

Serpin Family A Member 1 Is Prognostic and Involved in Immunological Regulation in Human Cancers
Abstract
1. Introduction
2. Results
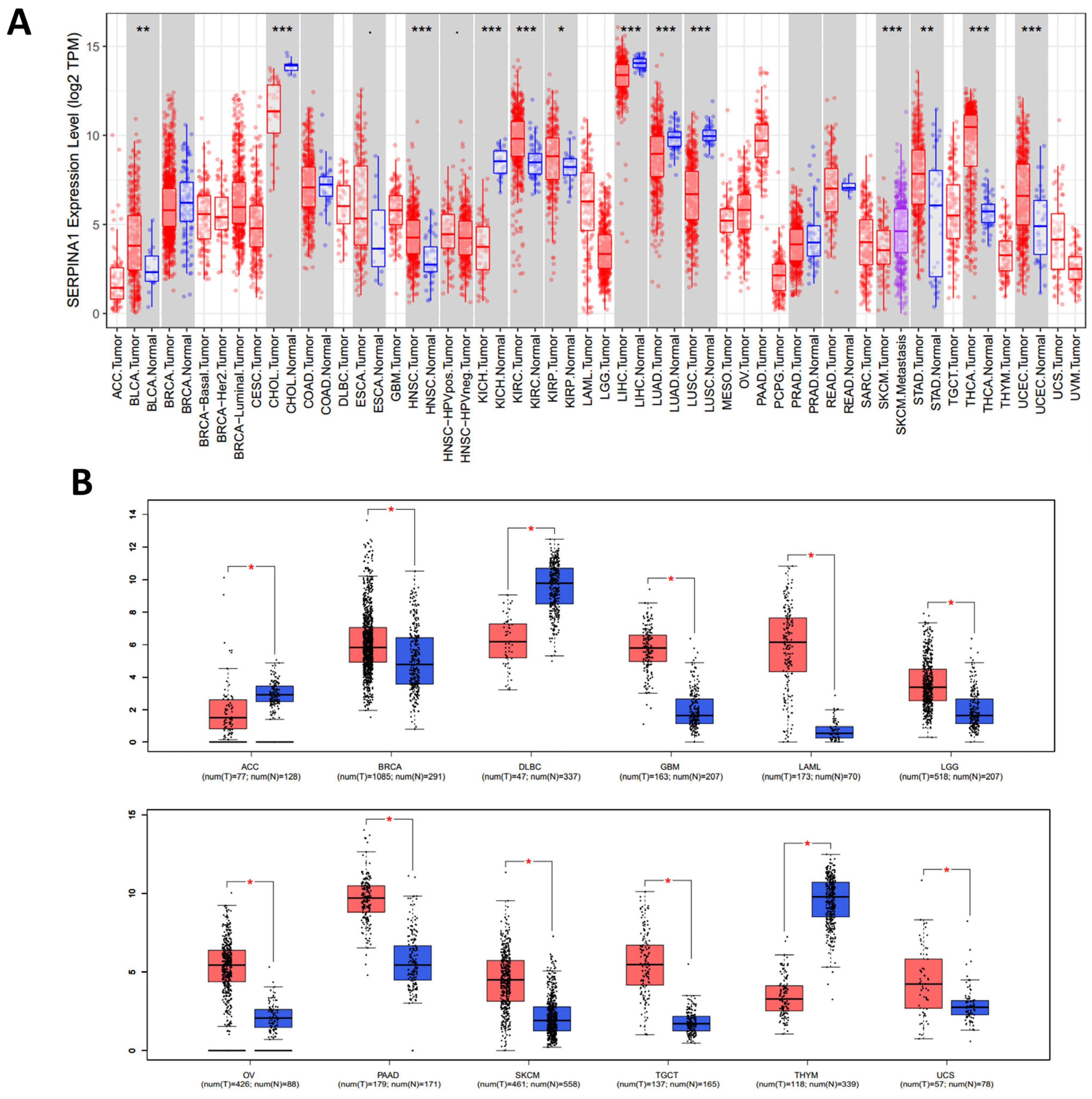
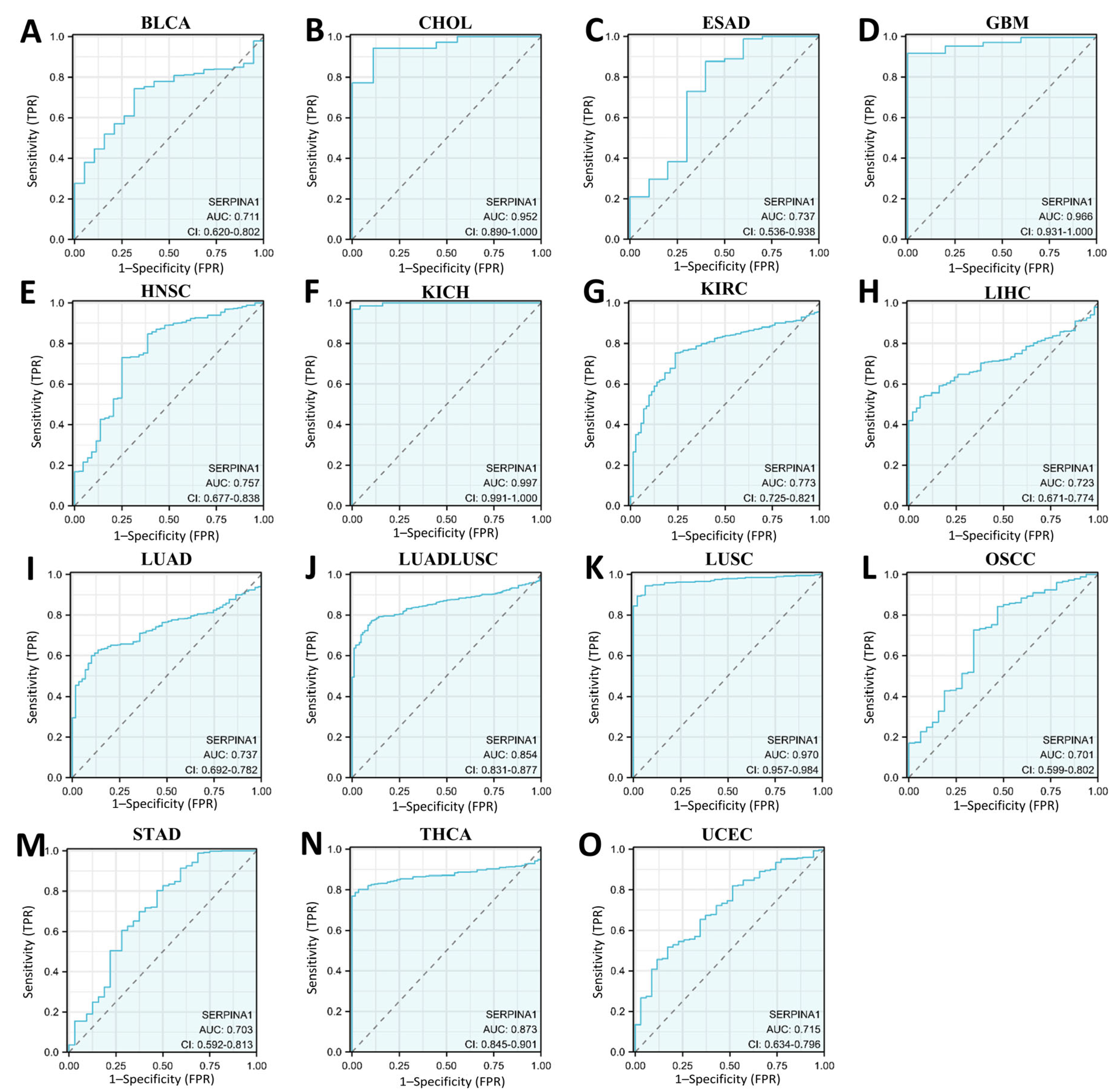
2.1. Pan-Cancer Differential Expressions of SERPINA1 between Tumor and Normal Tissues
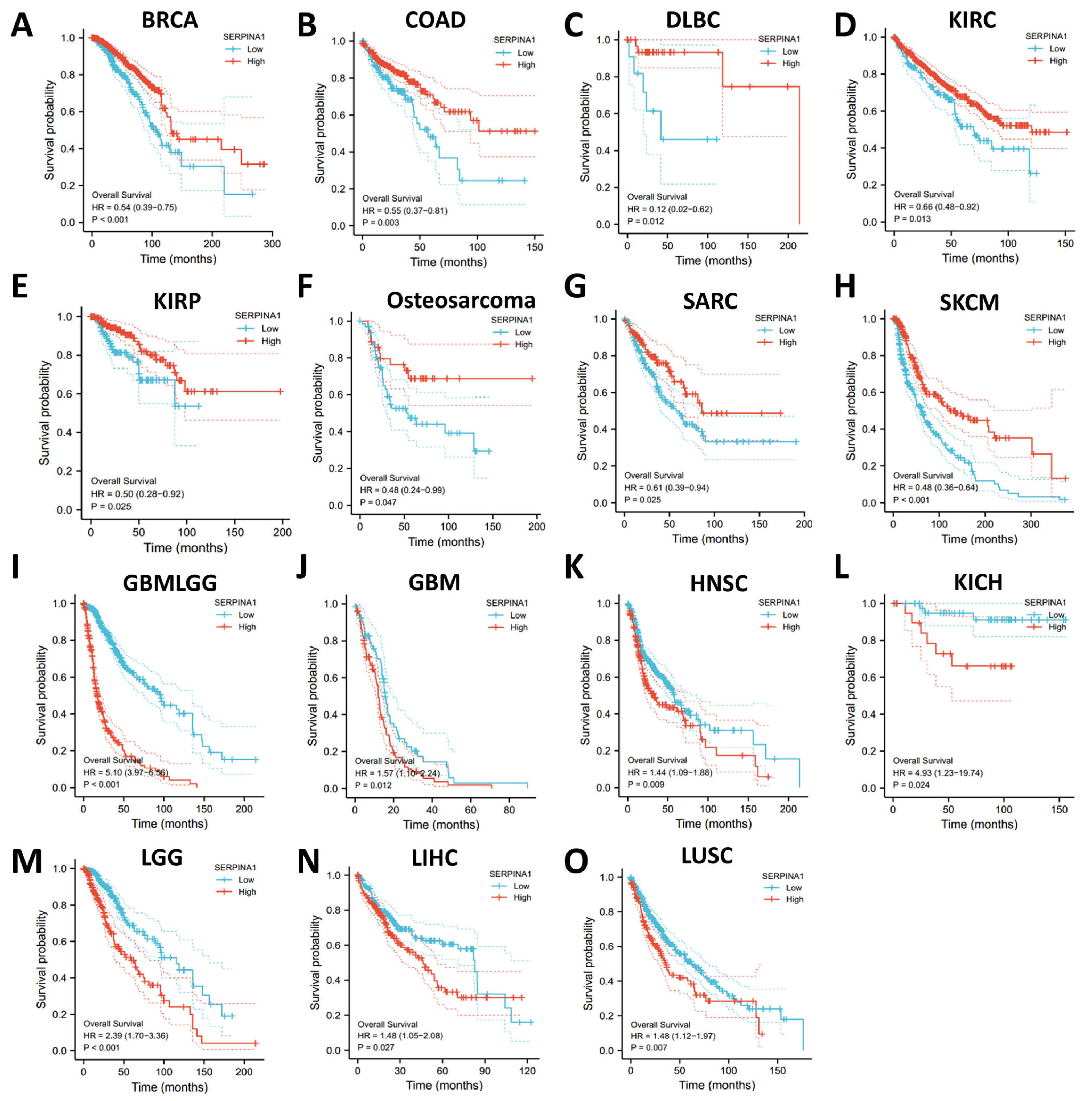
2.2. Dual Prognostic Role of SERPINA1 in Human Cancers
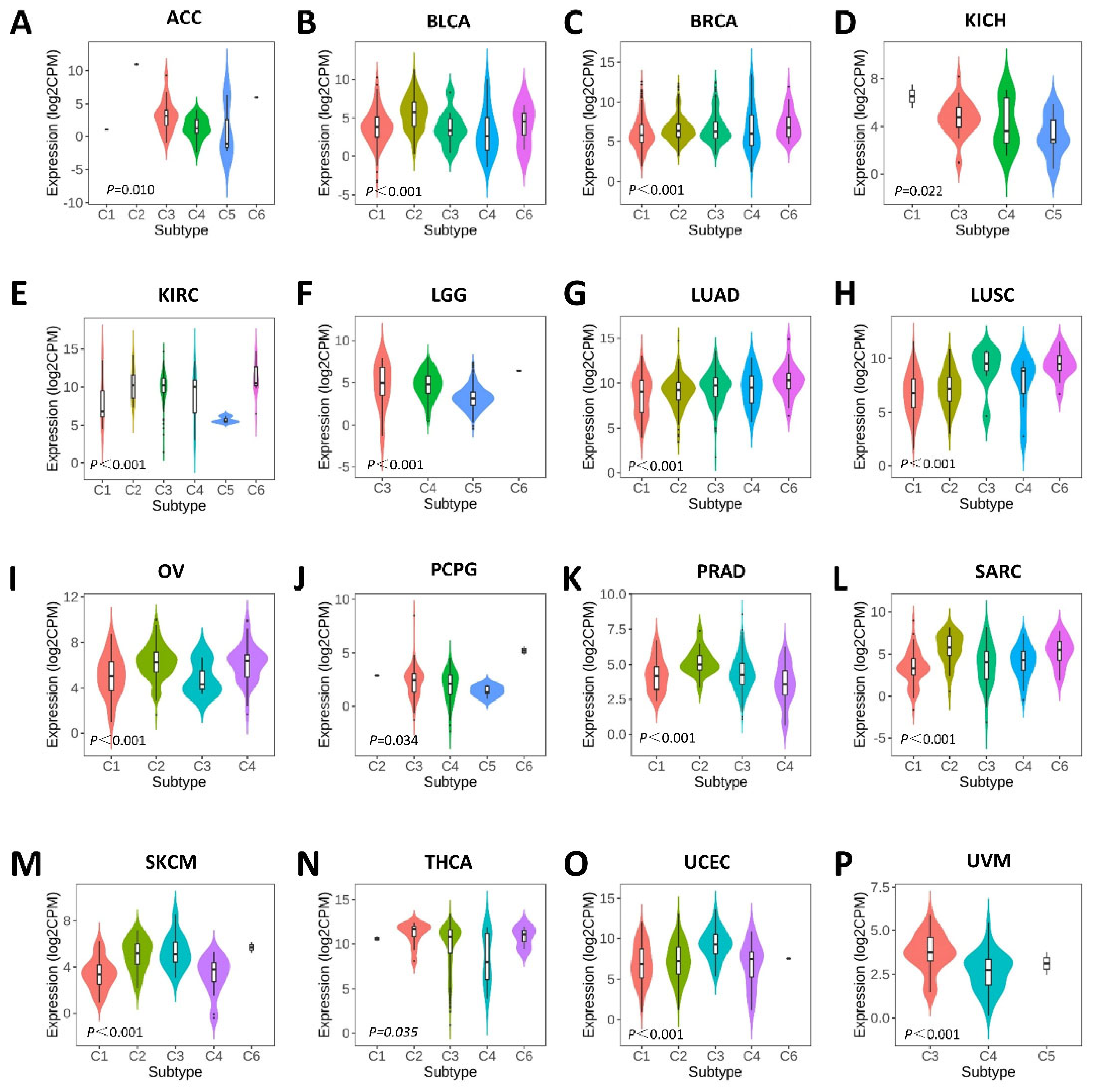
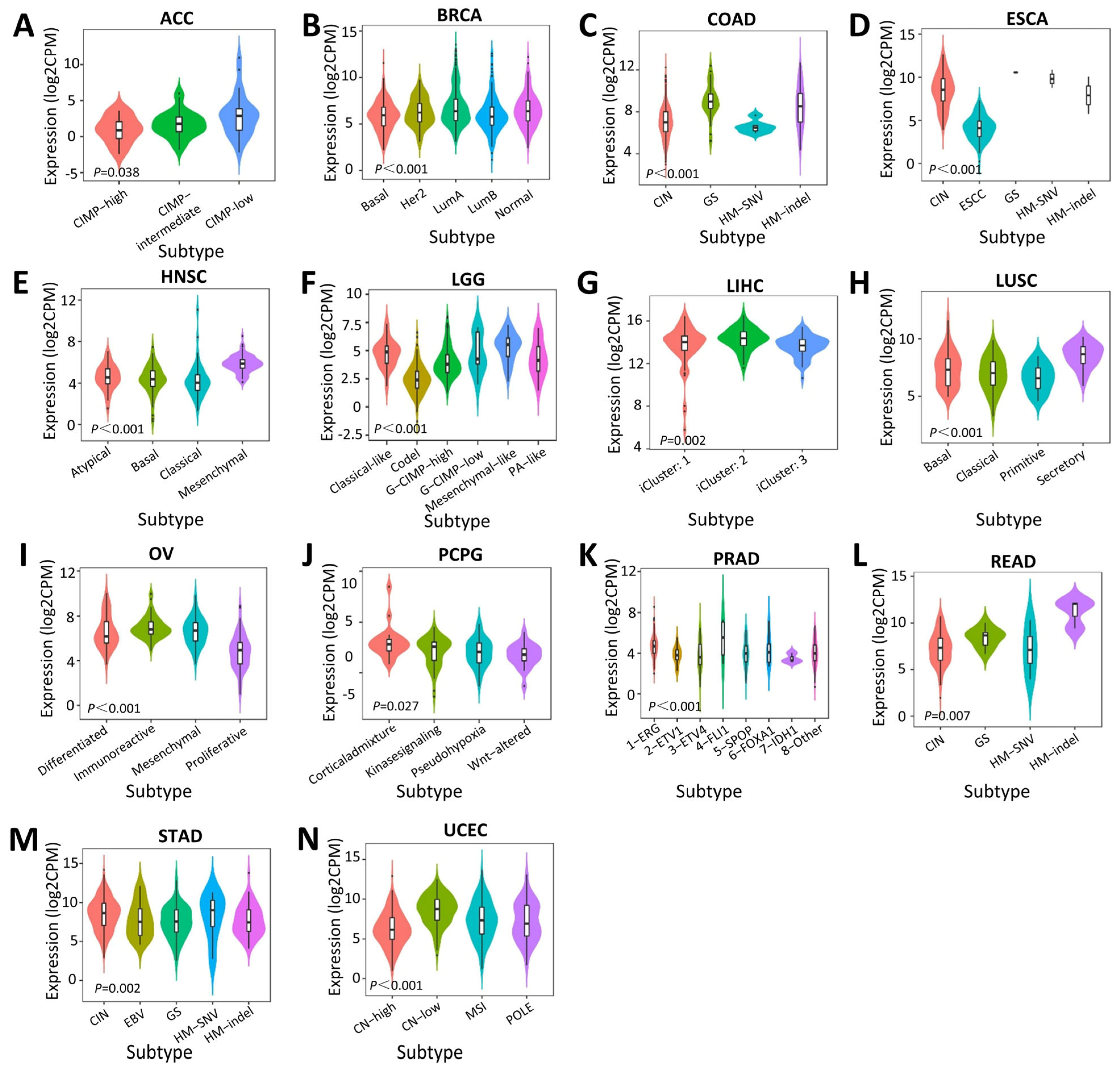
2.3. Associations between SERPINA1 and Immune and Molecular Subtypes
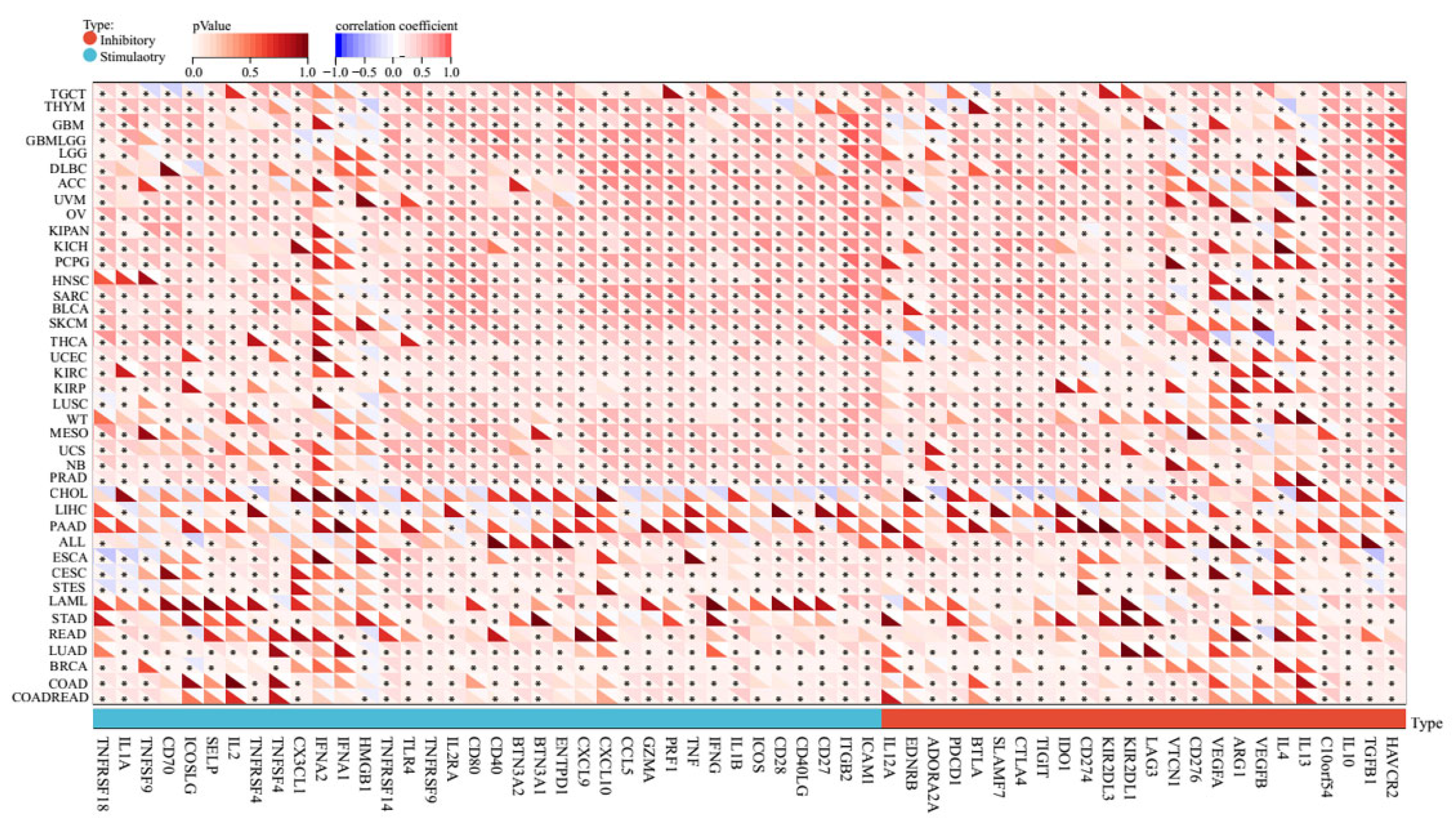
2.4. Pan-Cancer Relationships between AERPINA1 and Immune Checkpoint (ICP) Genes
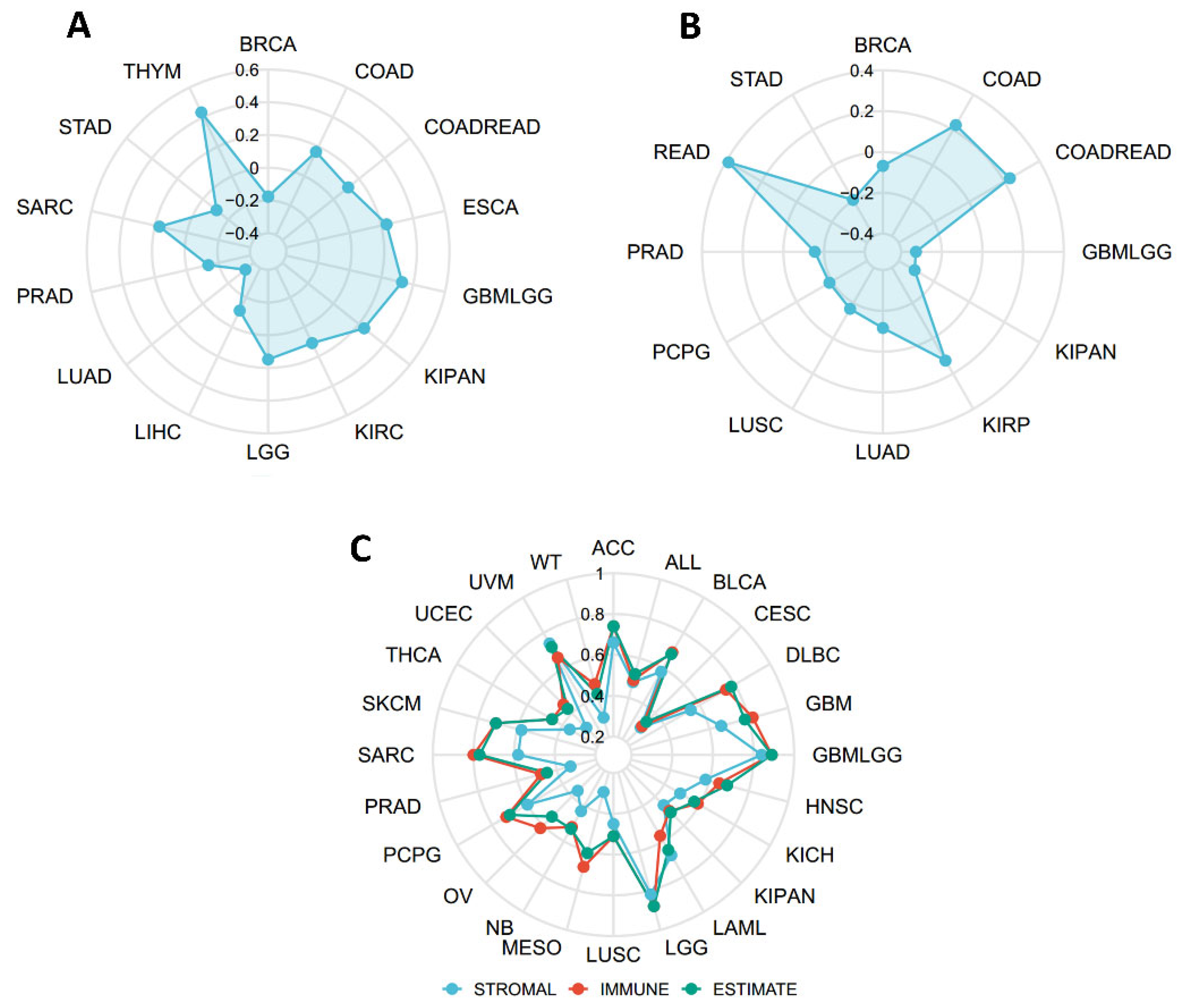
2.5. Connections between SERPINA1 and Tumor Mutational Burden (TMB), Microsatellite Instability (MSI), and Estimation of Stromal and Immune Cells in Malignant Tumor Tissues Using Expression Data (ESTIMATE)
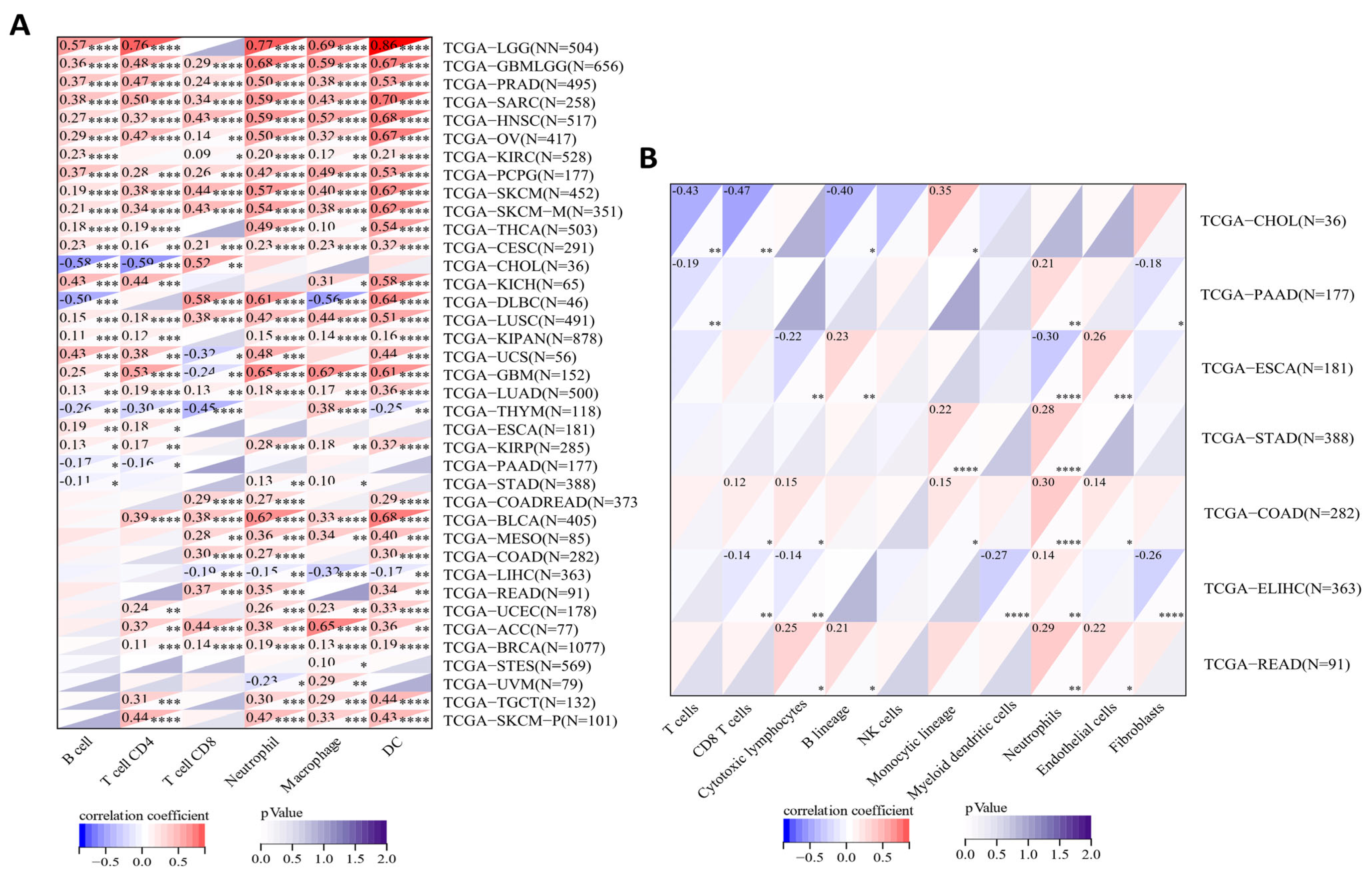
2.6. Correlations between SERPINA1 and Immune Cell Infiltration in the TME
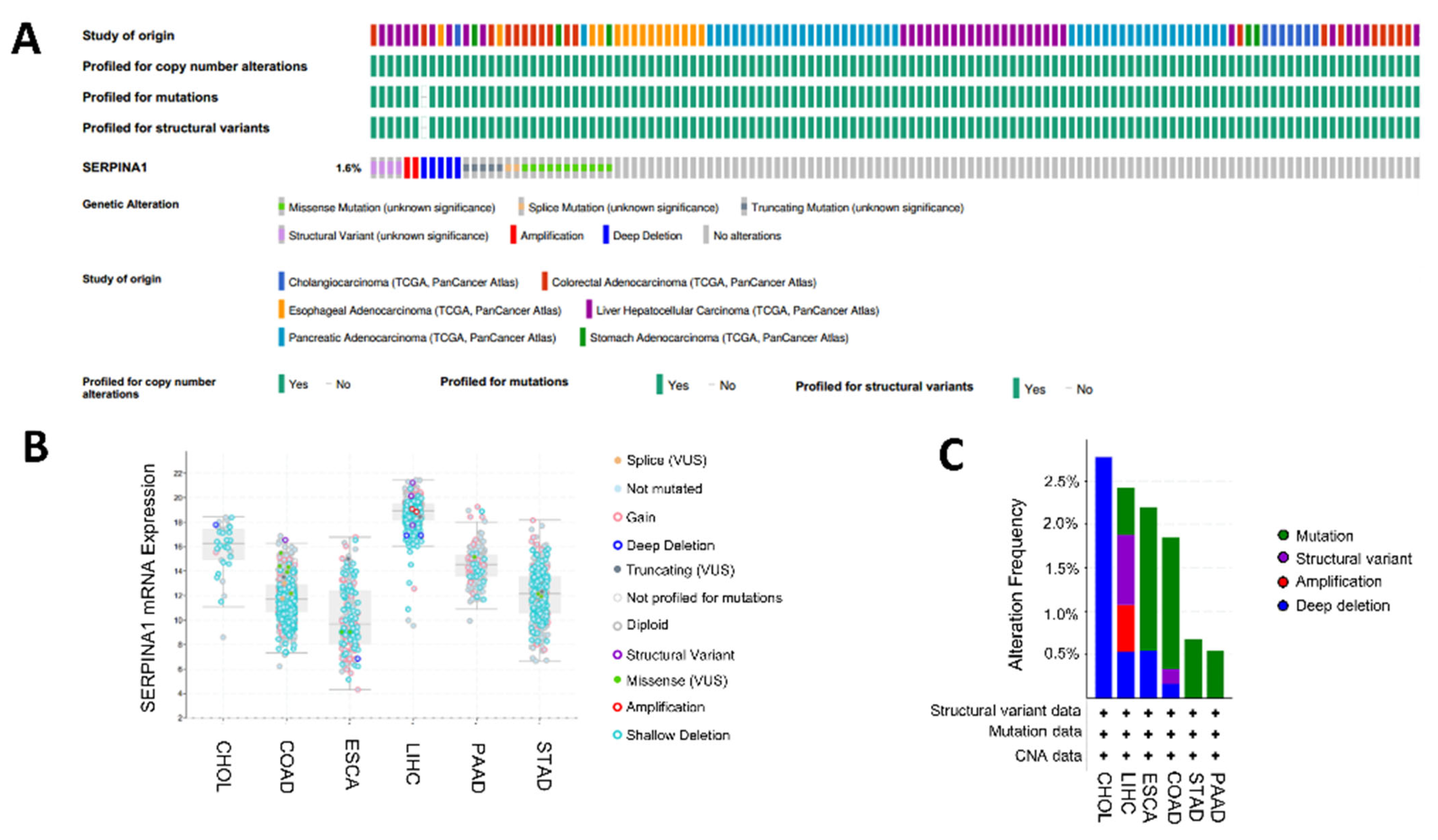
2.7. Alteration of SERPINA1 Gene in Different Subgroups of Digestive Carcinoma
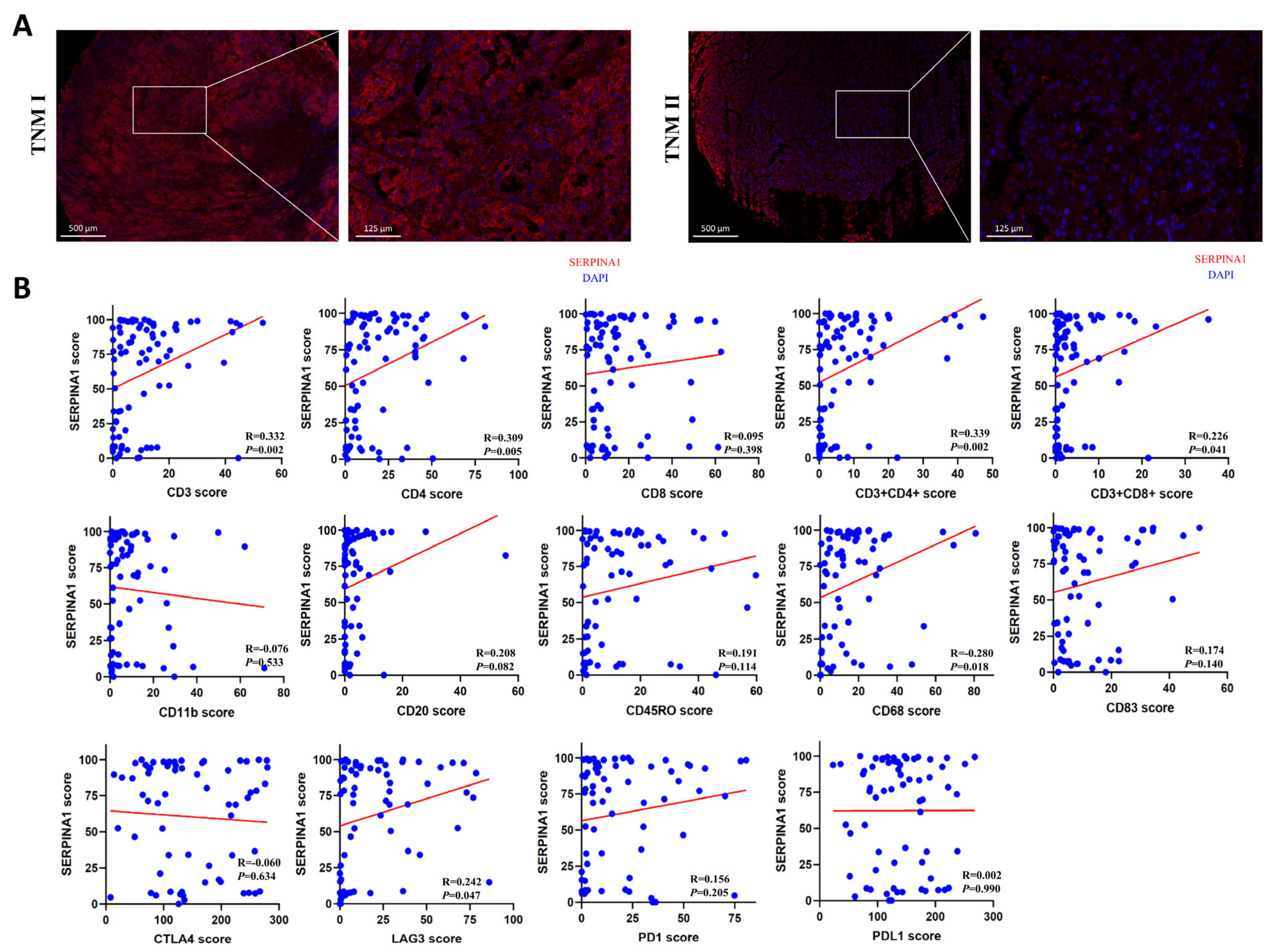
2.8. The Influence of SERPINA1 Protein on Clinicopathologic Features and Immune Infiltrates in LIHC
3. Discussion
4. Materials and Methods
4.1. Differential Expression Analysis
4.2. Survival Analysis
4.3. Analysis of SERPINA1 Expression and Immune and Molecular Subtypes
4.4. Analysis of SERPINA1 Expression and Biomarkers of Therapeutic Efficacy
4.5. Analysis of SERPINA1 Expression and Immune Cell Infiltration
4.6. Analysis of SERPINA1 Genomic Alterations
4.7. Analysis of SERPINA1 Expression and Clinicopathologic Features
4.8. Tissue Samples and Patient Data
4.9. Fluorescence-Based mIHC
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACC | Adrenocortical carcinoma |
| BLCA | Bladder urothelial carcinoma |
| BRCA | Breast invasive carcinoma |
| CESC | Cervical squamous cell carcinoma and endocervical adenocarcinoma |
| CHOL | Cholangiocarcinoma |
| CNV | Copy number variation |
| COAD | Colon adenocarcinoma |
| CTL | Cytotoxic lymphocyte |
| DC | Dendritic cell |
| DLBC | Lymphoid neoplasm diffuse large B-cell lymphoma |
| ESAD | Esophagus adenocarcinoma |
| ESCA | Esophageal carcinoma |
| ESTIMATE | Estimation of Stromal and Immune cells in Malignant Tumor tissues using Expression data |
| GBM | Glioblastoma multiforme |
| GBMLGG | Glioma |
| HNSC | Head and neck squamous cell carcinoma |
| ICP | Immune checkpoint |
| KICH | Kidney chromophobe |
| KIPAN | Pan-kidney cohort |
| KIRC | Kidney renal clear cell carcinoma |
| KIRP | Kidney renal papillary cell carcinoma |
| LGG | Brain lower grade glioma |
| LIHC | Liver hepatocellular carcinoma |
| LUAD | Lung adenocarcinoma |
| LUADLUSC | Lung adenocarcinoma and lung squamous cell carcinoma |
| LUSC | Lung squamous cell carcinoma |
| MESO | Mesothelioma |
| MSI | Microsatellite instability |
| NB | Neuroblastoma |
| NK | Natural killer cell |
| OS | Overall survival |
| OSCC | Oral squamous cell carcinoma |
| OV | Ovarian serous cystadenocarcinoma |
| PCPG | Pheochromocytoma and paraganglioma |
| PRAD | Prostate adenocarcinoma |
| SARC | Sarcoma |
| SERPINA1 | Serpin family A member 1 |
| SKCM | Skin cutaneous melanoma |
| STAD | Stomach adenocarcinoma |
| TCGA | The Cancer Genome Atlas |
| TGCT | Testicular germ cell tumors |
| THCA | Thyroid carcinoma |
| THYM | Thymoma |
| TMB | Tumor mutational burden |
| TME | Tumor microenvironment |
| UCEC | Uterine corpus endometrial carcinoma |
| UVM | Uveal melanoma |
| WT | High-risk Wilms tumor |
References
- Ruf, B.; Heinrich, B.; Greten, T.F. Immunobiology and immunotherapy of HCC:spotlight on innate and innate-like immune cells. Cell. Mol. Immunol. 2021, 18, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Li, J.; Shen, J.; Zhao, Y.; Xiao, Z. Identification of Prognostic Genes in the Tumor Microenvironment of Hepatocellular Carcinoma. Front. Immunol. 2021, 12, 653836. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yu, X.; Yang, F.; Zhang, Z.; Shen, J.; Sun, J.; Choksi, S.; Jitkaew, S.; Shu, Y. Reprogramming of Normal Fibroblasts into Cancer-Associated Fibroblasts by miRNAs-Mediated CCL2/VEGFA Signaling. PLoS Genet. 2016, 12, e1006244. [Google Scholar] [CrossRef] [PubMed]
- Kloth, J.N.; Gorter, A.; Fleuren, G.J.; Oosting, J.; Jordanova, E.S. Elevated expression of SerpinA1 and SerpinA3 in HLA-positive cervical carcinoma. J. Pathol. 2008, 215, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Seynes, C.D.; Ged, C.; Verneuil, H.D.; Chollet, N.; Balduyck, M.; Raherison, C. Identification of a novel alpha1-antitrypsin variant. Respir. Med. Case Rep. 2016, 20, 64–67. [Google Scholar] [CrossRef]
- Enewold, L.; Mechanic, L.E.; Bowman, E.D.; Platz, E.A.; Alberg, A.J. SERPINA1 and ELA2 polymorphisms are not associated with COPD or lung cancer. Anticancer. Res. 2012, 32, 3923–3928. [Google Scholar]
- Yang, J.; Xiong, X.; Wang, X.; Guo, B.; He, K.; Huang, C. Identification of peptide regions of SERPINA1 and ENOSF1 and their protein expression as potential serum biomarkers for gastric cancer. Tumour Biol. 2015, 36, 5109–5118. [Google Scholar] [CrossRef]
- Chan, H.J.; Li, H.; Zheng, L.; Yuan, Y.C.; Chen, S. SERPINA1 is a direct estrogen receptor target gene and a predictor of survival in breast cancer patients. Oncotarget 2015, 6, 25815–25827. [Google Scholar] [CrossRef]
- Morad, G.; Helmink, B.A.; Sharma, P.; Wargo, J.A.J.C. Hallmarks of response, resistance, and toxicity to immune checkpoint blockade. Cell 2021, 184, 5309–5337. [Google Scholar] [CrossRef]
- Hu, J.; Qiu, D.; Yu, A.; Hu, J.; Deng, H.; Li, H.; Yi, Z.; Chen, J.; Zu, X. YTHDF1 Is a Potential Pan-Cancer Biomarker for Prognosis and Immunotherapy. Front. Oncol. 2021, 11, 607224. [Google Scholar] [CrossRef]
- Liu, L.; Bai, X.; Wang, J.; Tang, X.R.; Dong, Z.Y. Combination of TMB and CNA Stratifies Prognostic and Predictive Responses to Immunotherapy Across Metastatic Cancer. Clin. Cancer Res. 2019, 25, 7413–7423. [Google Scholar] [CrossRef]
- Luchini, C.; Bibeau, F.; Ligtenberg, M.J.; Singh, N.; Nottegar, A.; Bosse, T.; Miller, R.; Riaz, N.; Douillard, J.Y.; Andre, F.; et al. ESMO recommendations on microsatellite instability testing for immunotherapy in cancer, and its relationship with PD-1/PD-L1 expression and tumour mutational burden: A systematic review-based approach. Ann. Oncol. 2019, 30, 1232–1243. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Denk, D.; Greten, F.R. Inflammation: The incubator of the tumor microenvironment. Trends Cancer 2022, 8, 901–914. [Google Scholar] [CrossRef]
- Kaneva, M.K.; Muley, M.M.; Krustev, E.; Reid, A.R.; Souza, P.R.; Dell’Accio, F.; McDougall, J.J.; Perretti, M. Alphantitrypsin reduces inflammation and exerts chondroprotection in arthritis. FASEB J. 2021, 35, e21472. [Google Scholar] [CrossRef]
- Bai, X.; Bai, A.; Tomasicchio, M.; Hagman, J.R.; Buckle, A.M.; Gupta, A.; Kadiyala, V.; Bevers, S.; Serban, K.A.; Kim, K.; et al. α1-Antitrypsin Binds to the Glucocorticoid Receptor with Anti-Inflammatory and Antimycobacterial Significance in Macrophages. J. Immunol. 2022, 209, 1746–1759. [Google Scholar] [CrossRef]
- Ehlers, M.R. Immune-modulating effects of alpha-1 antitrypsin. Biol. Chem. 2014, 395, 1187–1193. [Google Scholar] [CrossRef]
- Marcondes, A.M.; Karoopongse, E.; Lesnikova, M.; Margineantu, D.; Welte, T.; Dinarello, C.A.; Hockenbery, D.; Janciauskiene, S.; Deeg, H.J. α-1-Antitrypsin (AAT)-modified donor cells suppress GVHD but enhance the GVL effect: A role for mitochondrial bioenergetics. Blood 2014, 124, 2881–2891. [Google Scholar] [CrossRef]
- Tawara, I.; Sun, Y.; Lewis, E.C.; Toubai, T.; Evers, R.; Nieves, E.; Azam, T.; Dinarello, C.A.; Reddy, P. Alpha-1-antitrypsin monotherapy reduces graft-versus-host disease after experimental allogeneic bone marrow transplantation. Proc. Natl. Acad. Sci. USA 2012, 109, 564–569. [Google Scholar] [CrossRef]
- Elshikha, A.S.; Lu, Y.; Chen, M.-J.; Akbar, M.; Zeumer, L.; Ritter, A.; Elghamry, H.; Mahdi, M.A.; Morel, L.; Song, S. Alpha 1 Antitrypsin Inhibits Dendritic Cell Activation and Attenuates Nephritis in a Mouse Model of Lupus. PLoS ONE 2016, 11, e0156583. [Google Scholar] [CrossRef]
- Trachte, A.L.; Suthers, S.E.; Lerner, M.R.; Hanas, J.S.; Jupe, E.R.; Sienko, A.E.; Adesina, A.M.; Lightfoot, S.A.; Brackett, D.J.; Postier, R.G. Increased expression of alpha-1-antitrypsin, glutathione S-transferase p and vascular endothelial growth factor in human pancreatic adenocarcinoma. Am. J. Surg. 2002, 184, 642–647. [Google Scholar] [CrossRef] [PubMed]
- López-Árias, E.; Aguilar-Lemarroy, A.; Jave-Suárez, L.F.; Morgan-Villela, G.; Mariscal-Ramírez, I.; Martínez-Velázquez, M.; Álvarez, Á.H.; Gutiérrez-Ortega, A.; Gutiérrez, R.H. Alpha 1-antitrypsin: A novel tumor-associated antigen identified in patients with early-stage breast cancer. Electrophoresis 2012, 33, 2130–2137. [Google Scholar] [CrossRef] [PubMed]
- Ercetin, E.; Richtmann, S.; Delgado, B.M.; Gomez-Mariano, G.; Schneider, M.A. Clinical Significance of SERPINA1 Gene and Its Encoded Alpha1-antitrypsin Protein in NSCLC. Cancers 2019, 11, 1306. [Google Scholar] [CrossRef] [PubMed]
- Pirisi, M.; Fabris, C.; Soardo, G.; Toniutto, P.; Bartoli, E. Prognostic value of serum alpha-1-antitrypsin in hepatocellular carcinoma. Eur. J. Cancer 1996, 32A, 221–225. [Google Scholar] [CrossRef]
- Wu, C.-C.; Lu, Y.-T.; Yeh, T.-S.; Chan, Y.-H.; Dash, S.; Yu, J.-S. Identification of Fucosylated SERPINA1 as a Novel Plasma Marker for Pancreatic Cancer Using Lectin Affinity Capture Coupled with iTRAQ-Based Quantitative Glycoproteomics. Int. J. Mol. Sci. 2021, 22, 6079. [Google Scholar] [CrossRef]
- Kwon, C.H.; Park, H.J.; Choi, J.H.; Lee, J.R.; Kim, H.K.; Jo, H.-J.; Kim, H.S.; Oh, N.; Song, G.A.; Park, D.Y. Snail and serpinA1 promote tumor progression and predict prognosis in colorectal cancer. Oncotarget 2015, 6, 20312–20326. [Google Scholar] [CrossRef]
- Topalian, S.L.; Taube, J.M.; Anders, R.A.; Pardoll, D.M. Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy. Nat. Rev. Cancer 2016, 16, 275–287. [Google Scholar] [CrossRef]
- McGrail, D.J.; Pilié, P.G.; Rashid, N.U.; Voorwerk, L.; Slagter, M.; Kok, M.; Jonasch, E.; Khasraw, M.; Heimberger, A.B.; Lim, B.; et al. High tumor mutation burden fails to predict immune checkpoint blockade response across all cancer types. Ann. Oncol. 2021, 32, 661–672. [Google Scholar] [CrossRef]
- Zhao, P.; Li, L.; Jiang, X.; Li, Q. Mismatch repair deficiency/microsatellite instability-high as a predictor for anti-PD-1/PD-L1 immunotherapy efficacy. J. Hematol. Oncol. 2019, 12, 54. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef]
- Okadome, K.; Baba, Y.; Yagi, T.; Kiyozumi, Y.; Ishimoto, T.; Iwatsuki, M.; Miyamoto, Y.; Yoshida, N.; Watanabe, M.; Baba, H. Prognostic Nutritional Index, Tumor-infiltrating Lymphocytes, and Prognosis in Patients with Esophageal Cancer. Ann. Surg. 2020, 271, 693–700. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z. The history and advances in cancer immunotherapy:understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef]
- Ribas, A.; Dummer, R.; Puzanov, I.; VanderWalde, A.; Andtbacka, R.H.I.; Michielin, O.; Olszanski, A.J.; Malvehy, J.; Cebon, J.; Fernandez, E. Oncolytic Virotherapy Promotes Intratumoral T Cell Infiltration and Improves Anti-PD-1 Immunotherapy. Cell 2017, 170, 1109–1119.e10. [Google Scholar] [CrossRef]
- Chen, D.; Menon, H.; Verma, V.; Guo, C.; Ramapriyan, R.; Barsoumian, H.; Younes, A.; Hu, Y.; Wasley, M.; Cortez, M.A.; et al. Response and outcomes after anti-CTLA4 versus anti-PD1 combined with stereotactic body radiation therapy for metastatic non-small cell lung cancer: Retrospective analysis of two single-institution prospective trials. J. Immunother. Cancer 2020, 8, e000492. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; David, C.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Chu, L.; Yi, Q.; Yan, Y.; Peng, J.; Li, Z.; Jiang, F.; He, Q.; Ouyang, L.; Wu, S.; Fu, C.; et al. A Prognostic Signature Consisting of Pyroptosis-Related Genes and SCAF11 for Predicting Immune Response in Breast Cancer. Front. Med. 2022, 9, 882763. [Google Scholar] [CrossRef]
- Ru, B.; Ngar, W.C.; Tong, Y.; Zhong, J.Y.; Zhong, S.; Wu, W.C.; Chu, K.C.; Yiu, W.C.; Ying, L.C.; Chen, I. TISIDB: An integrated repository portal for tumor–immune system interactions. Bioinformatics 2019, 35, 4200–4202. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Gene Markers | ESCA | LIHC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| None | Purity | None | Purity | ||||||
| R | p | R | p | R | p | R | p | ||
| CD8+ T Cell | CD8A | 0.229 | 0.002 * | 0.194 | 0.009 | 0.106 | 0.041 * | 0.011 | 0.834 |
| CD8B | 0.321 | <0.001 * | 0.293 | <0.001 * | 0.069 | 0.184 | −0.020 | 0.718 | |
| T Cell (general) | CD3D | 0.358 | <0.001 * | 0.328 | <0.001 * | 0.144 | 0.006 * | 0.053 | 0.328 |
| CD3E | 0.321 | <0.001 * | 0.282 | <0.001 * | 0.228 | <0.001 * | 0.133 | 0.014 * | |
| CD2 | 0.384 | <0.001 * | 0.354 | <0.001 * | 0.191 | <0.001 * | 0.095 | 0.078 | |
| B Cell | CD19 | 0.362 | <0.001 * | 0.325 | <0.001 * | 0.082 | 0.114 | 0.006 | 0.905 |
| CD79A | 0.277 | <0.001 * | 0.238 | <0.001 * | 0.218 | <0.001 * | 0.116 | 0.031 * | |
| Monocyte | CD86 | 0.066 | 0.375 | 0.019 | 0.801 | 0.114 | 0.029 * | −0.003 | 0.961 |
| CSF1R | 0.282 | <0.001 * | 0.257 | <0.001 * | 0.098 | 0.059 | −0.002 | 0.680 | |
| TAM | CCL2 | 0.132 | 0.0744 | 0.088 | 0.242 | 0.083 | 0.112 | −0.045 | 0.407 |
| CD68 | 0.258 | <0.001 * | 0.258 | <0.001 * | 0.161 | 0.002 * | 0.069 | 0.199 | |
| IL10 | 0.125 | 0.090 | 0.087 | 0.247 | 0.119 | 0.022 * | 0.016 | 0.760 | |
| M1 | NOS2 | 0.451 | <0.001 * | 0.47 | <0.001 * | 0.035 | 0.499 | 0.022 | 0.680 |
| IRF5 | −0.286 | <0.001 * | −0.317 | <0.001 * | −0.109 | 0.036 * | −0.133 | 0.013 * | |
| PTGS2 | 0.06 | 0.417 | 0.043 | 0.571 | 0.195 | <0.001 * | 0.087 | 0.106 | |
| M2 | CD163 | 0.339 | <0.001 * | 0.318 | <0.001 * | 0.161 | 0.002 * | 0.070 | 0.197 |
| VSIG4 | 0.238 | 0.001 * | 0.206 | 0.005 * | 0.101 | 0.052 | −0.003 | 0.949 | |
| MS4A4A | 0.287 | <0.001 * | 0.253 | <0.001 * | 0.172 | <0.001 * | 0.065 | 0.229 | |
| Neutrophils | CEACAM8 | 0.293 | <0.001 * | 0.274 | <0.001 * | 0.074 | 0.155 | 0.058 | 0.284 |
| ITGAM | 0.251 | <0.001 * | 0.234 | 0.001 * | −0.063 | 0.226 | −0.162 | 0.003 * | |
| CCR7 | 0.428 | <0.001 * | 0.401 | <0.001 * | 0.278 | <0.001 * | 0.1869 | <0.001 * | |
| NK Cell | KIR2DL1 | 0.123 | 0.094 | 0.094 | 0.210 | −0.046 | 0.367 | −0.062 | 0.250 |
| KIR2DL3 | 0.035 | 0.636 | 0.033 | 0.665 | −0.061 | 0.240 | −0.118 | 0.028 * | |
| KIR2DL4 | 0.078 | 0.29 | 0.05 | 0.501 | −0.118 | 0.023 * | −0.163 | 0.002 * | |
| KIR3DL1 | 0.171 | 0.020 * | 0.153 | 0.040 * | −0.063 | 0.023 * | −0.109 | 0.042 * | |
| KIR3DL2 | −0.049 | 0.508 | −0.092 | 0.221 | −0.029 | 0.573 | −0.084 | 0.121 | |
| KIR2DL3 | 0 | 0.996 | 0 | 0.999 | −0.061 | 0.240 | −0.118 | 0.028 * | |
| KIR2DS4 | 0.042 | 0.571 | 0.041 | 0.583 | −0.078 | 0.135 | −0.085 | 0.117 | |
| Dendritic Cell | HLA-DPB1 | 0.4 | <0.001 * | 0.38 | <0.001 * | 0.15 | 0.004 * | 0.055 | 0.307 |
| HLA-DQB1 | 0.362 | <0.001 * | 0.329 | <0.001 * | 0.11 | 0.034 * | 0.018 | 0.743 | |
| HLA-DRA | 0.456 | <0.001 * | 0.441 | <0.001 * | 0.167 | 0.001 * | 0.078 | 0.151 | |
| HLA-DPA1 | 0.431 | <0.001 * | 0.414 | <0.001 * | 0.178 | <0.001 * | 0.089 | 0.094 | |
| CD1C | 0.063 | 0.393 | 0.002 | 0.977 | 0.217 | <0.001 * | 0.131 | 0.015 * | |
| NRP1 | 0.178 | 0.016 * | 0.142 | 0.058 * | 0.060 | 0.250 | 0.021 | 0.697 | |
| ITGAX | 0.355 | <0.001 * | 0.24 | <0.001 * | 0.126 | 0.015 * | 0.027 | 0.624 | |
| Th1 | TBX21 | 0.308 | <0.001 * | 0.272 | <0.001 * | 0.131 | 0.012 * | 0.034 | 0.526 |
| STAT4 | 0.275 | <0.001 * | 0.237 | 0.001 * | 0.12 | 0.021 * | 0.043 | 0.422 | |
| STAT1 | 0.111 | 0.134 | 0.085 | 0.250 | 0.175 | <0.001 * | 0.136 | 0.011 * | |
| IFNG | 0.205 | 0.005 * | 0.172 | 0.021 * | −0.022 | 0.669 | −0.083 | 0.122 | |
| TNF | 0.066 | 0.373 | 0.046 | 0.537 | 0.155 | 0.003 * | 0.075 | 0.165 | |
| IL12A | 0.201 | 0.006 * | 0.19 | 0.011 * | 0.008 | 0.884 | −0.040 | 0.454 | |
| IL12B | 0.262 | <0.001 * | 0.227 | 0.002 * | 0.131 | 0.012 * | 0.049 | 0.368 | |
| Th2 | GATA3 | 0.07 | 0.340 | 0.034 | 0.649 | 0.107 | 0.040 * | −0.002 | 0.965 |
| STAT6 | 0.278 | <0.001 * | 0.302 | <0.001 * | −0.040 | 0.444 | −0.050 | 0.352 | |
| STAT5A | 0.444 | <0.001 * | 0.432 | <0.001 * | −0.116 | 0.026 * | −0.189 | <0.001 * | |
| IL13 | 0.173 | 0.019 * | 0.146 | 0.050 | −0.099 | 0.057 | −0.12 | 0.026 * | |
| Tfh | BCL6 | −0.504 | <0.001 * | −0.512 | <0.001 * | 0.019 | 0.711 | 0.011 | 0.833 |
| IL21 | 0.09 | 0.225 | 0.059 | 0.432 | 0.045 | 0.392 | 0.012 | 0.821 | |
| Th17 | STAT3 | 0.059 | 0.427 | 0.047 | 0.531 | 0.161 | 0.002 * | 0.129 | 0.016 * |
| IL17A | 0.378 | <0.001 * | 0.379 | <0.001 * | 0.079 | 0.131 | 0.067 | 0.218 | |
| Treg | FOXP3 | 0.255 | <0.001 * | 0.222 | 0.003 * | 0.025 | 0.633 | −0.035 | 0.518 |
| CCR8 | 0.314 | <0.001 * | 0.282 | <0.001 * | 0.179 | <0.001 * | 0.107 | 0.048 * | |
| STAT5B | 0.189 | 0.010 * | 0.193 | 0.009 * | −0.022 | 0.677 | 0.013 | 0.814 | |
| TGFB1 | −0.508 | <0.001 * | −0.574 | <0.001 * | 0.205 | <0.001 * | 0.113 | 0.036 * | |
| T cell exhaustion | PDCD1 | 0.301 | <0.001 * | 0.272 | <0.001 * | 0.119 | 0.021 * | 0.028 | 0.608 |
| CTLA4 | 0.292 | <0.001 * | 0.261 | <0.001 * | 0.114 | 0.028 * | 0.029 | 0.589 | |
| LAG3 | 0.143 | 0.052 | 0.106 | 0.158 | −0.032 | 0.537 | −0.091 | 0.091 | |
| HAVCR2 | 0.256 | <0.001 * | 0.227 | 0.002 * | 0.080 | 0.126 | −0.047 | 0.380 | |
| GZMB | 0.115 | 0.120 | 0.073 | 0.333 | −0.013 | 0.801 | −0.093 | 0.085 | |
| Characteristics | n | SERPINA1 Expression (%) | χ2 | p | |
|---|---|---|---|---|---|
| Low or No | High | ||||
| Total | 86 | 37 (43.02) | 49 (56.98) | ||
| Gender | 0.054 | 0.816 | |||
| Male | 64 | 28 (43.75) | 36 (56.25) | ||
| Female | 22 | 9 (40.91) | 13 (59.09) | ||
| Age | 0.042 | 0.837 | |||
| <60 | 76 | 33 (43.42) | 43 (56.58) | ||
| ≥60 | 10 | 4 (40.00) | 6 (60.00) | ||
| Hepatitis B virus infection | 2.447 | 0.118 | |||
| 0 | 10 | 2 (20.00) | 8 (80.00) | ||
| 1 | 76 | 35 (46.05) | 41 (53.95) | ||
| Differentiation | 3.046 | 0.218 | |||
| Well | 8 | 3 (37.50) | 5 (62.50) | ||
| Moderate | 62 | 24 (38.71) | 38 (61.29) | ||
| Poor | 16 | 10 (62.50) | 6 (37.50) | ||
| Vascular invasion | 6.394 | 0.011 * | |||
| 0 | 46 | 14 (30.43) | 32 (69.57) | ||
| 1 | 40 | 23 (57.50) | 17 (42.50) | ||
| T | 4.879 | 0.027 * | |||
| T1 | 42 | 13 (30.95) | 29 (69.05) | ||
| T2 + T3 | 44 | 24 (54.55) | 20 (45.45) | ||
| N | 2.712 | 0.100 | |||
| N0 | 84 | 35 (41.67) | 49 (58.33) | ||
| N1 | 2 | 2 (100.00) | 0 (0.00) | ||
| TNM stage | 4.879 | 0.027 * | |||
| Ⅰ | 42 | 13 (30.95) | 29 (69.05) | ||
| Ⅱ + Ⅲ | 44 | 24 (54.55) | 20 (45.45) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuai, X.; Lv, J.; Zhang, J.; Xu, M.; Ji, J. Serpin Family A Member 1 Is Prognostic and Involved in Immunological Regulation in Human Cancers. Int. J. Mol. Sci. 2023, 24, 11566. https://doi.org/10.3390/ijms241411566
Kuai X, Lv J, Zhang J, Xu M, Ji J. Serpin Family A Member 1 Is Prognostic and Involved in Immunological Regulation in Human Cancers. International Journal of Molecular Sciences. 2023; 24(14):11566. https://doi.org/10.3390/ijms241411566
Chicago/Turabian StyleKuai, Xingwang, Jiaying Lv, Junyu Zhang, Manyu Xu, and Juling Ji. 2023. "Serpin Family A Member 1 Is Prognostic and Involved in Immunological Regulation in Human Cancers" International Journal of Molecular Sciences 24, no. 14: 11566. https://doi.org/10.3390/ijms241411566
APA StyleKuai, X., Lv, J., Zhang, J., Xu, M., & Ji, J. (2023). Serpin Family A Member 1 Is Prognostic and Involved in Immunological Regulation in Human Cancers. International Journal of Molecular Sciences, 24(14), 11566. https://doi.org/10.3390/ijms241411566