
Protective Effect of Amaranthus cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts



Abstract
1. Introduction
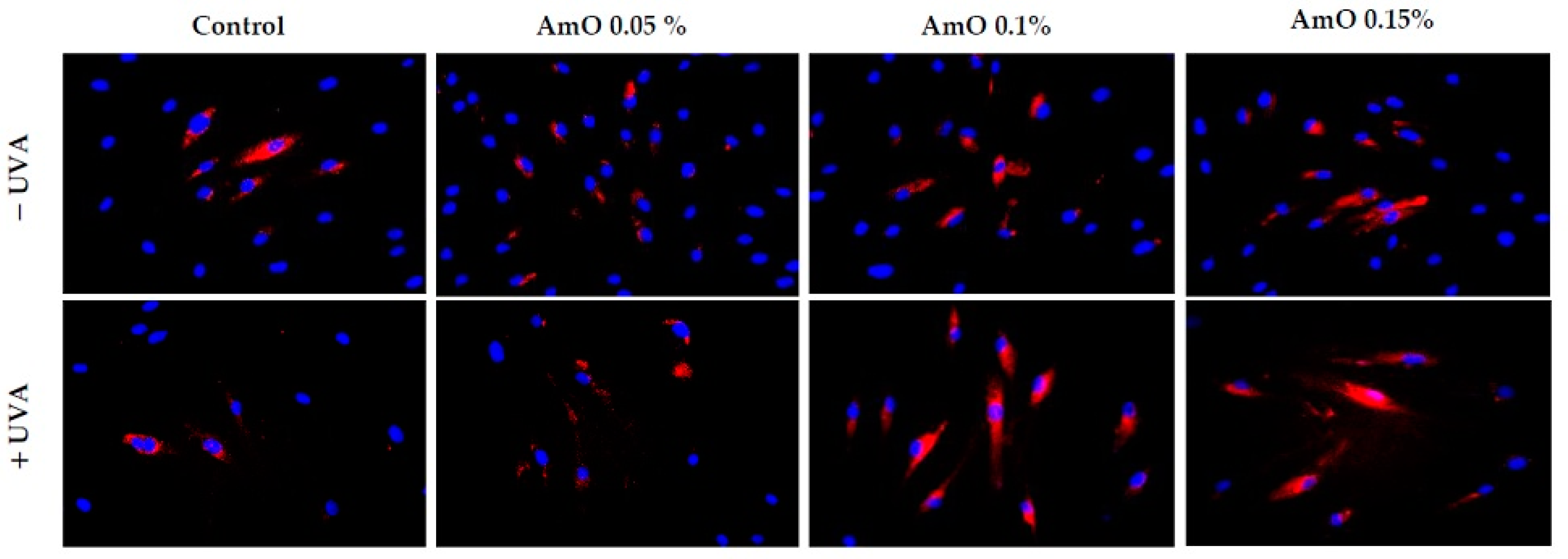
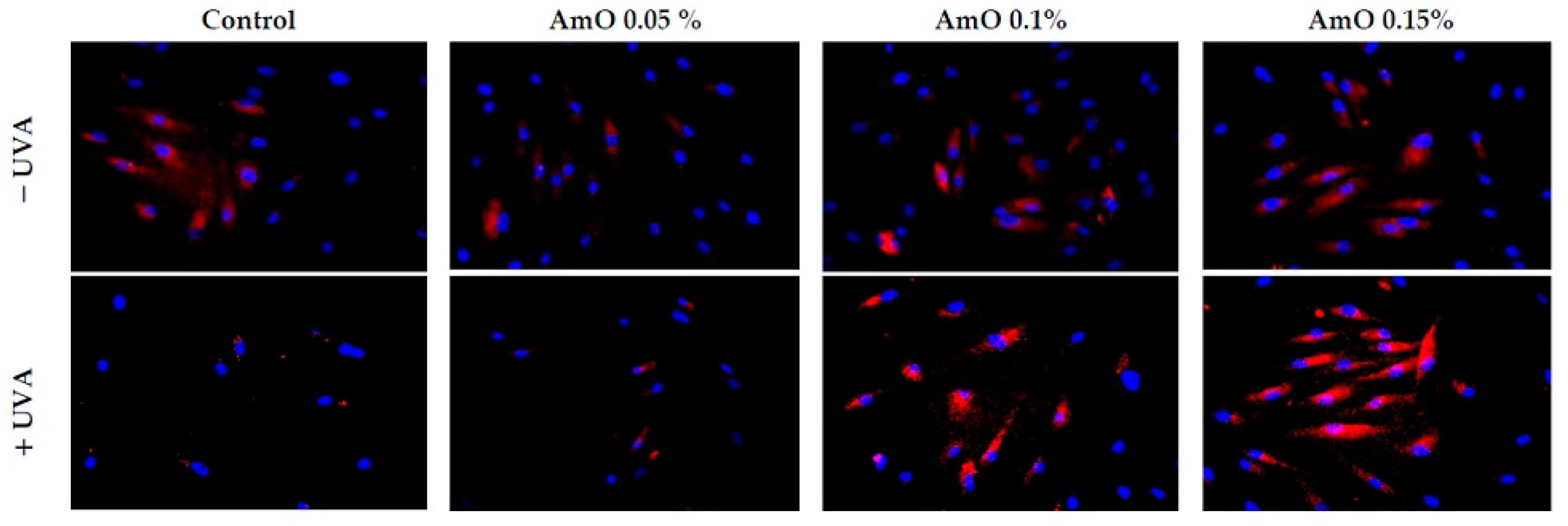
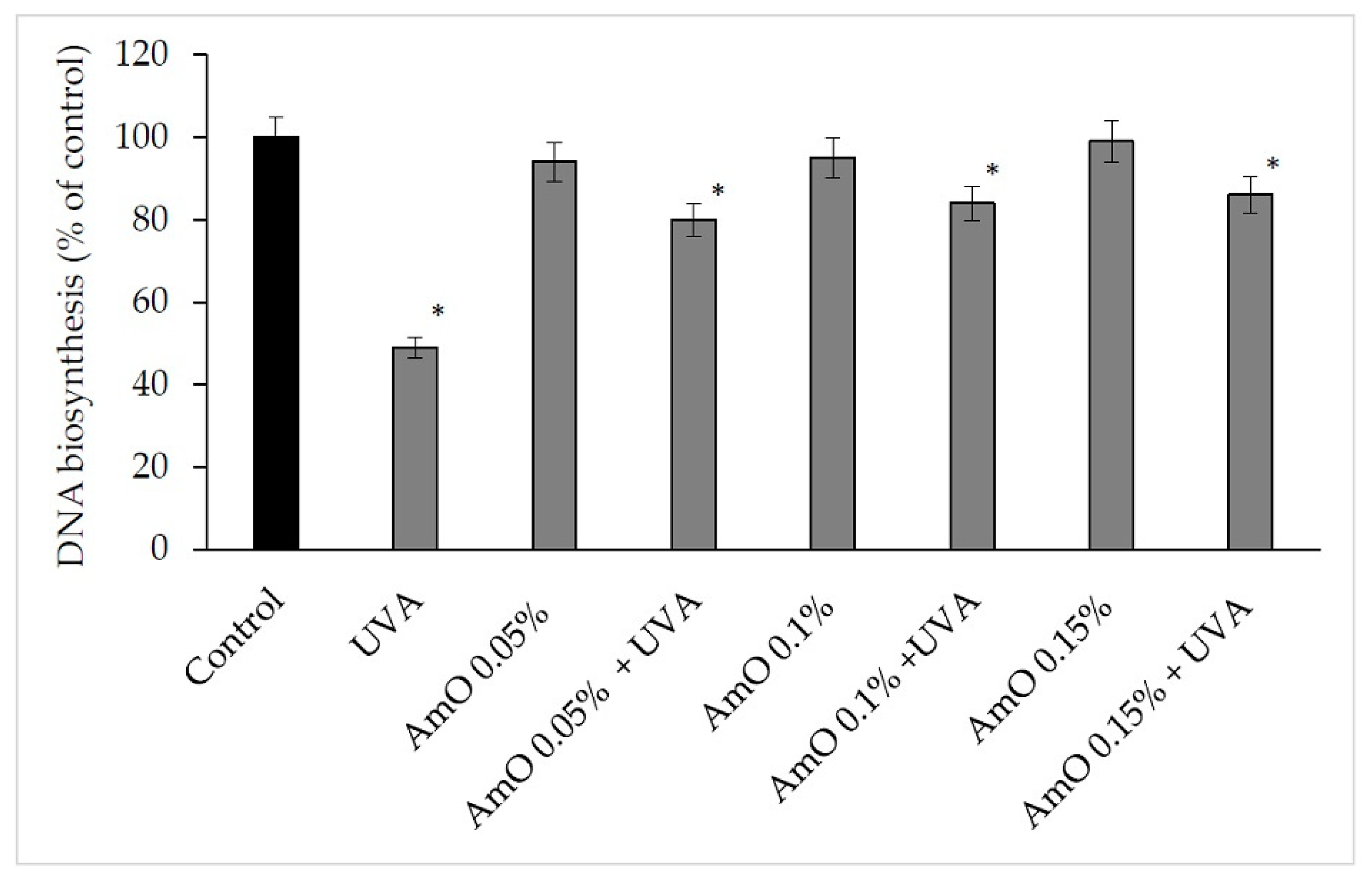
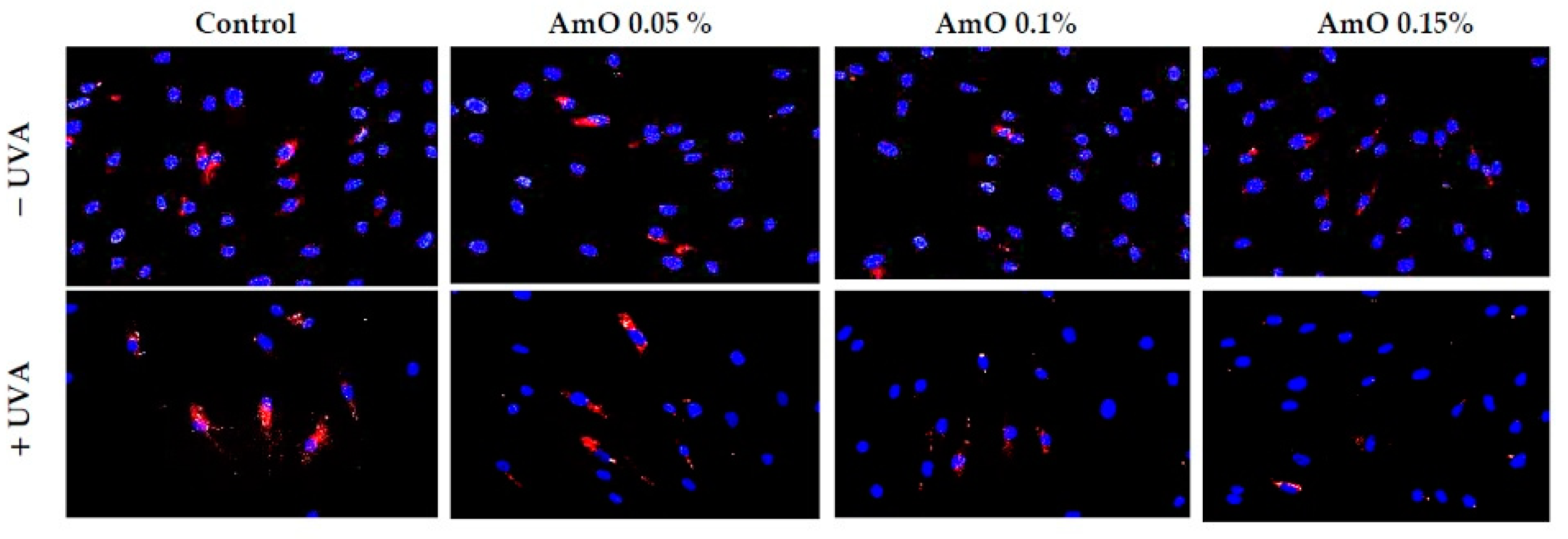
2. Results and Discussion
3. Materials and Methods
3.1. Materials
Plant Materials
3.2. Methods
3.2.1. Cell Lines and Culture
3.2.2. Cell Viability Test MTT
3.2.3. ROS Formation
3.2.4. Cytometric Assay for Apoptosis
3.2.5. Free Thiol Groups Assay
3.2.6. DNA Biosynthesis Assay
3.2.7. Western Immunoblot Analysis
3.2.8. Immunofluorescence Staining and Confocal Microscopy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, Prevention and Therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef]
- Lavker, R.M.; Gerberick, G.F.; Veres, D.; Irwin, C.J.; Kaidbey, K.H. Cumulative Effects from Repeated Exposures to Suberythemal Doses of UVB and UVA in Human Skin. J. Am. Acad. Dermatol. 1995, 32, 53–62. [Google Scholar] [CrossRef]
- Bernerd, F.; Asselineau, D. UVA Exposure of Human Skin Reconstructed in Vitro Induces Apoptosis of Dermal Fibroblasts: Subsequent Connective Tissue Repair and Implications in Photoaging. Cell Death Differ. 1998, 5, 792–802. [Google Scholar] [CrossRef]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New Insights in Photoaging, UVA Induced Damage and Skin Types. Exp. Dermatol. 2014, 23, 7–12. [Google Scholar] [CrossRef]
- Obeng, E. Apoptosis (Programmed Cell Death) and Its Signals—A Review. Braz. J. Biol. 2021, 81, 1133–1143. [Google Scholar] [CrossRef]
- Strozyk, E.; Kulms, D. The Role of AKT/MTOR Pathway in Stress Response to UV-Irradiation: Implication in Skin Carcinogenesis by Regulation of Apoptosis, Autophagy and Senescence. Int. J. Mol. Sci. 2013, 14, 15260–15285. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, H.; Yang, Y.; Zhang, S.; Wang, J.; Zhang, D.; Yu, H. Metformin Attenuates UVA-Induced Skin Photoaging by Suppressing Mitophagy and the PI3K/AKT/MTOR Pathway. Int. J. Mol. Sci. 2022, 23, 6960. [Google Scholar] [CrossRef]
- Gegotek, A.; Jastrzab, A.; Jarocka-Karpowicz, I.; Muszynska, M.; Skrzydlewska, E. The Effect of Sea Buckthorn (Hippophae Rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells. Antioxidants 2018, 7, 110. [Google Scholar] [CrossRef]
- Wolosik, K.; Markowska, A. Amaranthus Cruentus Taxonomy, Botanical Description, and Review of Its Seed Chemical Composition. Nat. Prod. Commun. 2019, 14, 1934578X1984414. [Google Scholar] [CrossRef]
- Ogrodowska, D.; Zadernowski, R.; Czaplicki, S.; Derewiaka, D.; Roszkowska, B. Amaranth Seeds and Products—The Source of Bioactive Compounds. Pol. J. Food Nutr. Sci. 2014, 64, 165–170. [Google Scholar] [CrossRef]
- Gamel, T.; Linssen, J.; Mesallem, A.; Damir, A.; Shekib, L. Effect of Seed Treatments on the Chemical Composition and Properties of Two Amaranth Species: Starch and Protein. J. Sci. Food Agric. 2005, 85, 319–327. [Google Scholar] [CrossRef]
- Berganza, B.; Moran, A.; Rodríguez, G.; Coto, N.; Santamaría, M.; Bressani, R. Effect of Variety and Location on the Total Fat, Fatty Acids and Squalene Content of Amaranth. Plant Foods Hum. Nutr. 2003, 58, 1–6. [Google Scholar] [CrossRef]
- Wolosik, K.; Knas, M.; Zalewska, A.; Niczyporuk, M.; Przystupa, A.W. The Importance and Perspective of Plant-Based Squalene in Cosmetology. J. Cosmet. Sci. 2013, 64, 59–66. [Google Scholar]
- Rittie, L.; Fisher, G.J. Natural and Sun-Induced Aging of Human Skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- Wolosik, K.; Zareba, I.; Surazynski, A.; Markowska, A. The Possible Pre -and Post-UVA Radiation Protective Effect of Amaranth Oil on Human Skin Fibroblast Cells. Pharmacogn. Mag. 2017, 13, 339. [Google Scholar] [CrossRef]
- Semen, K.O.; Den Hartog, G.J.M.; Kaminskyy, D.V.; Sirota, T.V.; Maij NG, A.A.; Yelisyeyeva, O.P.; Bast, A. Redox Modulation by Amaranth Oil in Human Lung Fibroblasts. Nat. Prod. Chem. Res. 2013, 2, 122. [Google Scholar] [CrossRef]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; Roychoudhury, S.; Rathi, B.; Kumar, D. Oxidative Stress in Cancer Cell Metabolism. Antioxidants 2021, 10, 642. [Google Scholar] [CrossRef]
- Manku, M.S.; Horrobin, D.F.; Morse, N.L.; Wright, S.; Burton, J.L. Essential Fatty Acids in the Plasma Phospholipids of Patients with Atopic Eczema. Br. J. Dermatol. 1984, 110, 643–648. [Google Scholar] [CrossRef]
- Emonet, N.; Leccia, M.T.; Favier, A.; Beani, J.C.; Richard, M.J. Thiols and Selenium: Protective Effect on Human Skin Fibroblasts Exposed to UVA Radiation. J. Photochem. Photobiol. B 1997, 40, 84–90. [Google Scholar] [CrossRef]
- Bernerd, F.; Passeron, T.; Castiel, I.; Marionnet, C. The Damaging Effects of Long UVA (UVA1) Rays: A Major Challenge to Preserve Skin Health and Integrity. Int. J. Mol. Sci. 2022, 23, 8243. [Google Scholar] [CrossRef]
- Baba, S.P.; Bhatnagar, A. Role of Thiols in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, F.; Ame, J.; Schreiber, V.; Nakamura, J.; Menissier-de Murcia, J.; De Murcia, G. Poly(ADP-ribose) Polymerase–1 Activation During DNA Damage and Repair. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2006; Volume 409, pp. 493–510. [Google Scholar] [CrossRef]
- Rysava, A.; Vostalova, J.; Rajnochova Svobodova, A. Effect of Ultraviolet Radiation on the Nrf2 Signaling Pathway in Skin Cells. Int. J. Radiat. Biol. 2021, 97, 1383–1403. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a Tetrazolium-Based Semiautomated Colorimetric Assay: Assessment of Chemosensitivity Testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-UVA-Treated Cells | ||||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | Mean Value | % Value | SD | SEM | Me | |
| Control | 0.794 | 0.789 | 0.74 | 0.774 | 100 | 0.029838 | 0.023 | 0.789 |
| AmO 0.05% | 0.74 | 0.685 | 0.542 | 0.655 | 84.711 | 0.102207 | 0.076 | 0.685 |
| AmO 0.1% | 0.78 | 0.76 | 0.709 | 0.749 | 96.856 | 0.036611 | 0.027 | 0.76 |
| AmO 0.15% | 0.804 | 0.731 | 0.721 | 0.752 | 97.157 | 0.04531 | 0.035 | 0.731 |
| UVA-Treated Cells | ||||||||
| UVA (+) | 0.449 | 0.413 | 0.425 | 0.429 | 55.426 * | 0.01833 | 0.013 | 0.425 |
| AmO 0.05% | 0.564 | 0.496 | 0.51 | 0.523 | 67.614 * | 0.035907 | 0.027 | 0.51 |
| AmO 0.1% | 0.461 | 0.636 | 0.625 | 0.574 | 74.160 */** | 0.098015 | 0.075 | 0.625 |
| AmO 0.15% | 0.51 | 0.672 | 0.554 | 0.578 | 74.763 */** | 0.083769 | 0.062 | 0.554 |
| Non-UVA-Treated Cells | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | I | II | III | I | II | III | Mean Value | % Value | |
| Control | 4411 | 4316 | 4363 | 3991 | 3556 | 3639 | 4039 | 4058 | 4049 | 4046.8 | 100 |
| AmO 0.05% | 3951 | 3740 | 3709 | 3513 | 3556 | 3534 | 3977 | 3895 | 3936 | 3756.7 | 92.85 |
| AmO 0.1% | 3808 | 3564 | 3532 | 3862 | 4265 | 3910 | 3688 | 4064 | 3710 | 3822.5 | 94.47 |
| AmO 0.15% | 4000 | 4278 | 4292 | 3816 | 3622 | 3719 | 3732 | 4142 | 4098 | 3966.5 | 98.03 |
| UVA-Treated Cells | |||||||||||
| UVA (+) | 1937 | 1833 | 1944 | 2616 | 2253 | 2326 | 2371 | 2301 | 2336 | 2213 | 54.69 * |
| AmO 0.05% | 2838 | 2828 | 2833 | 2838 | 2826 | 2832 | 2542 | 2525 | 2533 | 2732.7 | 67.54 * |
| AmO 0.1% | 3421 | 3501 | 3461 | 3228 | 3214 | 3221 | 3622 | 3425 | 3462 | 3395 | 83.91 * |
| AmO 0.15% | 3359 | 3342 | 3350 | 3950 | 4038 | 3994 | 3241 | 3175 | 3208 | 3517.4 | 86.93 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolosik, K.; Chalecka, M.; Palka, J.; Surazynski, A. Protective Effect of Amaranthus cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts. Int. J. Mol. Sci. 2023, 24, 10795. https://doi.org/10.3390/ijms241310795
Wolosik K, Chalecka M, Palka J, Surazynski A. Protective Effect of Amaranthus cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts. International Journal of Molecular Sciences. 2023; 24(13):10795. https://doi.org/10.3390/ijms241310795
Chicago/Turabian StyleWolosik, Katarzyna, Magda Chalecka, Jerzy Palka, and Arkadiusz Surazynski. 2023. "Protective Effect of Amaranthus cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts" International Journal of Molecular Sciences 24, no. 13: 10795. https://doi.org/10.3390/ijms241310795
APA StyleWolosik, K., Chalecka, M., Palka, J., & Surazynski, A. (2023). Protective Effect of Amaranthus cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts. International Journal of Molecular Sciences, 24(13), 10795. https://doi.org/10.3390/ijms241310795