
Rapid Generation and Molecular Docking Analysis of Single-Chain Fragment Variable (scFv) Antibody Selected by Ribosome Display Targeting Cholecystokinin B Receptor (CCK-BR) for Reduction of Chronic Neuropathic Pain
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
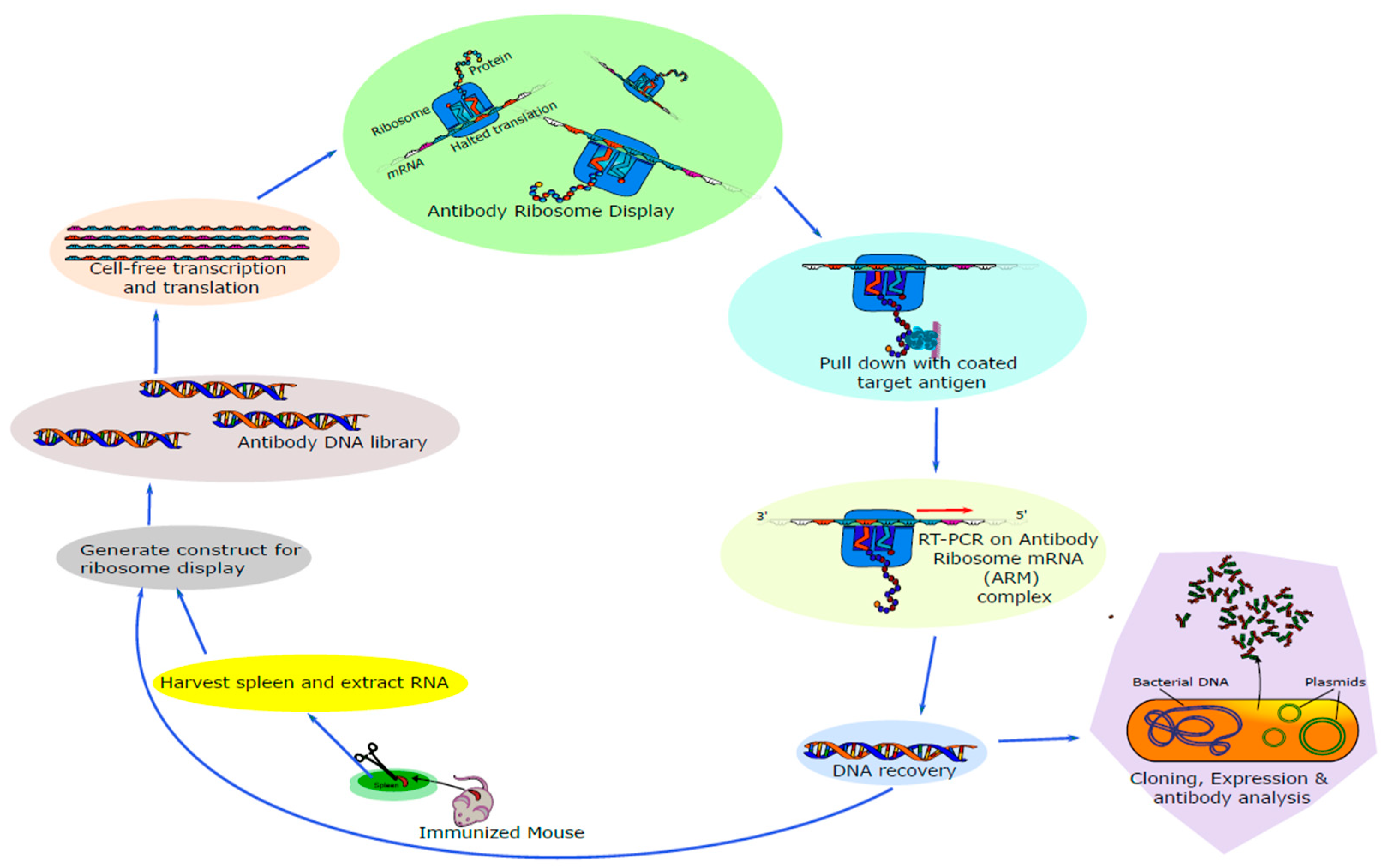
2.1. Anti-CCK-B scFv Antibodies Generated from Cell-Free Ribosomal Display
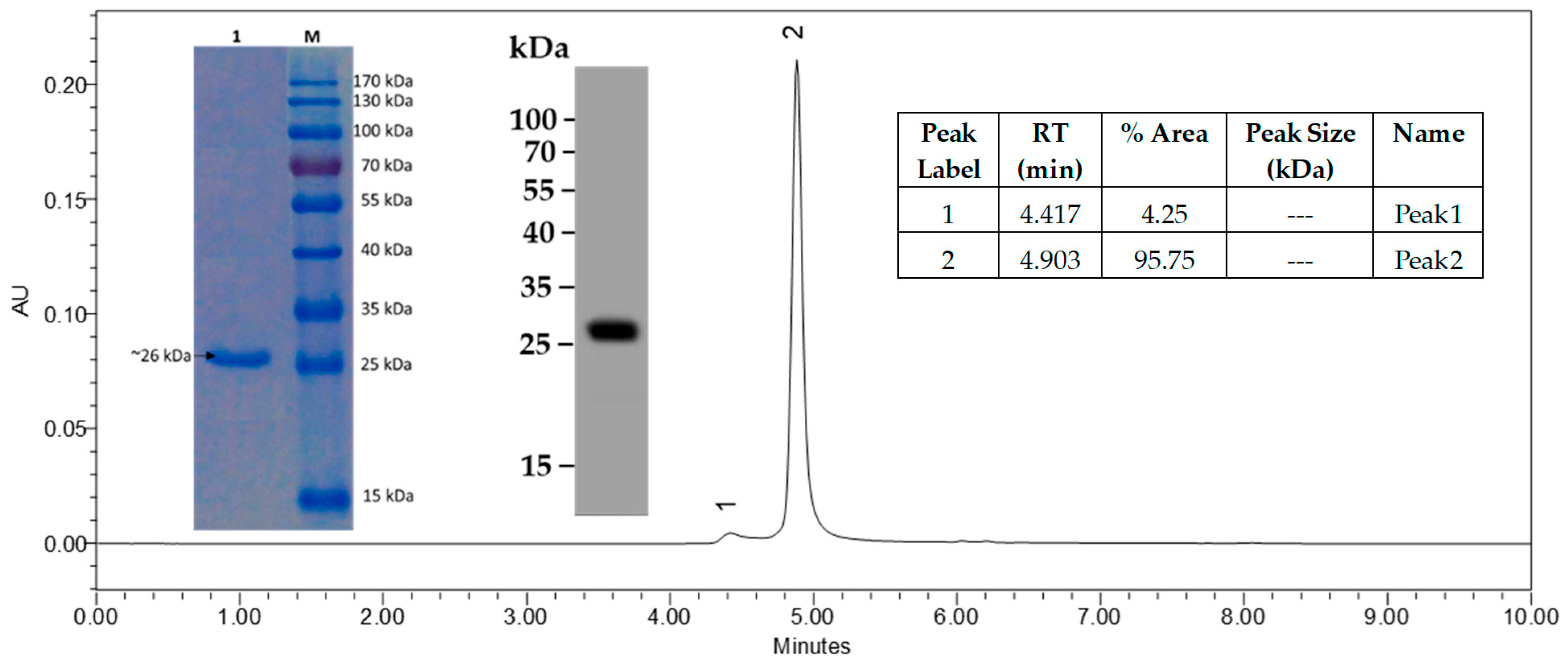
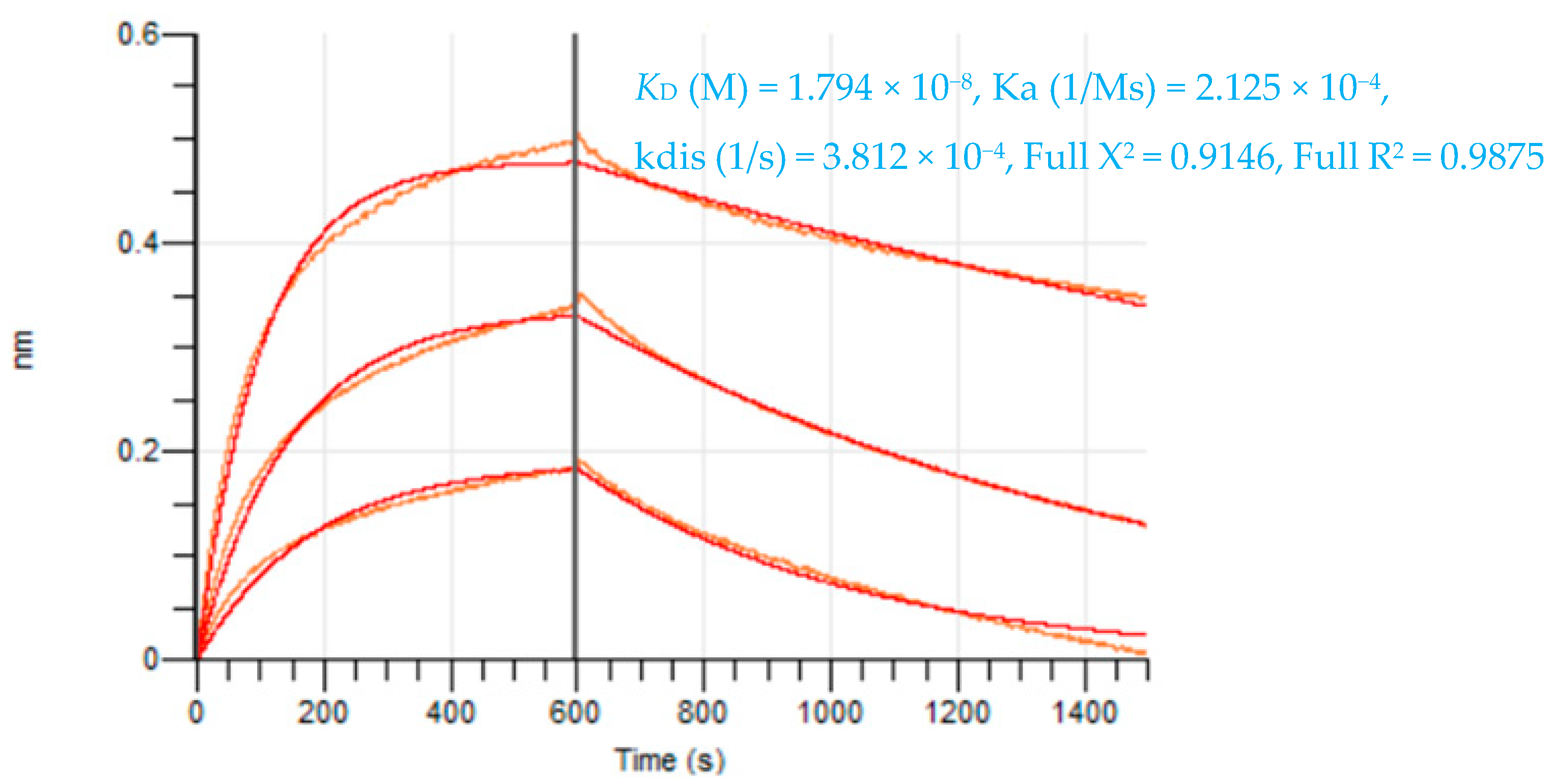
2.1.1. Characteristics of the Lead CCKBR scFv 77-2
2.1.2. Protein Surface Analysis
2.1.3. Prediction of scFv77-2 Interaction with CCK-BR Peptide
2.2. In Vivo Validation
2.2.1. Selection of the Lead CCK-BR scFv
2.2.2. CCK-BR scFv for Evoked Hypersensitivity in Male and Female Mice
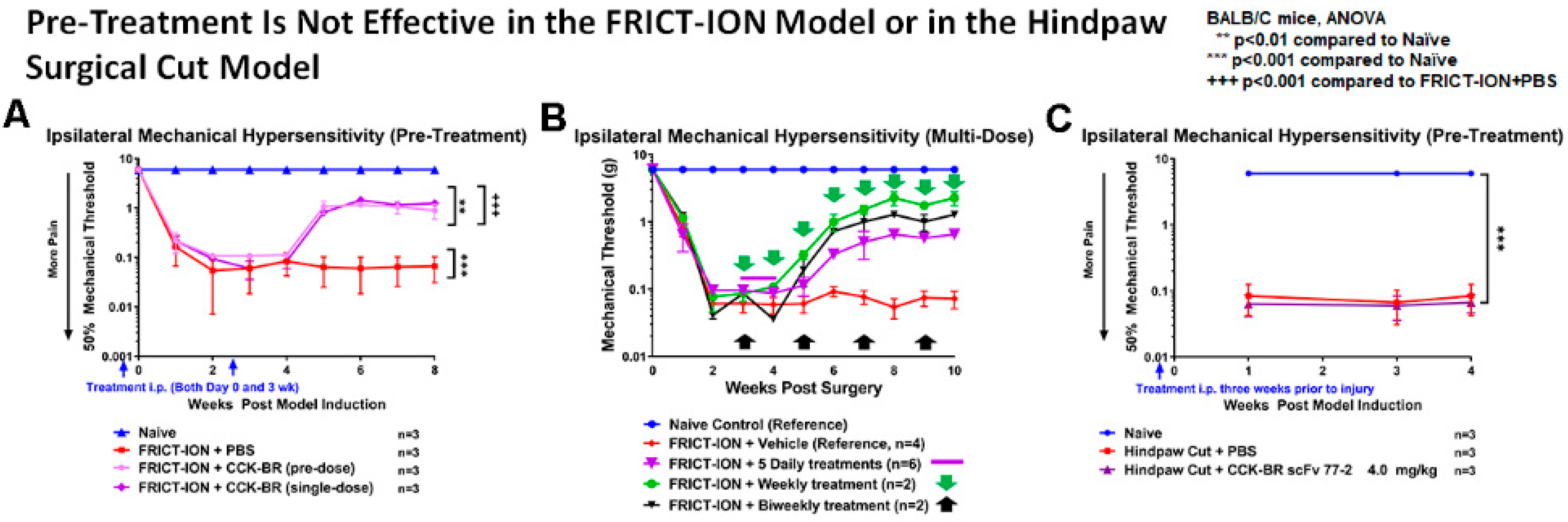
2.2.3. Pre-Treatment Is Not Efficacious
2.2.4. CCKB Pharmacologic Comparator LY225910
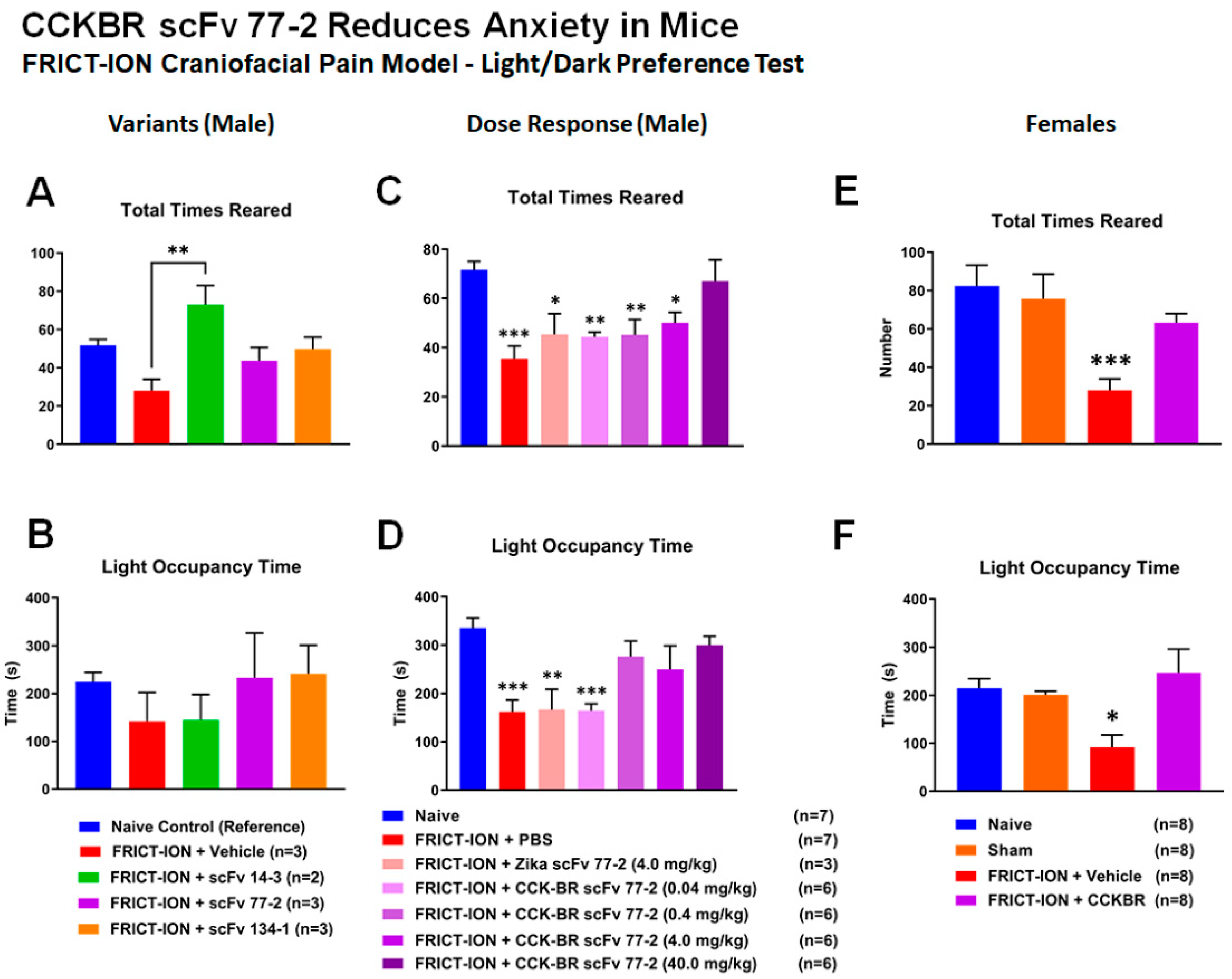
2.2.5. Efficacy of CCK-BR scFv for Prevention of Pain-Related Anxiety
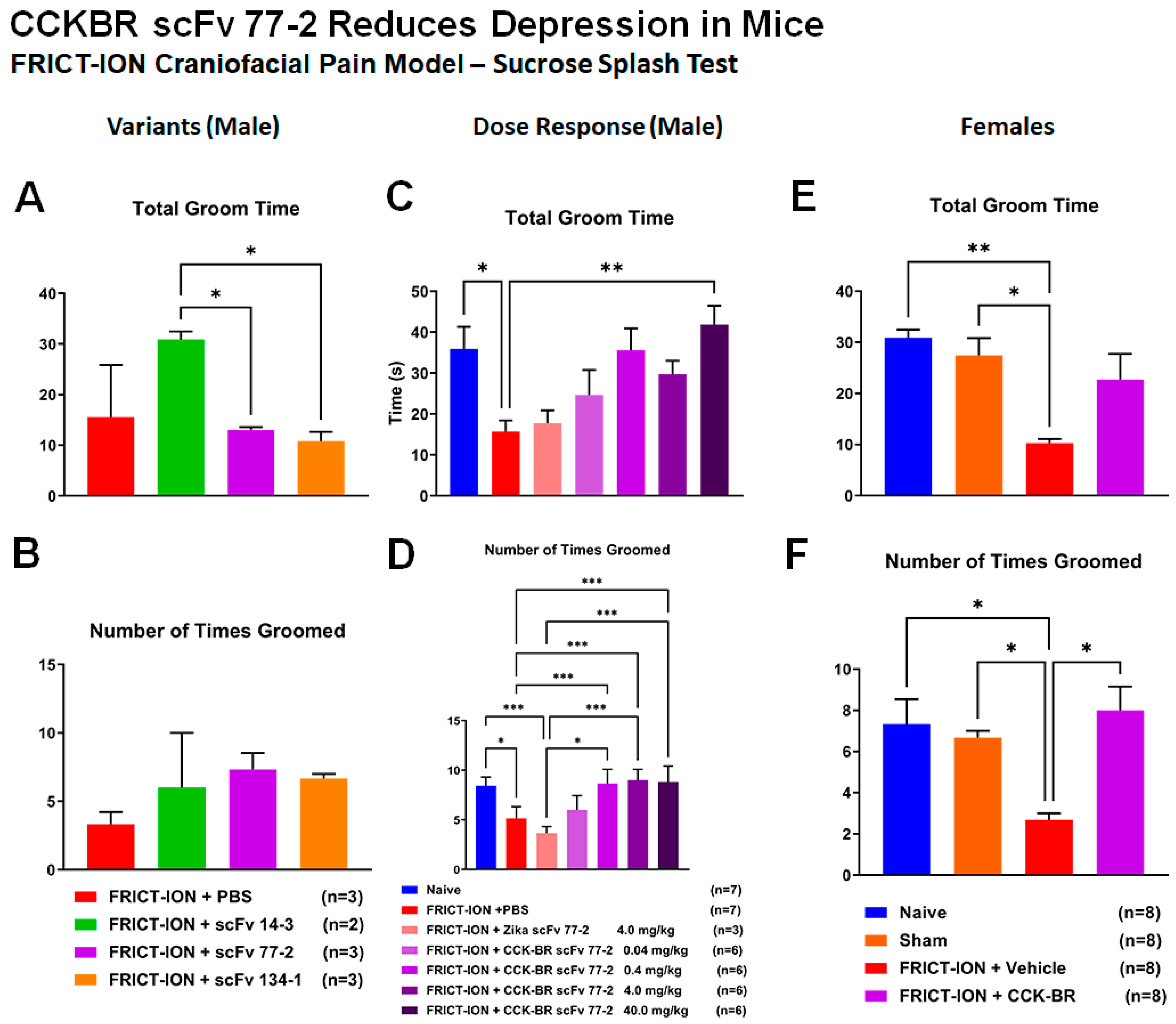
2.2.6. Efficacy of CCK-BR scFv for Prevention of Depression
2.2.7. Preliminary Demonstration of Brain Penetrance
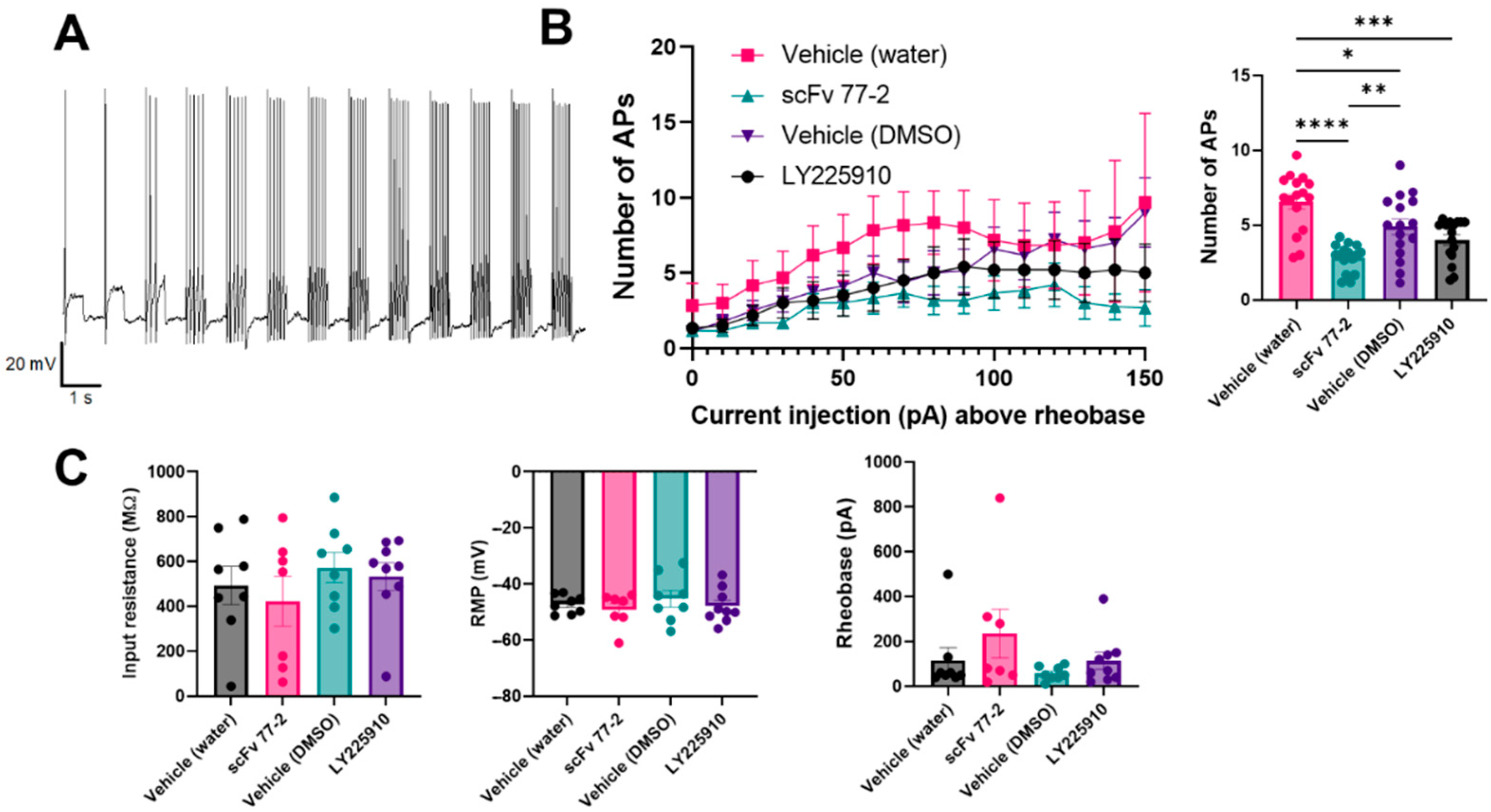
2.2.8. Murine CCKBR scFv 77-2 Reduces DRG Neuron Excitability
3. Discussion
3.1. Generation of CCKBR scFv
3.2. In Vivo Efficacy of CCKBR scFv
3.3. Efficacy of CCKBR scFv on Mechanical and Cold Hypersensitivity
3.4. Efficacy for Anxiety- and Depression-like Behavior
3.5. Effect of scFv 77-2 on DRG Neuron Excitability
4. Materials and Methods
4.1. Generation of Cholecystokinin B (CCK-BR) scFvs
4.2. Protein Surface Analysis
4.3. Computational Modeling
4.3.1. Homology Modeling
4.3.2. Molecular Docking
4.3.3. Molecular Dynamics (MD) Simulations
4.4. Surgical Induction of the Trigeminal Neuropathic Pain Model
4.5. Lead CCK-BR scFv Determination
4.6. In Vivo Behavioral Read-Outs
4.6.1. Von Frey Fiber Assessment of Hypersensitivity
4.6.2. Cognition Dependent Anxiety-like and Depression-like Behavioral Tests
4.6.3. Light/Dark Place Preference Test
4.6.4. Elevated Zero Maze
4.6.5. Sucrose Splash Test
4.7. Western Blot
4.8. Dorsal Root Ganglion Cultures
4.9. Whole-Cell Patch Clamp Electrophysiology
4.10. Statistical and Data Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayoub, M.A.; Crepieux, P.; Koglin, M.; Parmentier, M.; Pin, J.P.; Poupon, A.; Reiter, E.; Smit, M.; Steyaert, J.; Watier, H.; et al. Antibodies targeting G protein-coupled receptors: Recent advances and therapeutic challenges. MAbs 2017, 9, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Butler, D.C.; McLear, J.A.; Messer, A. Engineered antibody therapies to counteract mutant huntingtin and related toxic intracellular proteins. Prog. Neurobiol. 2012, 97, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Škrlj, N.; Dolinar, M. New engineered antibodies against prions. Bioengineered 2014, 5, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Angelini, A.; Miyabe, Y.; Newsted, D.; Kwan, B.H.; Miyabe, C.; Kelly, R.L.; Jamy, M.N.; Luster, A.D.; Wittrup, K.D. Directed evolution of broadly crossreactive chemokine-blocking antibodies efficacious in arthritis. Nat. Commun. 2018, 9, 1461. [Google Scholar] [CrossRef]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.; Hamid, M. scFv antibody: Principles and clinical application. Clin. Dev. Immunol. 2012, 2012, 980250. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, M.; Boido, V.; Colla, P.; Loddo, R.; Posocco, P.; Paneni, M.S.; Fermeglia, M.; Pricl, S. Pharmacophore modeling, resistant mutant isolation, docking, and MM-PBSA analysis: Combined experimental/computer-assisted approaches to identify new inhibitors of the bovine viral diarrhea virus (BVDV). Bioorg. Med. Chem. 2010, 18, 2304–2316. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zhang, C.; Miao, L.; Zhang, D.; Bai, Y.; Hou, C.; Liu, J.; Yan, F.; Mu, Y.; Luo, G. Triple mutated antibody scFv2F3 with high GPx activity: Insights from MD, docking, MDFE, and MM-PBSA simulation. Amino Acids 2013, 44, 1009–1019. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, C.F.; Zhao, B.B.; Gong, L.L.; Jin, W.J.; Liu, J.J.; Wang, J.F.; Wang, T.T.; Yuan, X.H.; He, Y.W. A novel antibody humanization method based on epitopes scanning and molecular dynamics simulation. PLoS ONE 2013, 8, e80636. [Google Scholar] [CrossRef]
- Rehfeld, J.F. Cholecystokinin and panic disorder--three unsettled questions. Regul. Pept. 2000, 93, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.J.; Puke, M.J.; Verge, V.M.; Wiesenfeld-Hallin, Z.; Hughes, J.; Hökfelt, T. Up-regulation of cholecystokinin in primary sensory neurons is associated with morphine insensitivity in experimental neuropathic pain in the rat. Neurosci. Lett. 1993, 152, 129–132. [Google Scholar] [CrossRef]
- Gutierrez-Mecinas, M.; Bell, A.M.; Shepherd, F.; Polgár, E.; Watanabe, M.; Furuta, T.; Todd, A.J. Expression of cholecystokinin by neurons in mouse spinal dorsal horn. J. Comp. Neurol. 2019, 527, 1857–1871. [Google Scholar] [CrossRef] [PubMed]
- Bangash, M.A.; Alles, S.R.A.; Santana-Varela, S.; Millet, Q.; Sikandar, S.; de Clauser, L.; Ter Heegde, F.; Habib, A.M.; Pereira, V.; Sexton, J.E.; et al. Distinct transcriptional responses of mouse sensory neurons in models of human chronic pain conditions. Wellcome Open Res. 2018, 3, 78. [Google Scholar] [CrossRef]
- Danaher, R.J.; Zhang, L.; Donley, C.J.; Laungani, N.A.; Hui, S.E.; Miller, C.S.; Westlund, K.N. Histone deacetylase inhibitors prevent persistent hypersensitivity in an orofacial neuropathic pain model. Mol. Pain 2018, 14, 1744806918796763. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.E.; Gebhart, G.F. Modulation of visceral hyperalgesia by morphine and cholecystokinin from the rat rostroventral medial medulla. Pain 2003, 104, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Kovelowski, C.J.; Ossipov, M.H.; Sun, H.; Lai, J.; Malan, T.P., Jr.; Porreca, F. Supraspinal cholecystokinin may drive tonic descending facilitation mechanisms to maintain neuropathic pain in the rat. Pain 2000, 87, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Cutler, N.R.; Ballenger, J.C.; Patterson, W.M.; Mendels, J.; Chenault, A.; Shrivastava, R.; Matzura-Wolfe, D.; Lines, C.; Reines, S. A placebo-controlled trial of L-365,260, a CCKB antagonist, in panic disorder. Biol. Psychiatry 1995, 37, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Bradwejn, J.; Koszycki, D.; Meterissian, G. Cholecystokinin-tetrapeptide induces panic attacks in patients with panic disorder. Can. J. Psychiatry Rev. Can. Psychiatr. 1990, 35, 83–85. [Google Scholar] [CrossRef]
- Pande, A.C.; Greiner, M.; Adams, J.B.; Lydiard, R.B.; Pierce, M.W. Placebo-controlled trial of the CCK-B antagonist, CI-988, in panic disorder. Biol. Psychiatry 1999, 46, 860–862. [Google Scholar] [CrossRef]
- Vanderah, T.W.; Lai, J.; Yamamura, H.I.; Porreca, F. Antisense oligodeoxynucleotide to the CCKB receptor produces naltrindole- and [Leu5]enkephalin antiserum-sensitive enhancement of morphine antinociception. Neuroreport 1994, 5, 2601–2605. [Google Scholar] [CrossRef] [PubMed]
- Roques, B.P.; Noble, F. Association of enkephalin catabolism inhibitors and CCK-B antagonists: A potential use in the management of pain and opioid addiction. Neurochem. Res. 1996, 21, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Wiesenfeld-Hallin, Z.; Xu, X.J. The role of cholecystokinin in nociception, neuropathic pain and opiate tolerance. Regul. Pept. 1996, 65, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Chou, J.; Iadarola, M.; Yang, H.Y.; Costa, E. Proglumide prevents and curtails acute tolerance to morphine in rats. Neuropharmacology 1984, 23, 715–718. [Google Scholar] [CrossRef]
- Watkins, L.R.; Kinscheck, I.B.; Mayer, D.J. Potentiation of opiate analgesia and apparent reversal of morphine tolerance by proglumide. Science 1984, 224, 395–396. [Google Scholar] [CrossRef]
- Watkins, L.R.; Kinscheck, I.B.; Mayer, D.J. Potentiation of morphine analgesia by the cholecystokinin antagonist proglumide. Brain Res. 1985, 327, 169–180. [Google Scholar] [CrossRef]
- Panerai, A.E.; Rovati, L.C.; Cocco, E.; Sacerdote, P.; Mantegazza, P. Dissociation of tolerance and dependence to morphine: A possible role for cholecystokinin. Brain Res. 1987, 410, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Dourish, C.T.; Hawley, D.; Iversen, S.D. Enhancement of morphine analgesia and prevention of morphine tolerance in the rat by the cholecystokinin antagonist L-364. Eur. J. Pharmacol. 1988, 147, 469–472. [Google Scholar] [CrossRef]
- Dourish, C.T.; O’Neill, M.F.; Coughlan, J.; Kitchener, S.J.; Hawley, D.; Iversen, S.D. The selective CCK-B receptor antagonist L-365,260 enhances morphine analgesia and prevents morphine tolerance in the rat. Eur. J. Pharmacol. 1990, 176, 35–44. [Google Scholar] [CrossRef]
- Hoffmann, O.; Wiesenfeld-Hallin, Z. The CCK-B receptor antagonist Cl 988 reverses tolerance to morphine in rats. Neuroreport 1994, 5, 2565–2568. [Google Scholar] [CrossRef]
- Idänpään-Heikkilä, J.J.; Guilbaud, G.; Kayser, V. Prevention of tolerance to the antinociceptive effects of systemic morphine by a selective cholecystokinin-B receptor antagonist in a rat model of peripheral neuropathy. J. Pharmacol. Exp. Ther. 1997, 282, 1366–1372. [Google Scholar]
- Kissin, I.; Bright, C.A.; Bradley, E.L., Jr. Acute tolerance to continuously infused alfentanil: The role of cholecystokinin and N-methyl-D-aspartate-nitric oxide systems. Anesth. Analg. 2000, 91, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Westlund, K.N.; Montera, M.A.; Goins, A.E.; Alles, S.R.A.; Afaghpour-Becklund, M.; Bartel, R.; Durvasula, R.; Kunamneni, A. Single-chain Fragment variable antibody targeting cholecystokinin-B receptor for pain reduction. Neurobiol. Pain 2021, 10, 100067. [Google Scholar] [CrossRef]
- Kunamneni, A.; Ye, C.; Bradfute, S.B.; Durvasula, R. Ribosome display for the rapid generation of high-affinity Zika-neutralizing single-chain antibodies. PLoS ONE 2018, 13, e0205743. [Google Scholar] [CrossRef]
- Kunamneni, A.; Clarke, E.C.; Ye, C.; Bradfute, S.B.; Durvasula, R. Generation and Selection of a Panel of Pan-Filovirus Single-Chain Antibodies using Cell-Free Ribosome Display. Am. J. Trop. Med. Hyg. 2019, 101, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Giudicelli, V.; Chaume, D.; Lefranc, M.P. IMGT/V-QUEST, an integrated software program for immunoglobulin and T cell receptor V-J and V-D-J rearrangement analysis. Nucleic Acids Res. 2004, 32, W435–W440. [Google Scholar] [CrossRef]
- Kaleli, N.E.; Karadag, M.; Kalyoncu, S. Phage display derived therapeutic antibodies have enriched aliphatic content: Insights for developability issues. Proteins 2019, 87, 607–618. [Google Scholar] [CrossRef]
- Sankar, K.; Krystek, S.R., Jr.; Carl, S.M.; Day, T.; Maier, J.K.X. AggScore: Prediction of aggregation-prone regions in proteins based on the distribution of surface patches. Proteins 2018, 86, 1147–1156. [Google Scholar] [CrossRef]
- Miller, M.S.; Douglass, J.; Hwang, M.S.; Skora, A.D.; Murphy, M.; Papadopoulos, N.; Kinzler, K.W.; Vogelstein, B.; Zhou, S.; Gabelli, S.B. An engineered antibody fragment targeting mutant β-catenin via major histocompatibility complex I neoantigen presentation. J. Biol. Chem. 2019, 294, 19322–19334. [Google Scholar] [CrossRef] [PubMed]
- Montera, M.A.; Westlund, K.N. Minimally invasive oral surgery induction of the FRICT-ION chronic neuropathic pain model. Bio-Protocol 2020, 10, e3591. [Google Scholar] [CrossRef] [PubMed]
- Obrezanova, O.; Arnell, A.; de la Cuesta, R.G.; Berthelot, M.E.; Gallagher, T.R.; Zurdo, J.; Stallwood, Y. Aggregation risk prediction for antibodies and its application to biotherapeutic development. MAbs 2015, 7, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Sarker, A.; Rathore, A.S.; Gupta, R.D. Evaluation of scFv protein recovery from E. coli by in vitro refolding and mild solubilization process. Microb. Cell Fact. 2019, 18, 5. [Google Scholar] [CrossRef]
- Haviv, Y.; Zadik, Y.; Sharav, Y.; Benoliel, R. Painful traumatic trigeminal neuropathy: An open study on the pharmacotherapeutic response to stepped treatment. J. Oral Facial Pain Headache 2014, 28, 52–60. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Han, J. Modification of opioid receptors and uncoupling of receptors from G proteins as possible mechanisms underlying suppression of opioid binding by cholecystokinin octapeptide. Chin. Med. Sci. J. 1993, 8, 1–4. [Google Scholar] [PubMed]
- Westlund, K.N.; Zhang, M. Building and Testing PPARgamma Therapeutic ELB00824 with an Improved Therapeutic Window for Neuropathic Pain. Molecules 2020, 25, 1120. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput.-Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, W.; He, B.; Walker, S.E.; Zhang, H.; Govindarajoo, B.; Virtanen, J.; Xue, Z.; Shen, H.B.; Zhang, Y. Template-based protein structure prediction in CASP11 and retrospect of I-TASSER in the last decade. Proteins 2016, 84 (Suppl. 1), 233–246. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Day, T.; Warshaviak, D.; Murrett, C.; Friesner, R.; Pearlman, D. Antibody structure determination using a combination of homology modeling, energy-based refinement, and loop prediction. Proteins 2014, 82, 1646–1655. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhang, Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 2011, 101, 2525–2534. [Google Scholar] [CrossRef]
- Zhang, J.; Liang, Y.; Zhang, Y. Atomic-level protein structure refinement using fragment-guided molecular dynamics conformation sampling. Structure 2011, 19, 1784–1795. [Google Scholar] [CrossRef]
- Chuang, G.Y.; Kozakov, D.; Brenke, R.; Comeau, S.R.; Vajda, S. DARS (Decoys As the Reference State) potentials for protein-protein docking. Biophys. J. 2008, 95, 4217–4227. [Google Scholar] [CrossRef]
- Bhachoo, J.; Beuming, T. Investigating Protein–Peptide Interactions Using the Schrödinger Computational Suite. In Modeling Peptide-Protein Interactions: Methods and Protocols; Schueler-Furman, O., London, N., Eds.; Springer: New York, NY, USA, 2017; pp. 235–254. [Google Scholar]
- Kozakov, D.; Brenke, R.; Comeau, S.R.; Vajda, S. PIPER: An FFT-based protein docking program with pairwise potentials. Proteins 2006, 65, 392–406. [Google Scholar] [CrossRef]
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L.; et al. OPLS3: A Force Field Providing Broad Coverage of Drug-like Small Molecules and Proteins. J. Chem. Theory Comput. 2016, 12, 281–296. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A Gen. Phys. 1985, 31, 1695–1697. [Google Scholar] [CrossRef]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Ikeguchi, M. Partial rigid-body dynamics in NPT, NPAT and NPgammaT ensembles for proteins and membranes. J. Comput. Chem. 2004, 25, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Ma, F.; Zhang, L.; Lyons, D.; Westlund, K.N. Orofacial neuropathic pain mouse model induced by Trigeminal Inflammatory Compression (TIC) of the infraorbital nerve. Mol Brain. 2012, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Lyons, D.N.; Kniffin, T.C.; Zhang, L.P.; Danaher, R.J.; Miller, C.S.; Bocanegra, J.L.; Carlson, C.R.; Westlund, K.N. Trigeminal Inflammatory Compression (TIC) injury induces chronic facial pain and susceptibility to anxiety-related behaviors. Neuroscience 2015, 295, 126–138. [Google Scholar] [CrossRef]
- Lyons, D.N.; Zhang, L.; Danaher, R.J.; Miller, C.S.; Westlund, K.N. PPARγ Agonists Attenuate Trigeminal Neuropathic Pain. Clin. J. Pain 2017, 33, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, I.; Bohren, Y.; Waltisperger, E.; Sage-Ciocca, D.; Yin, J.C.; Freund-Mercier, M.J.; Barrot, M. A time-dependent history of mood disorders in a murine model of neuropathic pain. Biol. Psychiatry 2011, 70, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, I.; Barthas, F.; Barrot, M. Emotional consequences of neuropathic pain: Insight from preclinical studies. Neurosci. Biobehav. Rev. 2014, 47, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Malin, S.A.; Davis, B.M.; Molliver, D.C. Production of dissociated sensory neuron cultures and considerations for their use in studying neuronal function and plasticity. Nat. Protoc. 2007, 2, 152–160. [Google Scholar] [CrossRef] [PubMed]









 daily dose given for 5 days,
daily dose given for 5 days,  once a week, or
once a week, or  every other week). (C) If scFv is given once, then hindpaw surgical cut performed three weeks later, the scFv had no effect compared to the vehicle.
daily dose given for 5 days, once a week, or every other week). (C) If scFv is given once, then hindpaw surgical cut performed three weeks later, the scFv had no effect compared to the vehicle.
every other week). (C) If scFv is given once, then hindpaw surgical cut performed three weeks later, the scFv had no effect compared to the vehicle.
daily dose given for 5 days, once a week, or every other week). (C) If scFv is given once, then hindpaw surgical cut performed three weeks later, the scFv had no effect compared to the vehicle.






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunamneni, A.; Montera, M.A.; Durvasula, R.; Alles, S.R.A.; Goyal, S.; Westlund, K.N. Rapid Generation and Molecular Docking Analysis of Single-Chain Fragment Variable (scFv) Antibody Selected by Ribosome Display Targeting Cholecystokinin B Receptor (CCK-BR) for Reduction of Chronic Neuropathic Pain. Int. J. Mol. Sci. 2023, 24, 11035. https://doi.org/10.3390/ijms241311035
Kunamneni A, Montera MA, Durvasula R, Alles SRA, Goyal S, Westlund KN. Rapid Generation and Molecular Docking Analysis of Single-Chain Fragment Variable (scFv) Antibody Selected by Ribosome Display Targeting Cholecystokinin B Receptor (CCK-BR) for Reduction of Chronic Neuropathic Pain. International Journal of Molecular Sciences. 2023; 24(13):11035. https://doi.org/10.3390/ijms241311035
Chicago/Turabian StyleKunamneni, Adinarayana, Marena A. Montera, Ravi Durvasula, Sascha R. A. Alles, Sachin Goyal, and Karin N. Westlund. 2023. "Rapid Generation and Molecular Docking Analysis of Single-Chain Fragment Variable (scFv) Antibody Selected by Ribosome Display Targeting Cholecystokinin B Receptor (CCK-BR) for Reduction of Chronic Neuropathic Pain" International Journal of Molecular Sciences 24, no. 13: 11035. https://doi.org/10.3390/ijms241311035
APA StyleKunamneni, A., Montera, M. A., Durvasula, R., Alles, S. R. A., Goyal, S., & Westlund, K. N. (2023). Rapid Generation and Molecular Docking Analysis of Single-Chain Fragment Variable (scFv) Antibody Selected by Ribosome Display Targeting Cholecystokinin B Receptor (CCK-BR) for Reduction of Chronic Neuropathic Pain. International Journal of Molecular Sciences, 24(13), 11035. https://doi.org/10.3390/ijms241311035