
N,N-Dimethylglycine Sodium Salt Exerts Marked Anti-Inflammatory Effects in Various Dermatitis Models and Activates Human Epidermal Keratinocytes by Increasing Proliferation, Migration, and Growth Factor Release


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
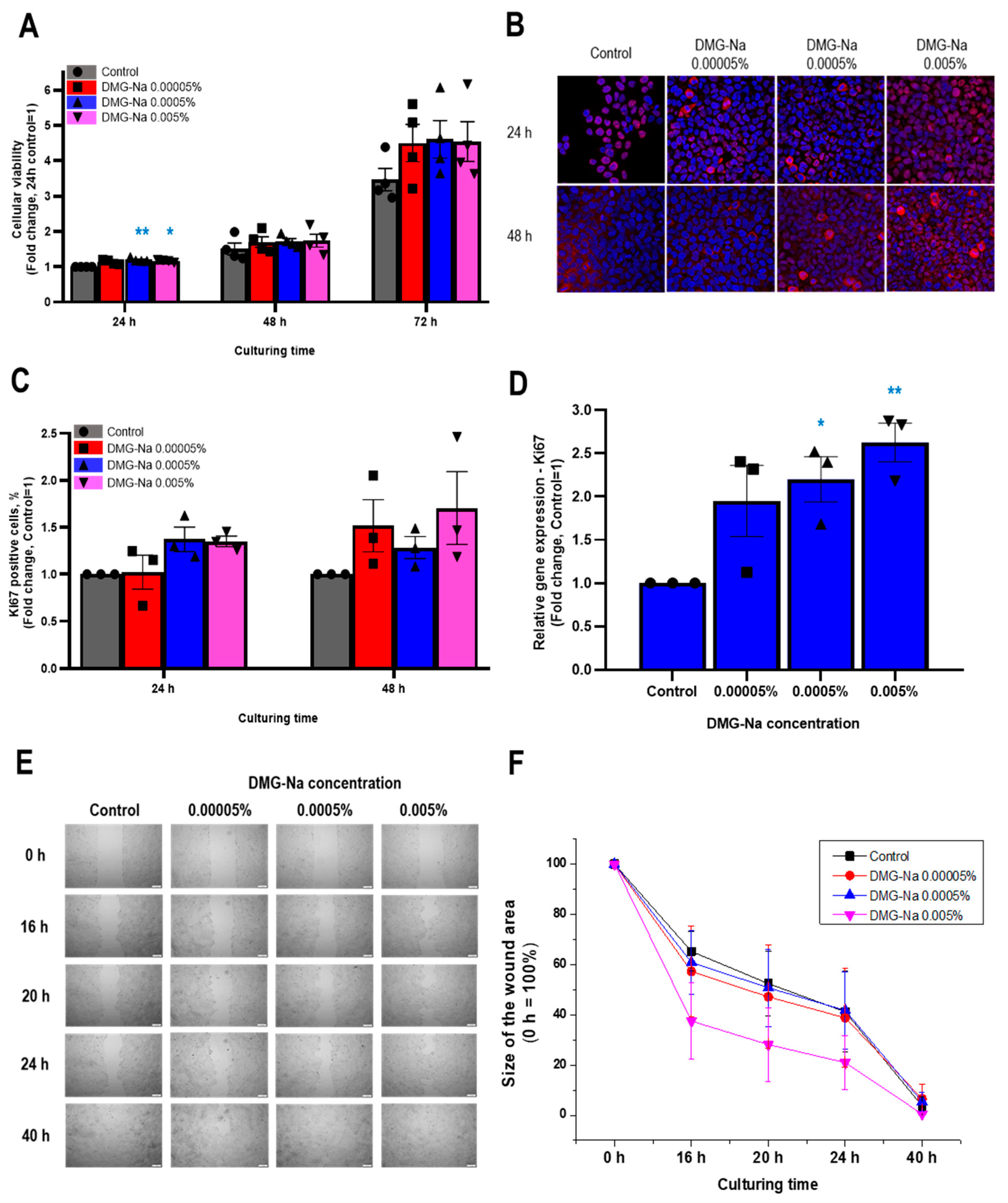
2.1. DMG-Na Promotes Viability, Cell Proliferation, Migration, and the Release of Specific Growth Factors in Human Epidermal Keratinocytes
2.1.1. DMG-Na Maintains Cellular Viability of Human Epidermal HaCaT Keratinocytes
2.1.2. DMG-Na Increases the Number of Ki67-Positive Cells and Upregulates the mRNA-Level Expression of Ki67
2.1.3. DMG-Na Promotes Migration of Human Epidermal HaCaT Keratinocytes
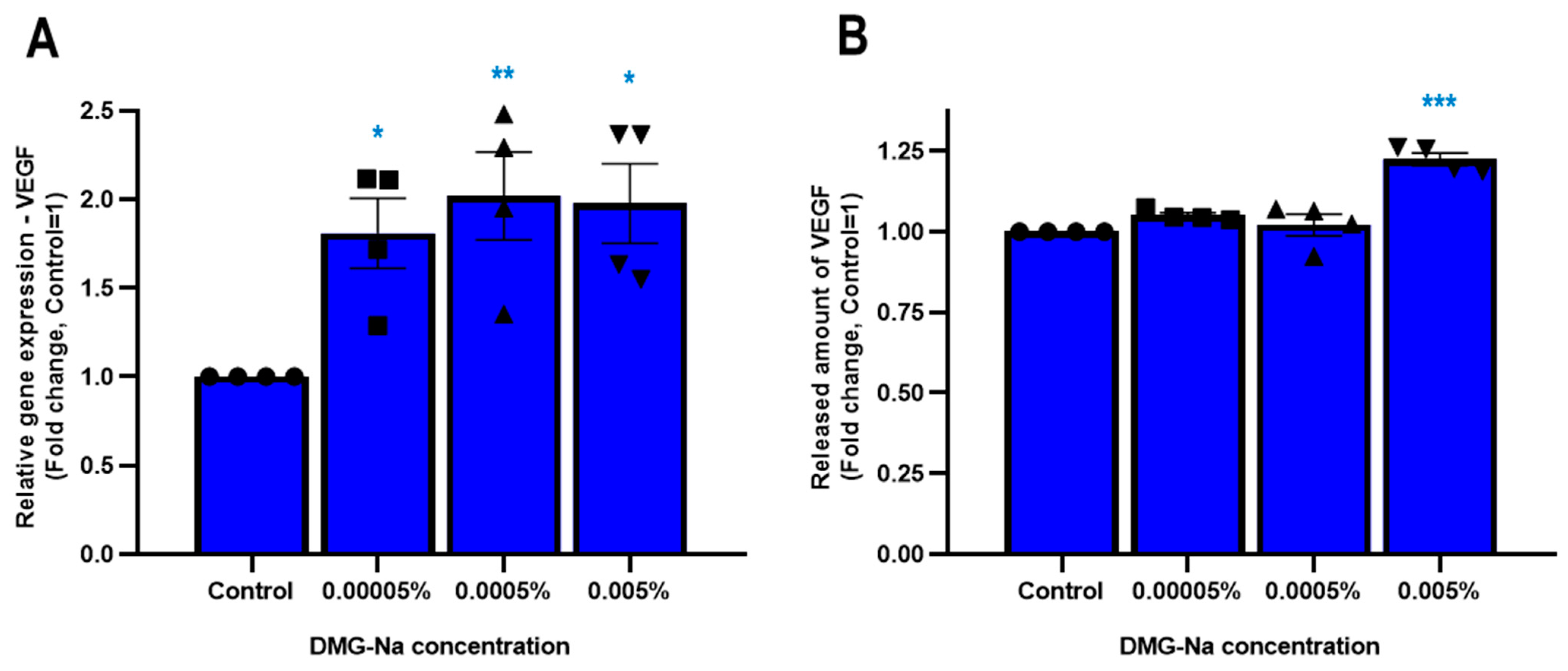
2.1.4. DMG-Na Upregulates the Synthesis and Release of Specific Growth Factors in Human Epidermal HaCaT Keratinocytes
2.2. DMG-Na Exerts Anti-Inflammatory Effects in Three In Vitro Keratinocyte Models of Various Inflammatory Skin Conditions
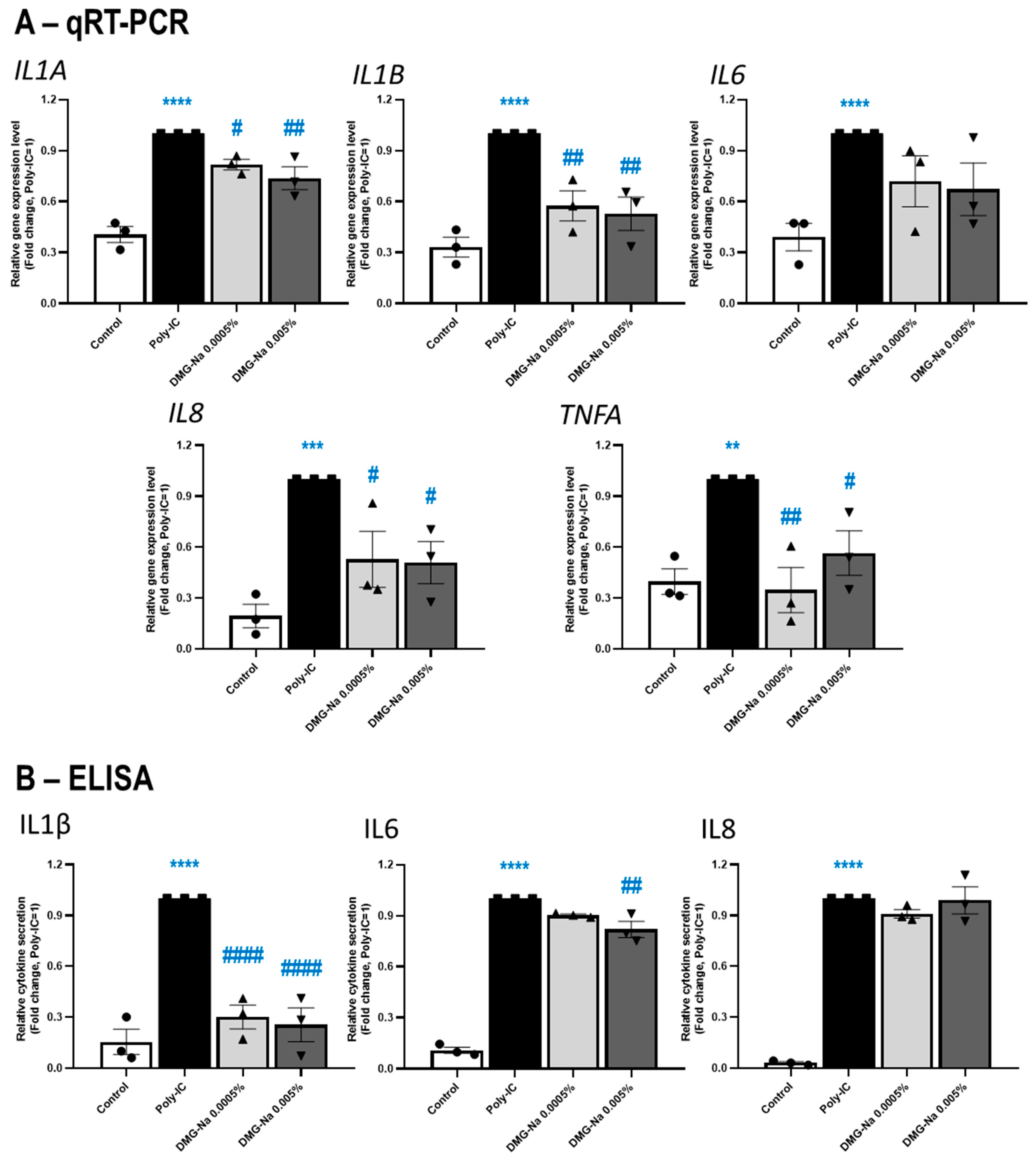
2.2.1. DMG-Na Exerts Anti-Inflammatory Effects in an In Vitro Keratinocyte Model of Microbial and Allergic Contact Dermatitis
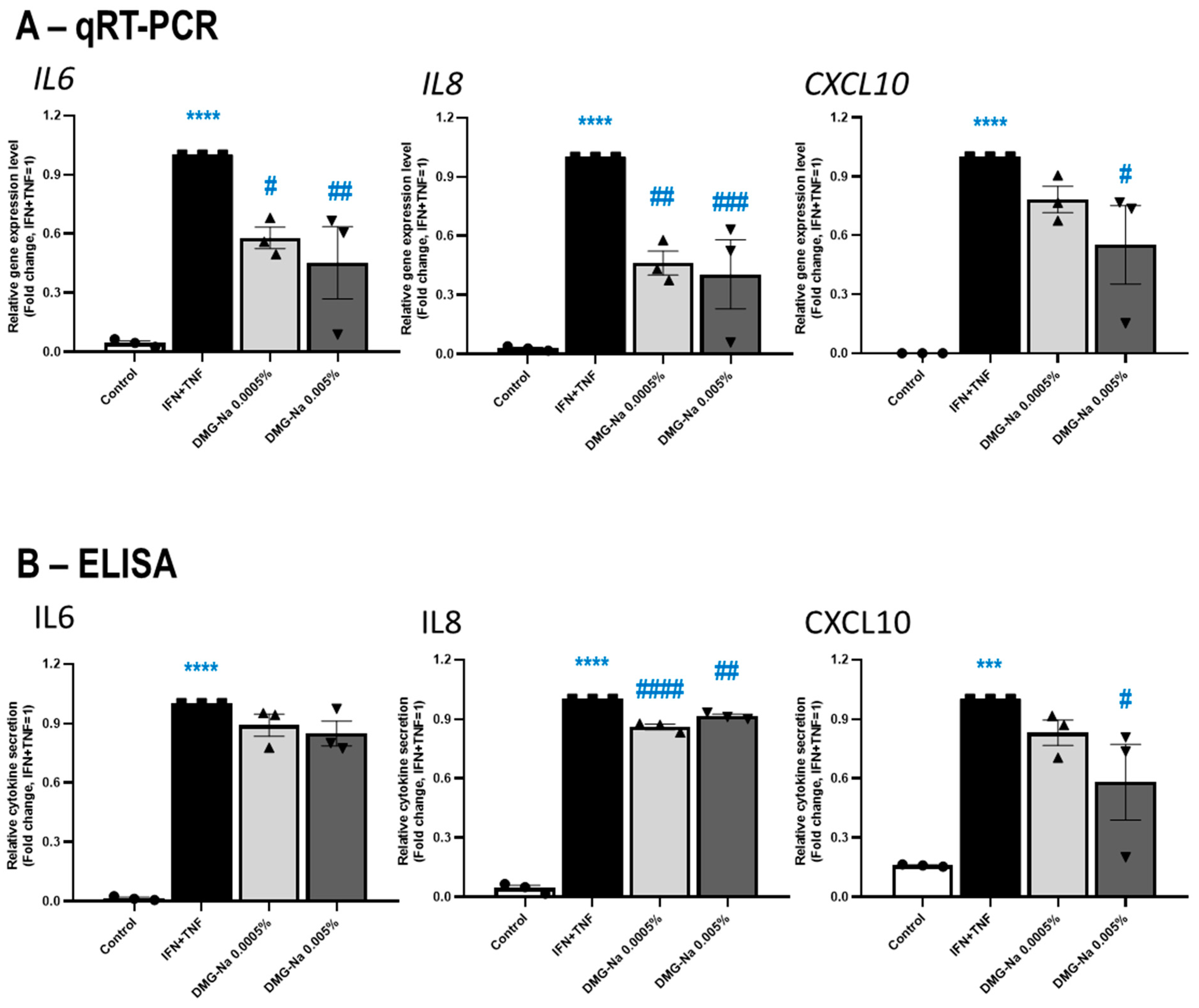
2.2.2. DMG-Na Exerts Anti-Inflammatory Effects in an In Vitro Keratinocyte Model of Various Inflammatory Skin Diseases Such as Psoriasis
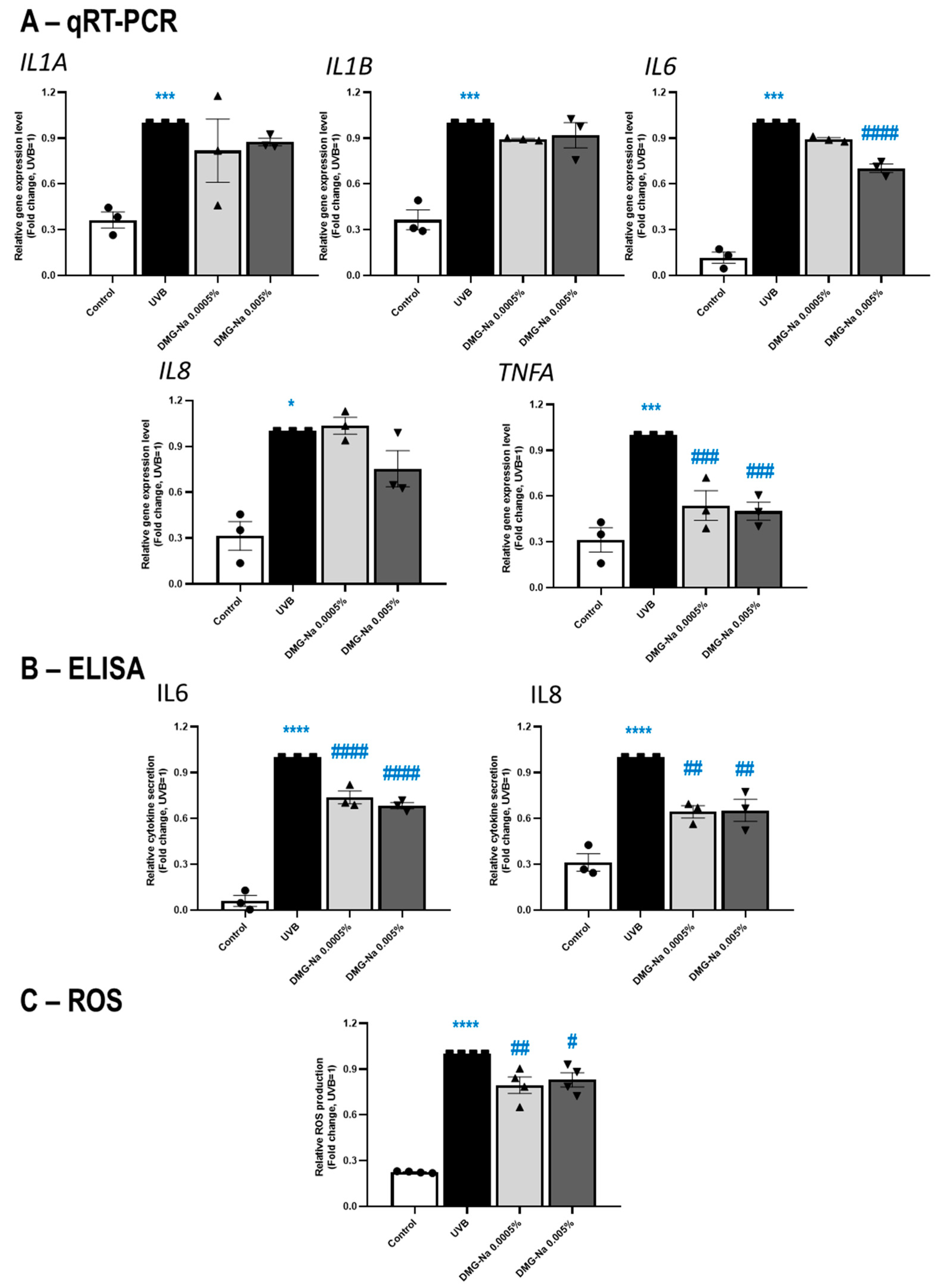
2.2.3. DMG-Na Exerts Anti-Inflammatory and Antioxidant Effects in an In Vitro Keratinocyte Model of UVB Irradiation-Induced Solar Dermatitis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culturing
4.3. MTT Assay
4.4. Immunocytochemistry
4.5. Scratch Wound-Closure Migration Assay
4.6. Inflammatory Model Systems
4.6.1. In Vitro Keratinocyte Model of Microbial and Allergic Contact Dermatitis
4.6.2. In Vitro Keratinocyte Model of Inflammatory Skin Diseases Such as Psoriasis
4.6.3. UVB Irradiation-Induced Inflammation and Oxidative Stress Model
4.7. RNA Isolation and Reverse Transcription
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. Ca-Imaging—Microfluorimetric Measurements of Intracellular Ca Concentration ([Ca2+]i)
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Freed, W.J. N,N-Dimethylglycine, Betaine, and Seizures. Arch. Neurol. 1984, 41, 1129–1130. [Google Scholar] [CrossRef] [PubMed]
- Cupp, M.J.; Tracy, T.S. Dimethylglycine (N,N-Dimethylglycine). In Dietary Supplements: Toxicology and Clinical Pharmacology; Cupp, M.J., Tracy, T.S., Eds.; Humana Press: Totowa, NJ, USA, 2003; pp. 149–160. ISBN 9781592593033. [Google Scholar]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Assessment of the Feed Additive Consisting of Dimethylglycine Sodium Salt (Taminizer D) for Chickens for Fattening for the Renewal of Its Authorisation (Taminco N.V.). EFSA J. 2021, 19, e06621. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Safety and Efficacy of Taminizer D (dimethylglycine Sodium Salt) as a Feed Additive for Chickens for Fattening. EFSA J. 2018, 16, e05268. [Google Scholar] [CrossRef]
- Kalmar, I.D.; Cools, A.; Buyse, J.; Roose, P.; Janssens, G.P.J. Dietary N,N-Dimethylglycine Supplementation Improves Nutrient Digestibility and Attenuates Pulmonary Hypertension Syndrome in Broilers. J. Anim. Physiol. Anim. Nutr. 2010, 94, e339–e347. [Google Scholar] [CrossRef]
- Kalmar, I.D.; Cools, A.; Verstegen, M.W.A.; Huyghebaert, G.; Buyse, J.; Roose, P.; Janssens, G.P.J. Dietary Supplementation with Dimethylglycine Affects Broiler Performance and Plasma Metabolites Depending on Dose and Dietary Fatty Acid Profile. J. Anim. Physiol. Anim. Nutr. 2011, 95, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Kalmar, I.D.; Verstegen, M.W.A.; Maenner, K.; Zentek, J.; Meulemans, G.; Janssens, G.P.J. Tolerance and Safety Evaluation of N,N-Dimethylglycine, a Naturally Occurring Organic Compound, as a Feed Additive in Broiler Diets. Br. J. Nutr. 2011, 107, 1635–1644. [Google Scholar] [CrossRef]
- Kalmar, I.D.; Verstegen, M.W.A.; Vanrompay, D.; Maenner, K.; Zentek, J.; Iben, C.; Leitgeb, R.; Schiavone, A.; Prola, L.; Janssens, G.P.J. Efficacy of Dimethylglycine as a Feed Additive to Improve Broiler Production. Livest. Sci. 2014, 164, 81–86. [Google Scholar] [CrossRef]
- Prola, L.; Nery, J.; Lauwaerts, A.; Bianchi, C.; Sterpone, L.; De Marco, M.; Pozzo, L.; Schiavone, A. Effects of N,N-Dimethylglycine Sodium Salt on Apparent Digestibility, Vitamin E Absorption, and Serum Proteins in Broiler Chickens Fed a High- or Low-Fat Diet. Poult. Sci. 2013, 92, 1221–1226. [Google Scholar] [CrossRef]
- Feng, C.; Bai, K.; Wang, A.; Ge, X.; Zhao, Y.; Zhang, L.; Wang, T. Effects of Dimethylglycine Sodium Salt Supplementation on Growth Performance, Hepatic Antioxidant Capacity, and Mitochondria-Related Gene Expression in Weanling Piglets Born with Low Birth weight1. J. Anim. Sci. 2018, 96, 3791–3803. [Google Scholar] [CrossRef]
- Greene, H.; Wickler, S.J.; Bray, R.E.; Burrill, M.J.; London, C. The Effect of N,N-Dimethylglycine on Athletic Performance at Altitude in Horses and Mules. Pferdeheilkunde 1996, 12, 499–501. [Google Scholar] [CrossRef]
- Pipes, T.V. The Effects of Pangamic Acid on Performance in Trained Athletes. Med. Sci. Sports. Exerc. 1980, 12, 98. [Google Scholar]
- Kern, J.K.; Miller, V.S.; Cauller, P.L.; Kendall, P.R.; Mehta, P.J.; Dodd, M. Effectiveness of N,N-Dimethylglycine in Autism and Pervasive Developmental Disorder. J. Child Neurol. 2001, 16, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Jiang, L.; Zhang, L.; Zhao, Y.; Lu, Y.; Zhu, J.; Cai, J.; Zhang, L.; Wang, T. In Vitro Free Radical Scavenging Capacity of Dimethylglycine Sodium Salt and Its Protective Ability against Oleic Acid Hydroperoxide-Induced Oxidative Damage in IPEC-J2 Cells. Int. J. Mol. Med. 2018, 42, 3447–3458. [Google Scholar] [CrossRef]
- Bai, K.; Jiang, L.; Zhu, S.; Feng, C.; Zhao, Y.; Zhang, L.; Wang, T. Dimethylglycine Sodium Salt Protects against Oxidative Damage and Mitochondrial Dysfunction in the Small Intestines of Mice. Int. J. Mol. Med. 2019, 43, 2199–2211. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Jiang, L.; Li, Q.; Zhang, J.; Zhang, L.; Wang, T. Dietary Dimethylglycine Sodium Salt Supplementation Alleviates Redox Status Imbalance and Intestinal Dysfunction in Weaned Piglets with Intrauterine Growth Restriction. Anim. Nutr. 2022, 10, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Bai, K.; Xu, W.; Zhang, J.; Kou, T.; Niu, Y.; Wan, X.; Zhang, L.; Wang, C.; Wang, T. Assessment of Free Radical Scavenging Activity of Dimethylglycine Sodium Salt and Its Role in Providing Protection against Lipopolysaccharide-Induced Oxidative Stress in Mice. PLoS ONE 2016, 11, e0155393. [Google Scholar] [CrossRef]
- Bai, K.; Jiang, L.; Wang, T. Dimethylglycine Sodium Salt Alleviates Intrauterine Growth Restriction-Induced Low Growth Performance, Redox Status Imbalance, and Hepatic Mitochondrial Dysfunction in Suckling Piglets. Front. Vet. Sci. 2022, 9, 905488. [Google Scholar] [CrossRef]
- Bai, K.; Jiang, L.; Li, Q.; Zhang, J.; Zhang, L.; Wang, T. Dietary Dimethylglycine Sodium Salt Supplementation Improves Growth Performance, Redox Status, and Skeletal Muscle Function of Intrauterine Growth-Restricted Weaned Piglets. J. Anim. Sci. 2021, 99, skab186. [Google Scholar] [CrossRef]
- Bai, K.; Jiang, L.; Li, Q.; Zhang, J.; Zhang, L.; Wang, T. Supplementation of Dimethylglycine Sodium Salt in Sow Milk Reverses Skeletal Muscle Redox Status Imbalance and Mitochondrial Dysfunction of Intrauterine Growth Restriction Newborns. Antioxidants 2022, 11, 1550. [Google Scholar] [CrossRef]
- Bai, K.; Jiang, L.; Wei, C.; Li, Q.; Zhang, L.; Zhang, J.; Wang, T. Dimethylglycine Sodium Salt Activates Nrf2/SIRT1/PGC1α Leading to the Recovery of Muscle Stem Cell Dysfunction in Newborns with Intrauterine Growth Restriction. Free Radic. Biol. Med. 2022, 184, 89–98. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, C.; Liu, X.; Jiang, C.-X.; Cao, Q.; Yu, B.; Ni, Y.; Mao, S. The Metabolomic Profiling Identifies N, N-Dimethylglycine as a Facilitator of Dorsal Root Ganglia Neuron Axon Regeneration after Injury. FASEB J. 2022, 36, e22305. [Google Scholar] [CrossRef]
- Takahashi, T.; Sasaki, K.; Somfai, T.; Nagai, T.; Manabe, N.; Edashige, K. N,N-Dimethylglycine Decreases Oxidative Stress and Improves in Vitro Development of Bovine Embryos. J. Reprod. Dev. 2016, 62, 209–212. [Google Scholar] [CrossRef]
- Graber, C.D.; Goust, J.M.; Glassman, A.D.; Kendall, R.; Loadholt, C.B. Immunomodulating Properties of Dimethylglycine in Humans. J. Infect. Dis. 1981, 143, 101–105. [Google Scholar] [CrossRef]
- Reap, E.A.; Lawson, J.W. Stimulation of the Immune Response by Dimethylglycine, a Nontoxic Metabolite. J. Lab. Clin. Med. 1990, 115, 481–486. [Google Scholar] [PubMed]
- Lawson, B.R.; Belkowski, S.M.; Whitesides, J.F.; Davis, P.; Lawson, J.W. Immunomodulation of Murine Collagen-Induced Arthritis by N, N-Dimethylglycine and a Preparation of Perna Canaliculus. BMC Complement. Altern. Med. 2007, 7, 20. [Google Scholar] [CrossRef]
- Maisel-Campbell, A.L.; Ismail, A.; Reynolds, K.A.; Poon, E.; Serrano, L.; Grushchak, S.; Farid, C.; West, D.P.; Alam, M. A Systematic Review of the Safety and Effectiveness of Platelet-Rich Plasma (PRP) for Skin Aging. Arch. Dermatol. Res. 2019, 312, 301–315. [Google Scholar] [CrossRef]
- Kuraitis, D.; Rosenthal, N.; Boh, E.; McBurney, E. Macrophages in Dermatology: Pathogenic Roles and Targeted Therapeutics. Arch. Dermatol. Res. 2021, 314, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.F.S.; Pandolfo, I.L.; de Almeida Cruz, M.; Parisi, J.R.; Garcia, L.A.; Martignago, C.C.S.; Renno, A.C.M.; Vassão, P.G. Intense Pulsed Light on Skin Rejuvenation: A Systematic Review. Arch. Dermatol. Res. 2021, 314, 823–838. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Schmuth, M.; Elias, P.M. The Regulation of Permeability Barrier Homeostasis. J. Investig. Dermatol. 2007, 127, 1574–1576. [Google Scholar] [CrossRef]
- Proksch, E.; Brandner, J.M.; Jensen, J.-M. The Skin: An Indispensable Barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The Skin Aging Exposome. J. Dermatol. Sci. 2016, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Parrado, C.; Mercado-Saenz, S.; Perez-Davo, A.; Gilaberte, Y.; Gonzalez, S.; Juarranz, A. Environmental Stressors on Skin Aging. Mechanistic Insights. Front. Pharmacol. 2019, 10, 759. [Google Scholar] [CrossRef] [PubMed]
- Niehues, H.; Bouwstra, J.A.; El Ghalbzouri, A.; Brandner, J.M.; Zeeuwen, P.L.J.M.; van den Bogaard, E.H. 3D Skin Models for 3R Research: The Potential of 3D Reconstructed Skin Models to Study Skin Barrier Function. Exp. Dermatol. 2018, 27, 501–511. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, X.; Paus, R.; Lu, Z. The Renaissance of Human Skin Organ Culture: A Critical Reappraisal. Differentiation 2018, 104, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Boarder, E.; Rumberger, B.; Howell, M.D. Modeling Skin Inflammation Using Human In Vitro Models. Curr. Protoc. 2021, 1, e72. [Google Scholar] [CrossRef]
- Sarama, R.; Matharu, P.K.; Abduldaiem, Y.; Corrêa, M.P.; Gil, C.D.; Greco, K.V. In Vitro Disease Models for Understanding Psoriasis and Atopic Dermatitis. Front. Bioeng. Biotechnol. 2022, 10, 803218. [Google Scholar] [CrossRef]
- Misiura, M.; Baszanowska, W.; Ościłowska, I.; Pałka, J.; Miltyk, W. Prolidase Stimulates Proliferation and Migration through Activation of the PI3K/Akt/mTOR Signaling Pathway in Human Keratinocytes. Int. J. Mol. Sci. 2020, 21, 9243. [Google Scholar] [CrossRef]
- Cataldi, S.; Ceccarini, M.R.; Patria, F.; Beccari, T.; Mandarano, M.; Ferri, I.; Lazzarini, A.; Curcio, F.; Albi, E. The Effect of Vitamin D3 and Silver Nanoparticles on HaCaT Cell Viability. Int. J. Mol. Sci. 2022, 23, 1410. [Google Scholar] [CrossRef] [PubMed]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef]
- Seo, G.Y.; Hyun, C.; Koh, D.; Park, S.; Lim, Y.; Kim, Y.M.; Cho, M. A Novel Synthetic Material, BMM, Accelerates Wound Repair by Stimulating Re-Epithelialization and Fibroblast Activation. Int. J. Mol. Sci. 2018, 19, 1164. [Google Scholar] [CrossRef]
- Sim, P.; Song, Y.; Yang, G.N.; Cowin, A.J.; Garg, S. In Vitro Wound Healing Properties of Novel Acidic Treatment Regimen in Enhancing Metabolic Activity and Migration of Skin Cells. Int. J. Mol. Sci. 2022, 23, 7188. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Won, K.-J.; Lee, R.; Cho, H.-S.; Hwang, S.-H.; Nah, S.-Y. Wound Healing Effect of Gintonin Involves Lysophosphatidic Acid Receptor/Vascular Endothelial Growth Factor Signaling Pathway in Keratinocytes. Int. J. Mol. Sci. 2021, 22, 10155. [Google Scholar] [CrossRef] [PubMed]
- Węgłowska, E.; Koziołkiewicz, M.; Kamińska, D.; Grobelski, B.; Pawełczak, D.; Kołodziejczyk, M.; Bielecki, S.; Gendaszewska-Darmach, E. Extracellular Nucleotides Affect the Proangiogenic Behavior of Fibroblasts, Keratinocytes, and Endothelial Cells. Int. J. Mol. Sci. 2021, 23, 238. [Google Scholar] [CrossRef]
- Liu, J.-X.; Cui, D.-L.; Yang, D.-L.; Li, J.-Y.; Yang, Z.-Y.; Su, J.-Z.; Ren, C.-X.; Niu, Y.-Y.; Xiang, P. Organophosphorus Flame Retardant TCPP Induces Cellular Senescence in Normal Human Skin Keratinocytes: Implication for Skin Aging. Int. J. Mol. Sci. 2022, 23, 14306. [Google Scholar] [CrossRef]
- Li, L.; Ngo, H.T.T.; Hwang, E.; Wei, X.; Liu, Y.; Liu, J.; Yi, T.-H. Conditioned Medium from Human Adipose-Derived Mesenchymal Stem Cell Culture Prevents UVB-Induced Skin Aging in Human Keratinocytes and Dermal Fibroblasts. Int. J. Mol. Sci. 2019, 21, 49. [Google Scholar] [CrossRef] [PubMed]
- Tortolani, D.; Di Meo, C.; Standoli, S.; Ciaramellano, F.; Kadhim, S.; Hsu, E.; Rapino, C.; Maccarrone, M. Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 2721. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.-W.; Lee, H.-J.; Youk, Y.-M.; Nam, G.-H.; Kim, Y.-M. Torilis Japonica Extract Suppresses the Induction of Atopic Inflammation. Int. J. Mol. Sci. 2023, 24, 2102. [Google Scholar] [CrossRef]
- Min, S.-Y.; Park, C.-H.; Yu, H.-W.; Park, Y.-J. Anti-Inflammatory and Anti-Allergic Effects of Saponarin and Its Impact on Signaling Pathways of RAW 264.7, RBL-2H3, and HaCaT Cells. Int. J. Mol. Sci. 2021, 22, 8431. [Google Scholar] [CrossRef]
- Zhou, M.-W.; Jiang, R.-H.; Kim, K.-D.; Lee, J.-H.; Kim, C.-D.; Yin, W.-T.; Lee, J.-H. Rosmarinic Acid Inhibits poly(I:C)-Induced Inflammatory Reaction of Epidermal Keratinocytes. Life Sci. 2016, 155, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Jung, K.E.; Yim, S.H.; Hong, D.K.; Kim, C.D.; Hong, J.Y.; Lee, H.J.; Lee, S.Y.; Kim, J.E.; Park, C.W. Autophagy Suppresses Toll-Like Receptor 3-Mediated Inflammatory Reaction in Human Epidermal Keratinocytes. Biomed Res. Int. 2020, 2020, 4584626. [Google Scholar] [CrossRef]
- Petrosino, S.; Verde, R.; Vaia, M.; Allarà, M.; Iuvone, T.; Di Marzo, V. Anti-Inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis. J. Pharmacol. Exp. Ther. 2018, 365, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Liu, J.; Wang, J.; Luo, Q.; Zhang, H.; Liu, B.; Xu, F.; Pang, Q.; Liu, Y.; Dong, J. Icariin Inhibits TNF-α/IFN-γ Induced Inflammatory Response via Inhibition of the Substance P and p38-MAPK Signaling Pathway in Human Keratinocytes. Int. Immunopharmacol. 2015, 29, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, S.K.; Bae, S.; Kim, H.; Park, Y.; Chu, N.K.; Kim, S.G.; Kim, H.-R.; Hwang, Y.-I.; Kang, J.S.; et al. The Anti-Inflammatory Effect of Alloferon on UVB-Induced Skin Inflammation through the down-Regulation of pro-Inflammatory Cytokines. Immunol. Lett. 2012, 149, 110–118. [Google Scholar] [CrossRef]
- Tang, S.-C.; Liao, P.-Y.; Hung, S.-J.; Ge, J.-S.; Chen, S.-M.; Lai, J.-C.; Hsiao, Y.-P.; Yang, J.-H. Topical Application of Glycolic Acid Suppresses the UVB Induced IL-6, IL-8, MCP-1 and COX-2 Inflammation by Modulating NF-κB Signaling Pathway in Keratinocytes and Mice Skin. J. Dermatol. Sci. 2017, 86, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, O.; Joung, D.; Mun, S.; Seo, Y.; Cha, M.; Ryu, S.; Shin, D.; Kwon, D. Anti-Inflammatory Effects of Tectroside on UVB-Induced HaCaT Cells. Int. J. Mol. Med. 2013, 31, 1471–1476. [Google Scholar] [CrossRef]
- Belső, N.; Gubán, B.; Manczinger, M.; Kormos, B.; Bebes, A.; Németh, I.; Veréb, Z.; Széll, M.; Kemény, L.; Bata-Csörgő, Z. Differential Role of D Cyclins in the Regulation of Cell Cycle by Influencing Ki67 Expression in HaCaT Cells. Exp. Cell Res. 2018, 374, 290–303. [Google Scholar] [CrossRef]
- Afnan, Q.; Kaiser, P.J.; Rafiq, R.A.; Nazir, L.A.; Bhushan, S.; Bhardwaj, S.C.; Sandhir, R.; Tasduq, S.A. Glycyrrhizic Acid Prevents Ultraviolet-B-Induced Photodamage: A Role for Mitogen-Activated Protein Kinases, Nuclear Factor Kappa B and Mitochondrial Apoptotic Pathway. Exp. Dermatol. 2016, 25, 440–446. [Google Scholar] [CrossRef]
- Fang, J.; Sun, Q.; Wang, Z.; Song, Z.; Geng, J.; Wang, C.; Li, M.; Wang, D. Enhancement of Human Epidermal Cell Defense against UVB Damage by Fermentation of Passiflora Edulis Sims Peel with Saccharomyces Cerevisiae. Nutrients 2023, 15, 501. [Google Scholar] [CrossRef]
- Quan, W.-Y.; Fan, L.-H.; Li, Q.; Lin, C.-M.; Liang, Y.-Y.; Zhang, Q.; Ye, H.; Li, X.-J.; Wu, K.-F.; Zhu, Y.-Z. DHPW1 Attenuation of UVB-Induced Skin Photodamage in Human Immortalized Keratinocytes. Exp. Gerontol. 2022, 166, 111897. [Google Scholar] [CrossRef]
- Aghaei, S.; Nilforoushzadeh, M.A.; Aghaei, M. The Role of Peroxisome Proliferator-Activated Receptor-Coactivator-1 Gene in Skin Aging. J. Res. Med. Sci. 2016, 21, 36. [Google Scholar] [CrossRef]
- Lee, S.-H.; Lee, J.-H.; Lee, H.-Y.; Min, K.-J. Sirtuin Signaling in Cellular Senescence and Aging. BMB Rep. 2019, 52, 24–34. [Google Scholar] [CrossRef]
- Bielach-Bazyluk, A.; Zbroch, E.; Mysliwiec, H.; Rydzewska-Rosolowska, A.; Kakareko, K.; Flisiak, I.; Hryszko, T. Sirtuin 1 and Skin: Implications in Intrinsic and Extrinsic Aging-A Systematic Review. Cells 2021, 10, 813. [Google Scholar] [CrossRef]
- Keren, A.; Bertolini, M.; Keren, Y.; Ullmann, Y.; Paus, R.; Gilhar, A. Human Organ Rejuvenation by VEGF-A: Lessons from the Skin. Sci. Adv. 2022, 8, eabm6756. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Kang, B.Y.; Wong, C.C.; Li, A.W.; Naseer, N.; Ibrahim, S.A.; Keimig, E.L.; Poon, E.; Alam, M. A Systematic Review of Autologous Adipose-Derived Stromal Vascular Fraction (SVF) for the Treatment of Acute Cutaneous Wounds. Arch. Dermatol. Res. 2021, 314, 417–425. [Google Scholar] [CrossRef]
- Machado Velho, J.C.; França, T.A.; Malagutti-Ferreira, M.J.; Albuquerque, E.R.; Lívero, F.A.D.R.; Soares, M.R.; Soares, A.E.E.; Ribeiro-Paes, J.T. Use of Propolis for Skin Wound Healing: Systematic Review and Meta-Analysis. Arch. Dermatol. Res. 2023, 315, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.E.; Wilgus, T.A. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Hedayatyanfard, K.; Haddadi, N.-S.; Ziai, S.A.; Karim, H.; Niazi, F.; Steckelings, U.M.; Habibi, B.; Modarressi, A.; Dehpour, A.-R. The Renin-Angiotensin System in Cutaneous Hypertrophic Scar and Keloid Formation. Exp. Dermatol. 2020, 29, 902–909. [Google Scholar] [CrossRef]
- Luengas-Martinez, A.; Hardman-Smart, J.; Rutkowski, D.; Purba, T.S.; Paus, R.; Young, H.S. Vascular Endothelial Growth Factor Blockade Induces Dermal Endothelial Cell Apoptosis in a Clinically Relevant Skin Organ Culture Model. Skin Pharmacol. Physiol. 2020, 33, 110–118. [Google Scholar] [CrossRef]
- Wei, Y.; Li, J.; Huang, Y.; Lei, X.; Zhang, L.; Yin, M.; Deng, J.; Wang, X.; Fu, X.; Wu, J. The Clinical Effectiveness and Safety of Using Epidermal Growth Factor, Fibroblast Growth Factor and Granulocyte-Macrophage Colony Stimulating Factor as Therapeutics in Acute Skin Wound Healing: A Systematic Review and Meta-Analysis. Burns Trauma 2022, 10, tkac002. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jia, L.; Zheng, H.; Feng, J.; Wei, S.; Li, J.; Cui, J.; Chen, F. The Stimulation of Macrophages by Systematical Administration of GM-CSF Can Accelerate Adult Wound Healing Process. Int. J. Mol. Sci. 2022, 23, 11287. [Google Scholar] [CrossRef]
- Kiani, M.T.; Higgins, C.A.; Almquist, B.D. The Hair Follicle: An Underutilized Source of Cells and Materials for Regenerative Medicine. ACS Biomater. Sci. Eng. 2017, 4, 1193–1207. [Google Scholar] [CrossRef]
- Morgun, E.I.; Vorotelyak, E.A. Epidermal Stem Cells in Hair Follicle Cycling and Skin Regeneration: A View from the Perspective of Inflammation. Front. Cell Dev. Biol. 2020, 8, 581697. [Google Scholar] [CrossRef]
- Premanand, A.; Reena Rajkumari, B. Androgen Modulation of Wnt/β-Catenin Signaling in Androgenetic Alopecia. Arch. Dermatol. Res. 2018, 310, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Lachgar, S.; Moukadiri, H.; Jonca, F.; Charveron, M.; Bouhaddioui, N.; Gall, Y.; Bonafe, J.L.; Plouët, J. Vascular Endothelial Growth Factor Is an Autocrine Growth Factor for Hair Dermal Papilla Cells. J. Investig. Dermatol. 1996, 106, 17–23. [Google Scholar] [CrossRef]
- Li, W.; Man, X.-Y.; Li, C.-M.; Chen, J.-Q.; Zhou, J.; Cai, S.-Q.; Lu, Z.-F.; Zheng, M. VEGF Induces Proliferation of Human Hair Follicle Dermal Papilla Cells through VEGFR-2-Mediated Activation of ERK. Exp. Cell Res. 2012, 318, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Park, S.; Choi, D.; Son, B.; Park, T. Activation of cAMP Signaling in Response to α-Phellandrene Promotes Vascular Endothelial Growth Factor Levels and Proliferation in Human Dermal Papilla Cells. Int. J. Mol. Sci. 2022, 23, 8959. [Google Scholar] [CrossRef]
- Messenger, A.G.; Rundegren, J. Minoxidil: Mechanisms of Action on Hair Growth. Br. J. Dermatol. 2004, 150, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Takai, T.; Le, T.A.; Kamijo, S.; Wang, X.L.; Ushio, H.; Hara, M.; Kawasaki, J.; Vu, A.T.; Ogawa, T.; et al. Cytokine Milieu Modulates Release of Thymic Stromal Lymphopoietin from Human Keratinocytes Stimulated with Double-Stranded RNA. J. Allergy Clin. Immunol. 2008, 123, 179–186. [Google Scholar] [CrossRef]
- McKenzie, R.C.; Sabin, E. Aberrant Signalling and Transcription Factor Activation as an Explanation for the Defective Growth Control and Differentiation of Keratinocytes in Psoriasis: A Hypothesis. Exp. Dermatol. 2003, 12, 337–345. [Google Scholar] [CrossRef]
- Tu, C.-L.; Bikle, D.D. Role of the Calcium-Sensing Receptor in Calcium Regulation of Epidermal Differentiation and Function. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 415–427. [Google Scholar] [CrossRef]
- Elsholz, F.; Harteneck, C.; Muller, W.; Friedland, K. Calcium—A Central Regulator of Keratinocyte Differentiation in Health and Disease. Eur. J. Dermatol. 2014, 24, 650–661. [Google Scholar] [CrossRef]
- Rinnerthaler, M.; Streubel, M.K.; Bischof, J.; Richter, K. Skin Aging, Gene Expression and Calcium. Exp. Gerontol. 2014, 68, 59–65. [Google Scholar] [CrossRef]
- Rinnerthaler, M.; Richter, K. The Influence of Calcium on the Skin pH and Epidermal Barrier during Aging. Curr. Probl. Dermatol. 2018, 54, 79–86. [Google Scholar] [CrossRef]
- King, K.E.; George, A.L.; Sakakibara, N.; Mahmood, K.; Moses, M.A.; Weinberg, W.C. Intersection of the p63 and NF-κB Pathways in Epithelial Homeostasis and Disease. Mol. Carcinog. 2019, 58, 1571–1580. [Google Scholar] [CrossRef]
- Bikle, D.D. Role of Vitamin D and Calcium Signaling in Epidermal Wound Healing. J. Endocrinol. Investig. 2022, 46, 205–212. [Google Scholar] [CrossRef]
- Papp, H.; Czifra, G.; Lázár, J.; Gönczi, M.; Csernoch, L.; Kovács, L.; Bíro, T. Protein Kinase C Isozymes Regulate Proliferation and High Cell Density-Mediated Differentiation in HaCaT Keratinocytes. Exp. Dermatol. 2003, 12, 811–824. [Google Scholar] [CrossRef]
- Oláh, A.; Szabó-Papp, J.; Soeberdt, M.; Knie, U.; Dähnhardt-Pfeiffer, S.; Abels, C.; Bíró, T. Echinacea Purpurea-Derived Alkylamides Exhibit Potent Anti-Inflammatory Effects and Alleviate Clinical Symptoms of Atopic Eczema. J. Dermatol. Sci. 2017, 88, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Min, G.-Y.; Kim, J.-H.; Kim, T.-I.; Cho, W.-K.; Yang, J.-H.; Ma, J.-Y. Indigo Pulverata Levis (Chung-Dae, Persicaria Tinctoria) Alleviates Atopic Dermatitis-like Inflammatory Responses In Vivo and In Vitro. Int. J. Mol. Sci. 2022, 23, 553. [Google Scholar] [CrossRef] [PubMed]
- Oláh, A.; Markovics, A.; Szabó-Papp, J.; Szabó, P.T.; Stott, C.; Zouboulis, C.C.; Bíró, T. Differential Effectiveness of Selected Non-Psychotropic Phytocannabinoids on Human Sebocyte Functions Implicates Their Introduction in Dry/seborrhoeic Skin and Acne Treatment. Exp. Dermatol. 2016, 25, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Markovics, A.; Tóth, K.F.; Sós, K.E.; Magi, J.; Gyöngyösi, A.; Benyó, Z.; Zouboulis, C.C.; Bíró, T.; Oláh, A. Nicotinic Acid Suppresses Sebaceous Lipogenesis of Human Sebocytes via Activating Hydroxycarboxylic Acid Receptor 2 (HCA(2)). J. Cell. Mol. Med. 2019, 23, 6203–6214. [Google Scholar] [CrossRef]
- Angyal, Á.; Pénzes, Z.; Alimohammadi, S.; Horváth, D.; Takács, L.; Vereb, G.; Zsebik, B.; Bíró, T.; Tóth, K.F.; Lisztes, E.; et al. Anandamide Concentration-Dependently Modulates Toll-Like Receptor 3 Agonism or UVB-Induced Inflammatory Response of Human Corneal Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 7776. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Zákány, N.; Tiede, S.; Purba, T.; Harries, M.; Tsuruta, D.; Bíró, T.; Paus, R. Human Epithelial Stem Cell Survival within Their Niche Requires “Tonic” Cannabinoid Receptor 1-Signalling-Lessons from the Hair Follicle. Exp. Dermatol. 2021, 30, 479–493. [Google Scholar] [CrossRef]
- Stessuk, T.; Puzzi, M.B.; Chaim, E.A.; Alves, P.C.M.; de Paula, E.V.; Forte, A.; Izumizawa, J.M.; Oliveira, C.C.; Frei, F.; Ribeiro-Paes, J.T. Platelet-Rich Plasma (PRP) and Adipose-Derived Mesenchymal Stem Cells: Stimulatory Effects on Proliferation and Migration of Fibroblasts and Keratinocytes In Vitro. Arch. Dermatol. Res. 2016, 308, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-H.; An, H.-J.; Kim, J.-Y.; Gwon, M.-G.; Gu, H.; Lee, S.-J.; Park, J.Y.; Park, K.-D.; Han, S.-M.; Kim, M.-K.; et al. Apamin Inhibits TNF-α- and IFN-γ-Induced Inflammatory Cytokines and Chemokines via Suppressions of NF-κB Signaling Pathway and STAT in Human Keratinocytes. Pharmacol. Rep. 2017, 69, 1030–1035. [Google Scholar] [CrossRef]
- Vasas, N.; Pénzes, Z.; Kistamás, K.; Nánási, P.P.; Molnár, S.; Szegedi, A.; Szöllősi, A.G.; Bíró, T. Transient Receptor Potential Vanilloid 3 Expression Is Increased in Non-Lesional Skin of Atopic Dermatitis Patients. Exp. Dermatol. 2022, 31, 807–813. [Google Scholar] [CrossRef]
- Hong, S.; Kim, E.-Y.; Lim, S.-E.; Kim, J.-H.; Sohn, Y.; Jung, H.-S. Dendrobium Nobile Lindley Administration Attenuates Atopic Dermatitis-like Lesions by Modulating Immune Cells. Int. J. Mol. Sci. 2022, 23, 4470. [Google Scholar] [CrossRef] [PubMed]
- Szöllősi, A.G.; Vasas, N.; Angyal, Á.; Kistamás, K.; Nánási, P.P.; Mihály, J.; Béke, G.; Herczeg-Lisztes, E.; Szegedi, A.; Kawada, N.; et al. Activation of TRPV3 Regulates Inflammatory Actions of Human Epidermal Keratinocytes. J. Investig. Dermatol. 2017, 138, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, L.; Kelemen, B.; Szabó, T.; Bíró, T.; Tóth, B.I. Human Podocytes Express Functional Thermosensitive TRPV Channels. Br. J. Pharmacol. 2017, 174, 4493–4507. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lendvai, A.; Béke, G.; Hollósi, E.; Becker, M.; Völker, J.M.; Schulze zur Wiesche, E.; Bácsi, A.; Bíró, T.; Mihály, J. N,N-Dimethylglycine Sodium Salt Exerts Marked Anti-Inflammatory Effects in Various Dermatitis Models and Activates Human Epidermal Keratinocytes by Increasing Proliferation, Migration, and Growth Factor Release. Int. J. Mol. Sci. 2023, 24, 11264. https://doi.org/10.3390/ijms241411264
Lendvai A, Béke G, Hollósi E, Becker M, Völker JM, Schulze zur Wiesche E, Bácsi A, Bíró T, Mihály J. N,N-Dimethylglycine Sodium Salt Exerts Marked Anti-Inflammatory Effects in Various Dermatitis Models and Activates Human Epidermal Keratinocytes by Increasing Proliferation, Migration, and Growth Factor Release. International Journal of Molecular Sciences. 2023; 24(14):11264. https://doi.org/10.3390/ijms241411264
Chicago/Turabian StyleLendvai, Alexandra, Gabriella Béke, Erika Hollósi, Maike Becker, Jörn Michael Völker, Erik Schulze zur Wiesche, Attila Bácsi, Tamás Bíró, and Johanna Mihály. 2023. "N,N-Dimethylglycine Sodium Salt Exerts Marked Anti-Inflammatory Effects in Various Dermatitis Models and Activates Human Epidermal Keratinocytes by Increasing Proliferation, Migration, and Growth Factor Release" International Journal of Molecular Sciences 24, no. 14: 11264. https://doi.org/10.3390/ijms241411264
APA StyleLendvai, A., Béke, G., Hollósi, E., Becker, M., Völker, J. M., Schulze zur Wiesche, E., Bácsi, A., Bíró, T., & Mihály, J. (2023). N,N-Dimethylglycine Sodium Salt Exerts Marked Anti-Inflammatory Effects in Various Dermatitis Models and Activates Human Epidermal Keratinocytes by Increasing Proliferation, Migration, and Growth Factor Release. International Journal of Molecular Sciences, 24(14), 11264. https://doi.org/10.3390/ijms241411264