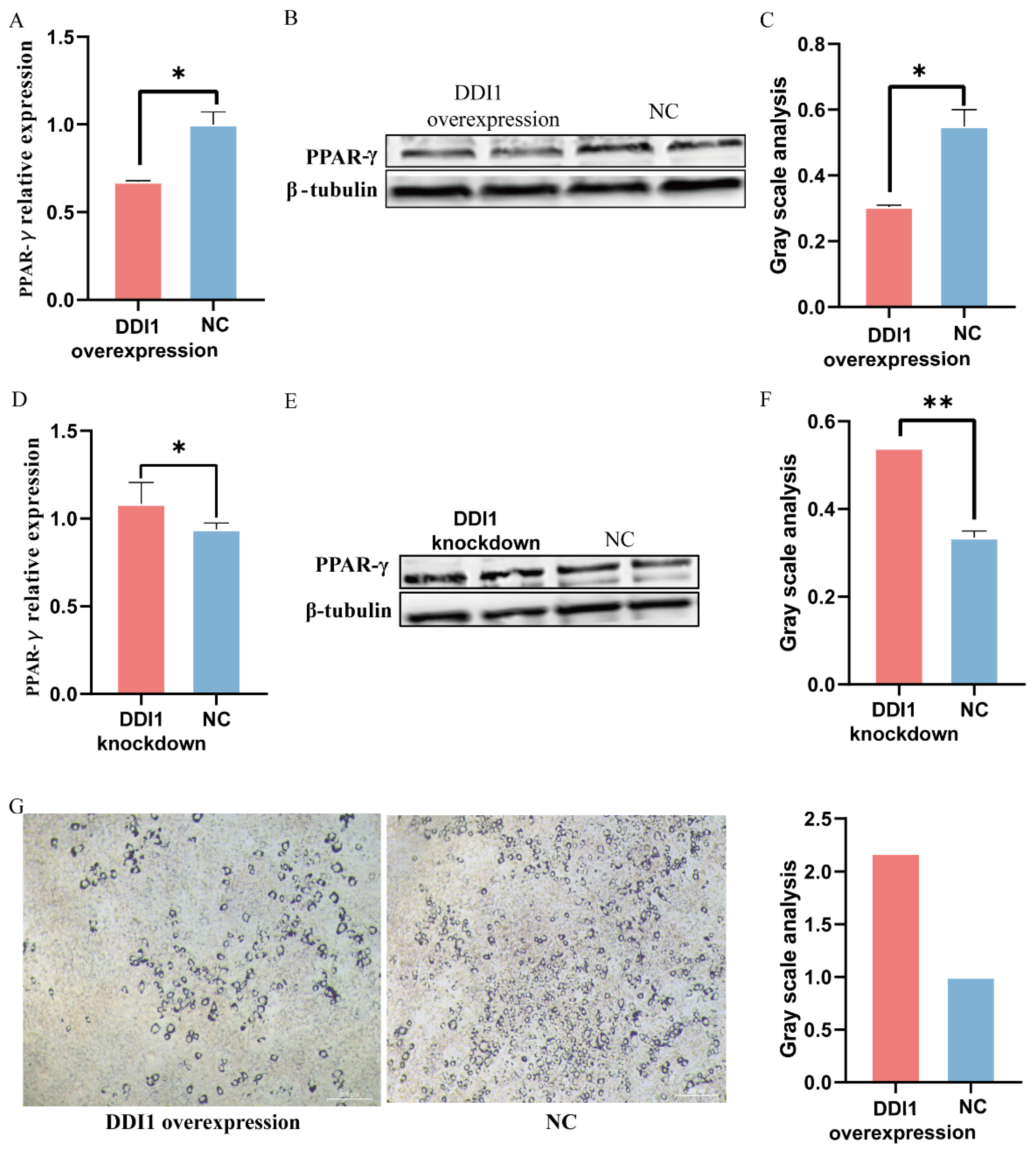
2.4. DDI1 Influences the Expression of Oar-miR-432 in Ovine Preadipocytes
2.5. DDI1 Inhibits Ovine Preadipocytes Inducing Differentiation
2.6. DDI1 Co-Expression Patterns with PDGFD
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Prediction of Oar-miR-432 Target Genes and Verification by a Dual Luciferase Reporter Assay
4.3. Cell Transfection
4.4. Oil Red O Staining
4.5. RT-qPCR
4.6. Western Blot (WB) Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Keim, S.M.; Guisto, J.A.; Sullivan, J.B., Jr. Environmental thermal stress. Ann. Agric. Environ. Med. AAEM 2002, 9, 1–15. [Google Scholar] [PubMed]
- Dzomba, E.F.; Van Der Nest, M.A.; Mthembu, J.N.T.; Soma, P.; Snyman, M.A.; Chimonyo, M.; Muchadeyi, F.C. Selection signature analysis and genome-wide divergence of South African Merino breeds from their founders. Front. Genet. 2022, 13, 932272. [Google Scholar] [CrossRef]
- Safdarian, M.; Zamiri, M.J.; Hashemi, M.; Noorolahi, H. Relationships of fat-tail dimensions with fat-tail weight and carcass characteristics at different slaughter weights of Torki-Ghashghaii sheep. Meat. Sci. 2008, 80, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Kalds, P.; Luo, Q.; Sun, K.; Zhou, S.; Chen, Y.; Wang, X. Trends towards revealing the genetic architecture of sheep tail patterning: Promising genes and investigatory pathways. Anim. Genet. 2021, 52, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Fei, X.; Li, T.; Lu, Z.; Chu, M.; Di, R.; He, X.; Wang, X.; Wang, Y.; Yuan, Z.; et al. Oar-miR-432 Regulates Fat Differentiation and Promotes the Expression of BMP2 in Ovine Preadipocytes. Front. Genet. 2022, 13, 844747. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Liu, E.; Liu, Z.; Kijas, J.W.; Zhu, C.; Hu, S.; Ma, X.; Zhang, L.; Du, L.; Wang, H.; et al. Selection signature analysis reveals genes associated with tail type in Chinese indigenous sheep. Anim. Genet. 2017, 48, 55–66. [Google Scholar] [CrossRef]
- Ingwersen, L.C.; Frank, M.; Naujokat, H.; Loger, K.; Bader, R.; Jonitz-Heincke, A. BMP-2 Long-Term Stimulation of Human Pre-Osteoblasts Induces Osteogenic Differentiation and Promotes Transdifferentiation and Bone Remodeling Processes. Int. J. Mol. Sci. 2022, 23, 3077. [Google Scholar] [CrossRef]
- Sirkis, R.; Gerst, J.E.; Fass, D. Ddi1, a eukaryotic protein with the retroviral protease fold. J. Mol. Biol. 2006, 364, 376–387. [Google Scholar] [CrossRef]
- Yip, M.C.J.; Bodnar, N.O.; Rapoport, T.A. Ddi1 is a ubiquitin-dependent protease. Proc. Natl. Acad. Sci. USA 2020, 117, 7776–7781. [Google Scholar] [CrossRef]
- Elu, N.; Osinalde, N.; Beaskoetxea, J.; Ramirez, J.; Lectez, B.; Aloria, K.; Rodriguez, J.A.; Arizmendi, J.M.; Mayor, U. Detailed Dissection of UBE3A-Mediated DDI1 Ubiquitination. Front. Physiol. 2019, 10, 534. [Google Scholar] [CrossRef]
- Ivantsiv, Y.; Kaplun, L.; Tzirkin-Goldin, R.; Shabek, N.; Raveh, D. Unique role for the UbL-UbA protein Ddi1 in turnover of SCFUfo1 complexes. Mol. Cell Biol. 2006, 26, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Olson, L.E.; Soriano, P. PDGFRbeta signaling regulates mural cell plasticity and inhibits fat development. Dev. Cell 2011, 20, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shao, J.; He, S.; Guo, Y.; Pan, X.; Wang, Y.; Nanaei, H.A.; Chen, L.; Li, R.; Xu, H.; et al. Allele-specific expression and splicing provide insight into the phenotypic differences between thin- and fat-tailed sheep breeds. J. Genet. Genom. 2022, 49, 583–586. [Google Scholar] [CrossRef]
- Dong, K.; Yang, M.; Han, J.; Ma, Q.; Han, J.; Song, Z.; Luosang, C.; Gorkhali, N.A.; Yang, B.; He, X.; et al. Genomic analysis of worldwide sheep breeds reveals PDGFD as a major target of fat-tail selection in sheep. BMC Genomics 2020, 21, 800. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Shen, M.; Xie, X.L.; Liu, G.J.; Xu, Y.X.; Lv, F.H.; Yang, H.; Yang, Y.L.; Liu, C.B.; et al. Whole-genome resequencing of wild and domestic sheep identifies genes associated with morphological and agronomic traits. Nat. Commun. 2020, 11, 2815. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Narayanan, R.; Schratt, G. miRNA regulation of social and anxiety-related behaviour. Cell Mol. Life Sci. 2020, 77, 4347–4364. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.W.; Lee, M.G.; Nam, K.H.; Park, J.H.; Oh, H.; Lee, J.; Han, J.; Yi, S.A.; Han, J.W. Reversine promotes browning of white adipocytes by suppressing miR-133a. J. Cell Physiol. 2019, 234, 3800–3813. [Google Scholar] [CrossRef]
- Trajkovski, M.; Ahmed, K.; Esau, C.C.; Stoffel, M. MyomiR-133 regulates brown fat differentiation through Prdm16. Nat. Cell Biol. 2012, 14, 1330–1335. [Google Scholar] [CrossRef]
- Giardina, S.; Hernández-Alonso, P.; Salas-Salvadó, J.; Rabassa-Soler, A.; Bulló, M. Modulation of Human Subcutaneous Adipose Tissue MicroRNA Profile Associated with Changes in Adiposity-Related Parameters. Mol. Nutr. Food Res. 2018, 62, 1700594. [Google Scholar] [CrossRef] [PubMed]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef]
- Gaddam, R.R.; Dhuri, K.; Kim, Y.R.; Jacobs, J.S.; Kumar, V.; Li, Q.; Irani, K.; Bahal, R.; Vikram, A. γ Peptide Nucleic Acid-Based miR-122 Inhibition Rescues Vascular Endothelial Dysfunction in Mice Fed a High-Fat Diet. J. Med. Chem. 2022, 65, 3332–3342. [Google Scholar] [CrossRef] [PubMed]
- Yaman, S.O.; Orem, A.; Yucesan, F.B.; Kural, B.V.; Orem, C. Evaluation of circulating miR-122, miR-30c and miR-33a levels and their association with lipids, lipoproteins in postprandial lipemia. Life Sci. 2021, 264, 118585. [Google Scholar] [CrossRef]
- Cornejo, P.J.; Vergoni, B.; Ohanna, M.; Angot, B.; Gonzalez, T.; Jager, J.; Tanti, J.F.; Cormont, M. The Stress-Responsive microRNA-34a Alters Insulin Signaling and Actions in Adipocytes through Induction of the Tyrosine Phosphatase PTP1B. Cells 2022, 11, 2581. [Google Scholar] [CrossRef]
- Alipour, M.R.; Naderi, R.; Alihemmati, A.; Sheervalilou, R.; Ghiasi, R. Swimming training attenuates pancreatic apoptosis through miR-34a/Sirtu in1/P53 Axis in high-fat diet and Streptozotocin-induced Type-2 diabetic rats. J. Diabetes Metab. Disord. 2020, 19, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Jin, M.; Wang, Y.; Li, T.; Lu, Z.; Yuan, Z.; Wang, H.; Lu, J.; Quan, K.; Di, R.; et al. Transcriptome reveals key microRNAs involved in fat deposition between different tail sheep breeds. PLoS ONE 2022, 17, e0264804. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Luo, Y.; Wang, J.; Hickford, J.G.H.; Zhou, H.; Hu, J.; Liu, X.; Li, S.; Shen, J.; Ke, N.; et al. MicroRNA-432 inhibits milk fat synthesis by targeting SCD and LPL in ovine mammary epithelial cells. Food Funct. 2021, 12, 9432–9442. [Google Scholar] [CrossRef]
- Romao, J.M.; Jin, W.; Dodson, M.V.; Hausman, G.J.; Moore, S.S.; Guan, L.L. MicroRNA regulation in mammalian adipogenesis. Exp. Biol. Med. 2011, 236, 997–1004. [Google Scholar] [CrossRef]
- Puri, V.; Ranjit, S.; Konda, S.; Nicoloro, S.M.; Straubhaar, J.; Chawla, A.; Chouinard, M.; Lin, C.; Burkart, A.; Corvera, S.; et al. Cidea is associated with lipid droplets and insulin sensitivity in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 7833–7838. [Google Scholar] [CrossRef]
- Agrawal, S.; Wang, M.; Klarqvist, M.D.R.; Smith, K.; Shin, J.; Dashti, H.; Diamant, N.; Choi, S.H.; Jurgens, S.J.; Ellinor, P.T.; et al. Inherited basis of visceral, abdominal subcutaneous and gluteofemoral fat depots. Nat. Commun. 2022, 13, 3771. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Zhu, Y.; Qiao, M.; Tang, X.; Zhao, W.; Xiao, F.; Liu, Y. The tomato DWD motif-containing protein DDI1 interacts with the CUL4-DDB1-based ubiquitin ligase and plays a pivotal role in abiotic stress responses. Biochem. Biophys Res. Commun. 2014, 450, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.Y.; Hu, G.S.; Tang, W.S.; Weng, L.; Zhang, Y.; Guo, H.; Yao, S.S.; Liu, S.Y.; Zhang, G.L.; et al. DDB1 binds histone reader BRWD3 to activate the transcriptional cascade in adipogenesis and promote onset of obesity. Cell Rep. 2021, 35, 109281. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, S.Y.; Hu, G.S.; Wang, H.Y.; Zhang, G.L.; Cen, X.; Xiang, S.T.; Peng, W.L.; Ye, H.B.; Zhao, T.J. DDB1 prepares brown adipocytes for cold-induced thermogenesis. Life Metab. 2022, 1, 39–53. [Google Scholar] [CrossRef]
- Groh, B.S.; Yan, F.; Smith, M.D.; Yu, Y.; Chen, X.; Xiong, Y. The antiobesity factor WDTC1 suppresses adipogenesis via the CRL4WDTC1 E3 ligase. EMBO Rep. 2016, 17, 638–647. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Moioli, B.; Ahbara, A.; Latairish, S.; Portolano, B.; Pilla, F.; Ciani, E. Genome-wide scan of fat-tail sheep identifies signals of selection for fat deposition and adaptation. Anim. Prod. Sci. 2018, 59, 835–848. [Google Scholar] [CrossRef]
- Zhang, G.; Xue, P.; Cui, S.; Yu, T.; Xiao, M.; Zhang, Q.; Cai, Y.; Jin, C.; Yang, J.; Wu, S.; et al. Different splicing isoforms of ERCC1 affect the expression of its overlapping genes CD3EAP and PPP1R13L, and indicate a potential application in non-small cell lung cancer treatment. Int. J. Oncol. 2018, 52, 2155–2165. [Google Scholar] [CrossRef]
- Wang, X.; Cao, X.; Dong, D.; Shen, X.; Cheng, J.; Jiang, R.; Yang, Z.; Peng, S.; Huang, Y.; Lan, X.; et al. Circular RNA TTN Acts As a miR-432 Sponge to Facilitate Proliferation and Differentiation of Myoblasts via the IGF2/PI3K/AKT Signaling Pathway. Mol. Ther. Nucleic Acids 2019, 18, 966–980. [Google Scholar] [CrossRef]
- Yau, W.W.; Singh, B.K.; Lesmana, R.; Zhou, J.; Sinha, R.A.; Wong, K.A.; Wu, Y.; Bay, B.H.; Sugii, S.; Sun, L.; et al. Thyroid hormone (T3) stimulates brown adipose tissue activation via mitochondrial biogenesis and MTOR-mediated mitophagy. Autophagy 2019, 15, 131–150. [Google Scholar] [CrossRef]
- Wakil, S.J.; Abu-Elheiga, L.A. Fatty acid metabolism: Target for metabolic syndrome. J. Lipid. Res. 2009, 50, S138–S143. [Google Scholar] [CrossRef]
- Tucci, S.; Behringer, S.; Spiekerkoetter, U. De novo fatty acid biosynthesis and elongation in very long-chain acyl-CoA dehydrogenase-deficient mice supplemented with odd or even medium-chain fatty acids. Febs. J. 2015, 282, 4242–4253. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Chen, W.J.; Zhang, J.W.; Xu, C.; Zeng, X.C.; Zhang, T.; Li, Y.; Wang, G.Y. Downregulation of miR-432 activates Wnt/β-catenin signaling and promotes human hepatocellular carcinoma proliferation. Oncotarget 2015, 6, 7866–7879. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.A.; Prasad, M.S.; Wong, M.; Charney, R.M.; Shelar, P.B.; Sandhu, N.; Hackland, J.O.S.; Hernandez, J.C.; Leung, A.W.; García-Castro, M.I. WNT/β-catenin modulates the axial identity of embryonic stem cell-derived human neural crest. Development 2019, 146, dev175604. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y. Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Front. Immunol. 2018, 9, 745. [Google Scholar] [CrossRef] [PubMed]
- Denton, N.F.; Eghleilib, M.; Al-Sharifi, S.; Todorčević, M.; Neville, M.J.; Loh, N.; Drakesmith, A.; Karpe, F.; Pinnick, K.E. Bone morphogenetic protein 2 is a depot-specific regulator of human adipogenesis. Int. J. Obes. 2019, 43, 2458–2468. [Google Scholar] [CrossRef]
- Yang, L.; Hao, W.; Wang, H.; Ren, W.; Yan, P.; Wei, S. BMP2 increases hyperplasia and hypertrophy of bovine subcutaneous preadipocytes via BMP/SMAD signaling. Vitr. Cell. Dev. Biol. Anim. 2022, 58, 210–219. [Google Scholar] [CrossRef]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Hailu, F.T.; Karimpour-Fard, A.; Toni, L.S.; Bristow, M.R.; Miyamoto, S.D.; Stauffer, B.L.; Sucharov, C.C. Integrated analysis of miRNA-mRNA interaction in pediatric dilated cardiomyopathy. Pediatr. Res. 2022, 92, 98–108. [Google Scholar] [CrossRef]
- Li, Q.; Lu, Z.; Jin, M.; Fei, X.; Quan, K.; Liu, Y.; Ma, L.; Chu, M.; Wang, H.; Wei, C. Verification and Analysis of Sheep Tail Type-Associated PDGF-D Gene Polymorphisms. Animals 2020, 10, 89. [Google Scholar] [CrossRef]
- Gallo-Oller, G.; Ordoñez, R.; Dotor, J. A new background subtraction method for Western blot densitometry band quantification through image analysis software. J. Immunol. Methods 2018, 457, 1–5. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Prime (5′ to 3′) | Tm | GenBank |
|---|---|---|---|
| DDI1-F | CACAGTCTATTGCGTGCGGA | 57.6 | XM_012095299.4 |
| DDI1-R | AGGAGCCCAGGGAGCAGTGA | 57.2 | |
| PDGFD-F | CTCAGGCGAGATGAGAGCAA | 55.9 | XM_004015965.5 |
| PDGFD-R | TCCTTGTGTCCACACCATCG | 57.7 | |
| PPAR-γ-F | ATAAAGCGTCAGGGTTCCAC | 56.3 | NM_001100921.1 |
| PPAR-γ-R | ATCCGACAGTTAAGATCACACC | 54.0 | |
| FABP4-F | AGCCACTTTCCTGGTAGCAA | 56.5 | NM_001114667.1 |
| FABP4-R | ACACACGCCTGCTCTTTCTTA | 56.5 | |
| ACACA-F | GTTTCGTCTCCACCACCGAA | 57.7 | NM_001009256.1 |
| ACACA-R | CTGACTCGTCAGTGAGACCG | 57.8 | |
| β-actin-F | CCAACCGTGAGAAGATGACC | 56.5 | NM_001009784.3 |
| β-actin-R | CCCGAGGCGTACAGGGACAG | 57.1 | |
| oar-miR-432-RT | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCCACCC | - | NR_107875.1 |
| oar-miR-432-F | CGCGTCTTGGAGTAGGTCATT | 58.5 | |
| oar-5s-RT | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGCCTA | - | XR_003590556.1 |
| oar-5s-F | TGGGAATACCGGGTGCTG | 57.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, M.; Yuan, Z.; Li, T.; Wang, H.; Wei, C. The Effects of DDI1 on Inducing Differentiation in Ovine Preadipocytes via Oar-miR-432. Int. J. Mol. Sci. 2023, 24, 11567. https://doi.org/10.3390/ijms241411567
Jin M, Yuan Z, Li T, Wang H, Wei C. The Effects of DDI1 on Inducing Differentiation in Ovine Preadipocytes via Oar-miR-432. International Journal of Molecular Sciences. 2023; 24(14):11567. https://doi.org/10.3390/ijms241411567
Chicago/Turabian StyleJin, Meilin, Zehu Yuan, Taotao Li, Huihua Wang, and Caihong Wei. 2023. "The Effects of DDI1 on Inducing Differentiation in Ovine Preadipocytes via Oar-miR-432" International Journal of Molecular Sciences 24, no. 14: 11567. https://doi.org/10.3390/ijms241411567
APA StyleJin, M., Yuan, Z., Li, T., Wang, H., & Wei, C. (2023). The Effects of DDI1 on Inducing Differentiation in Ovine Preadipocytes via Oar-miR-432. International Journal of Molecular Sciences, 24(14), 11567. https://doi.org/10.3390/ijms241411567