
Integrated Transcriptome and Metabolome Dynamic Analysis of Galls Induced by the Gall Mite Aceria pallida on Lycium barbarum Reveals the Molecular Mechanism Underlying Gall Formation and Development



 and
and
Abstract
1. Introduction
2. Results
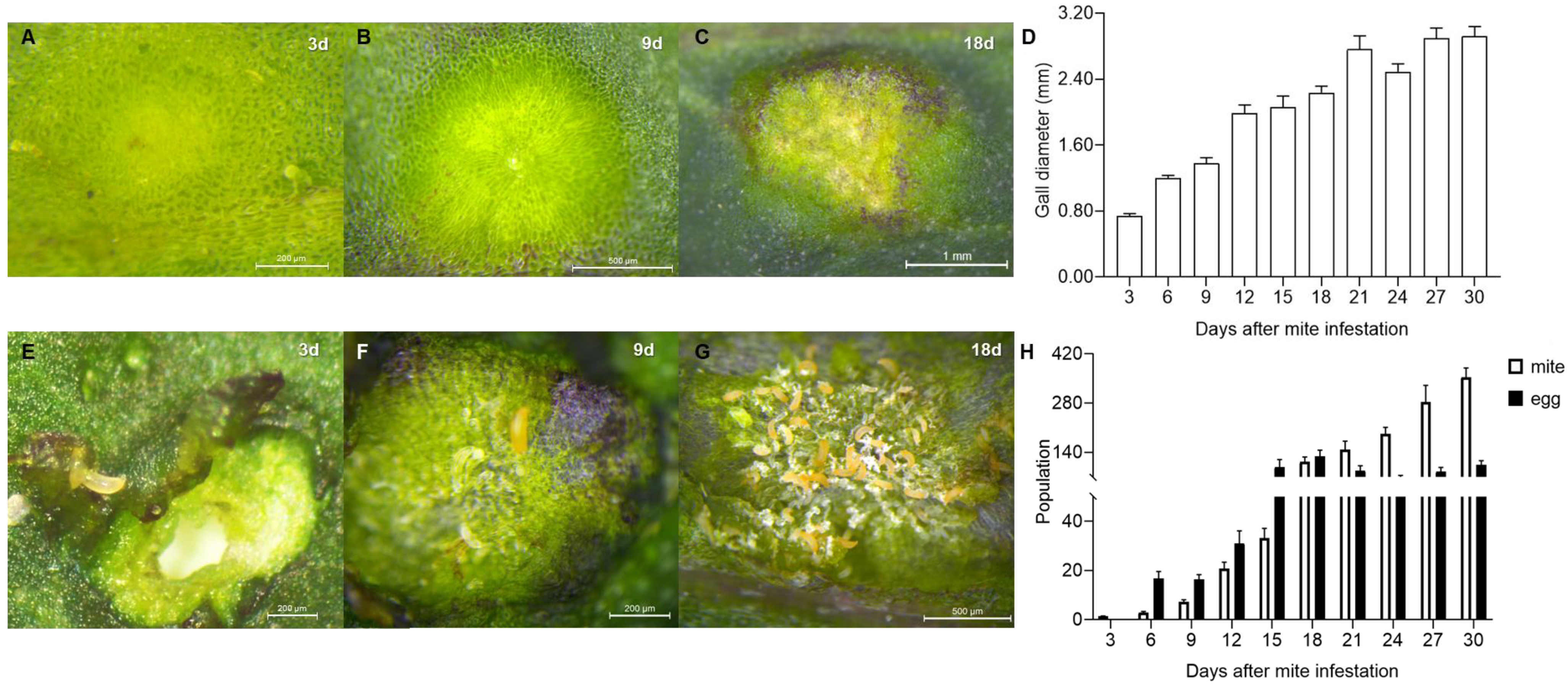
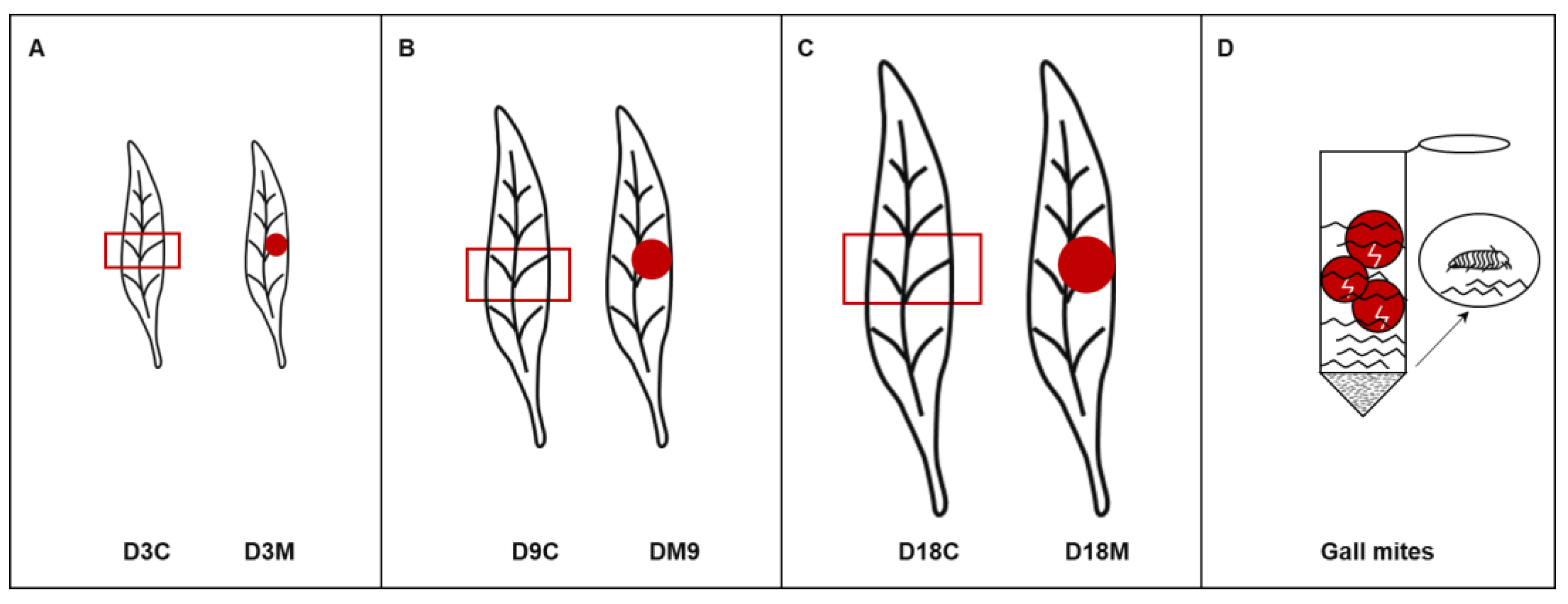
2.1. Alteration to L. barbarum Morphology and Population Dynamics of A. pallida
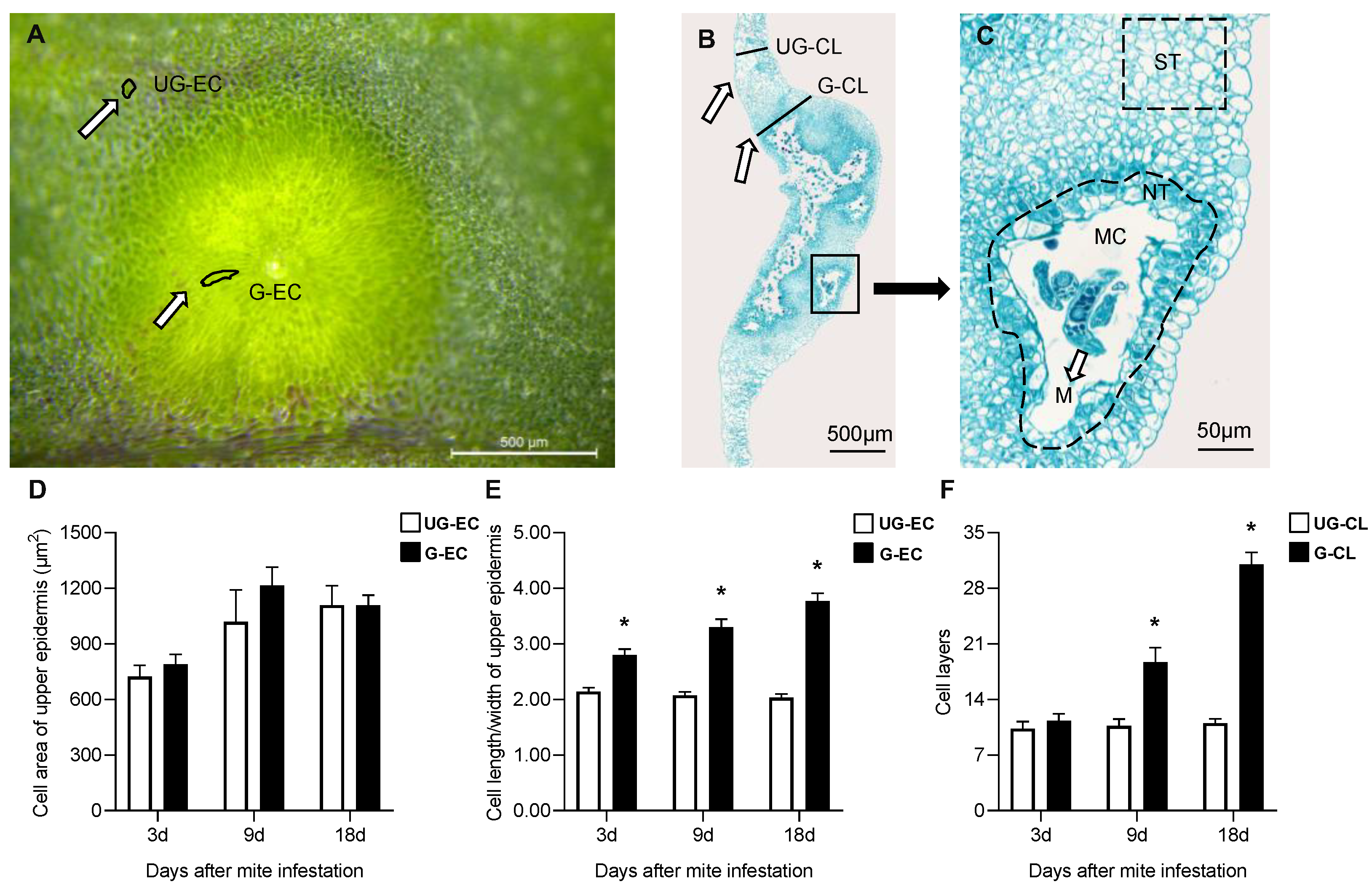
2.2. Histologic Structures of Galls Induced by A. pallida
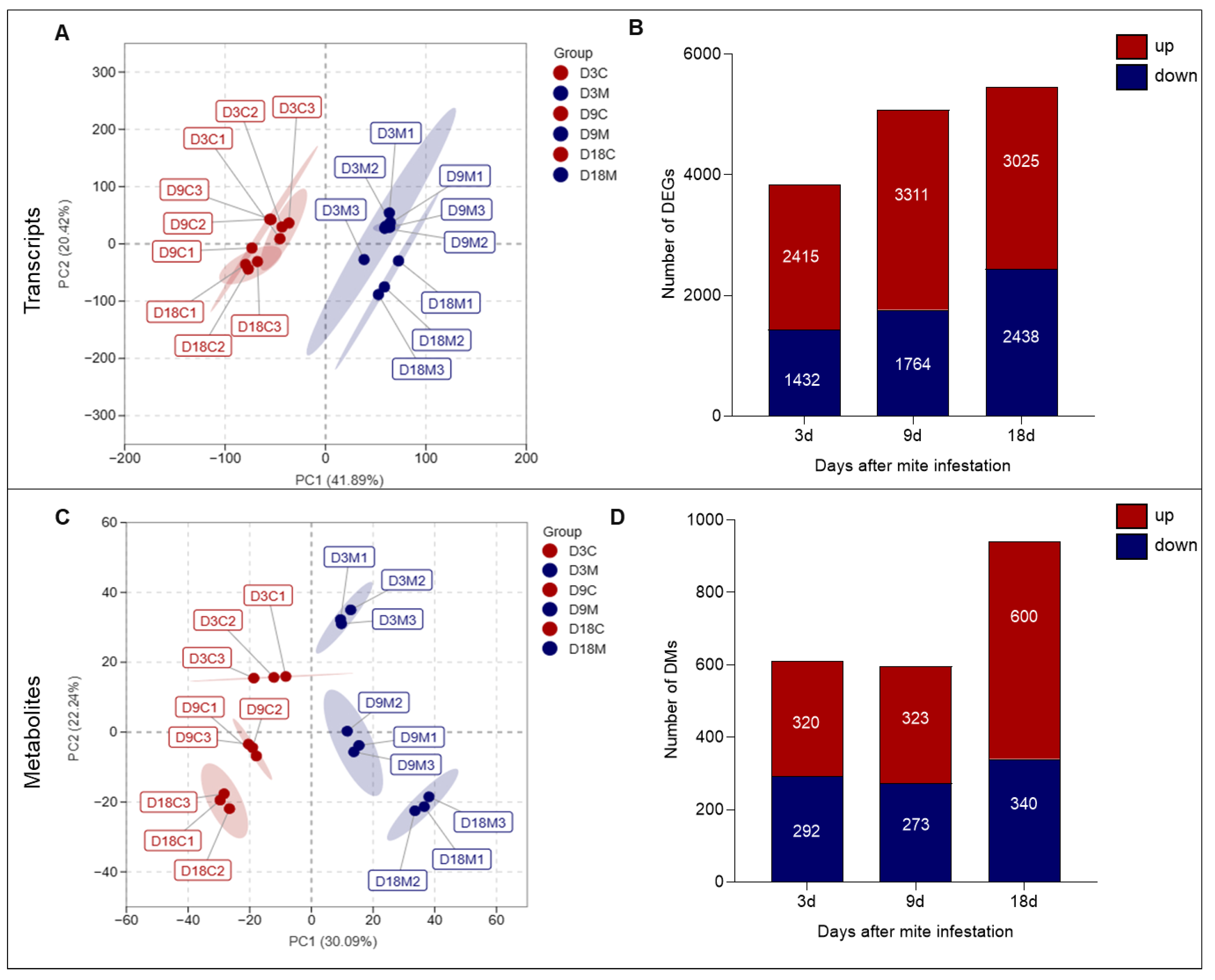
2.3. Alteration of L. barbarum Transcripts and Metabolites by A. pallida
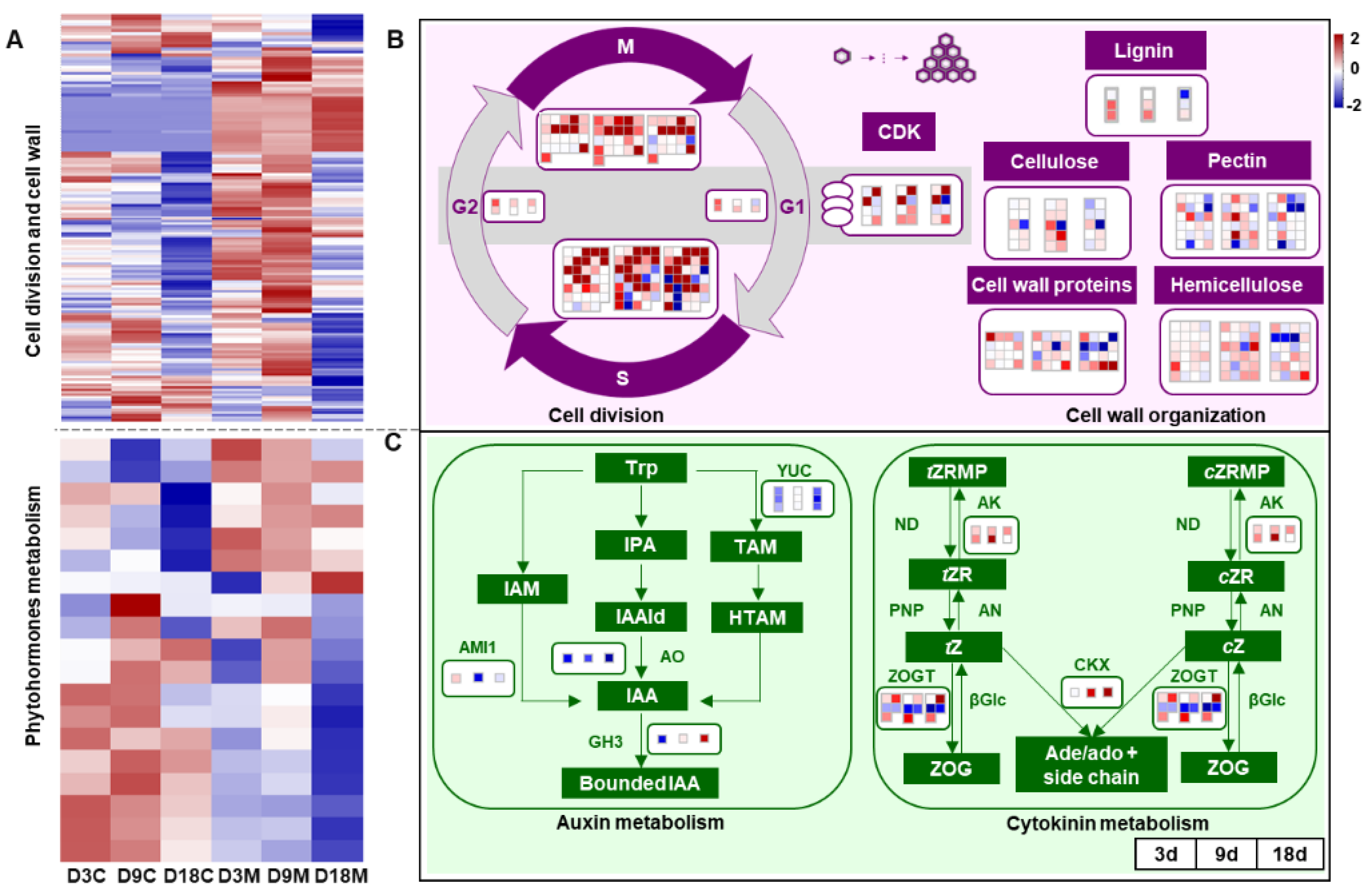
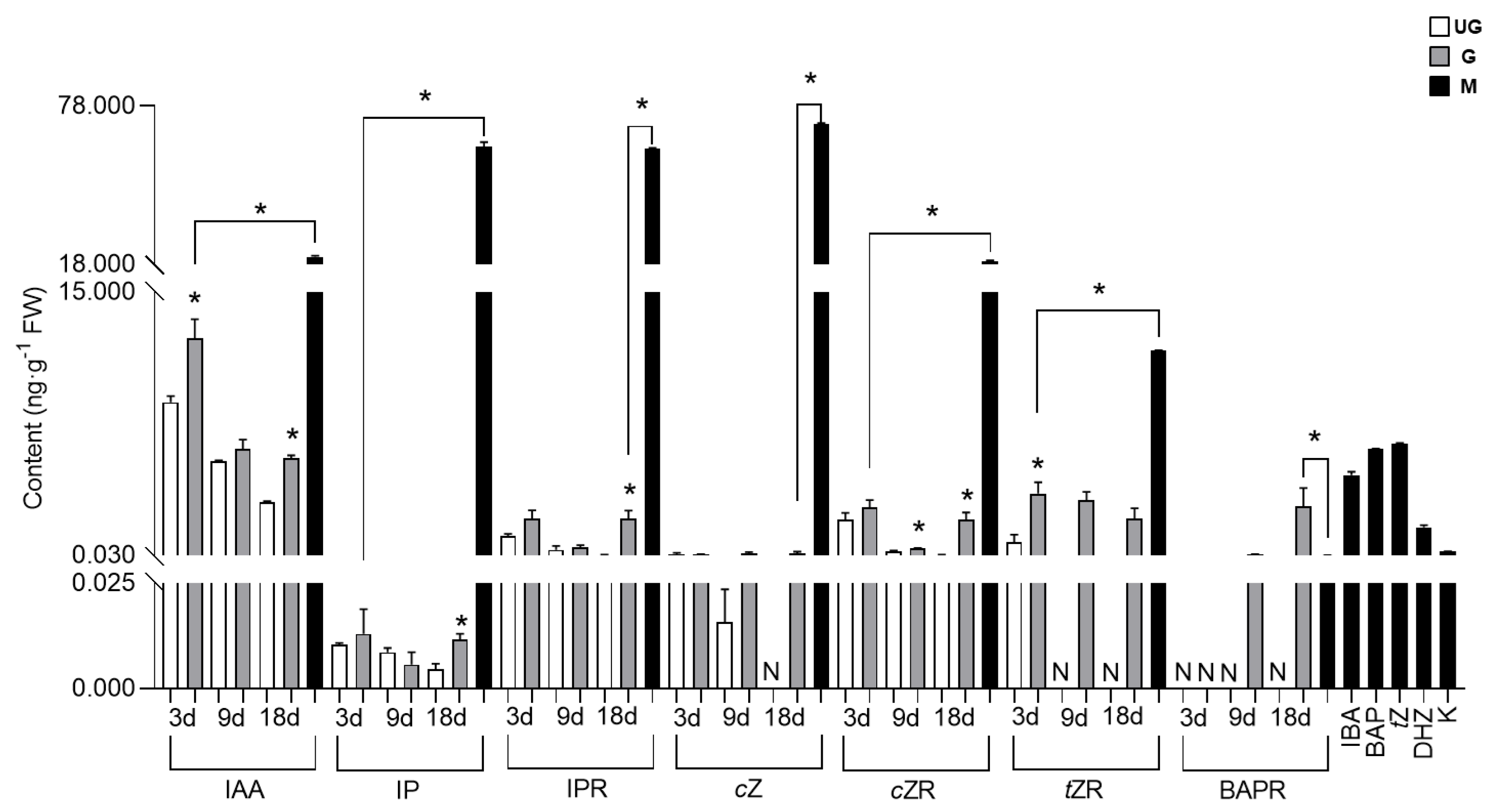
2.4. Manipulation of Hormone Balance and Cell Development in Galls by A. pallida
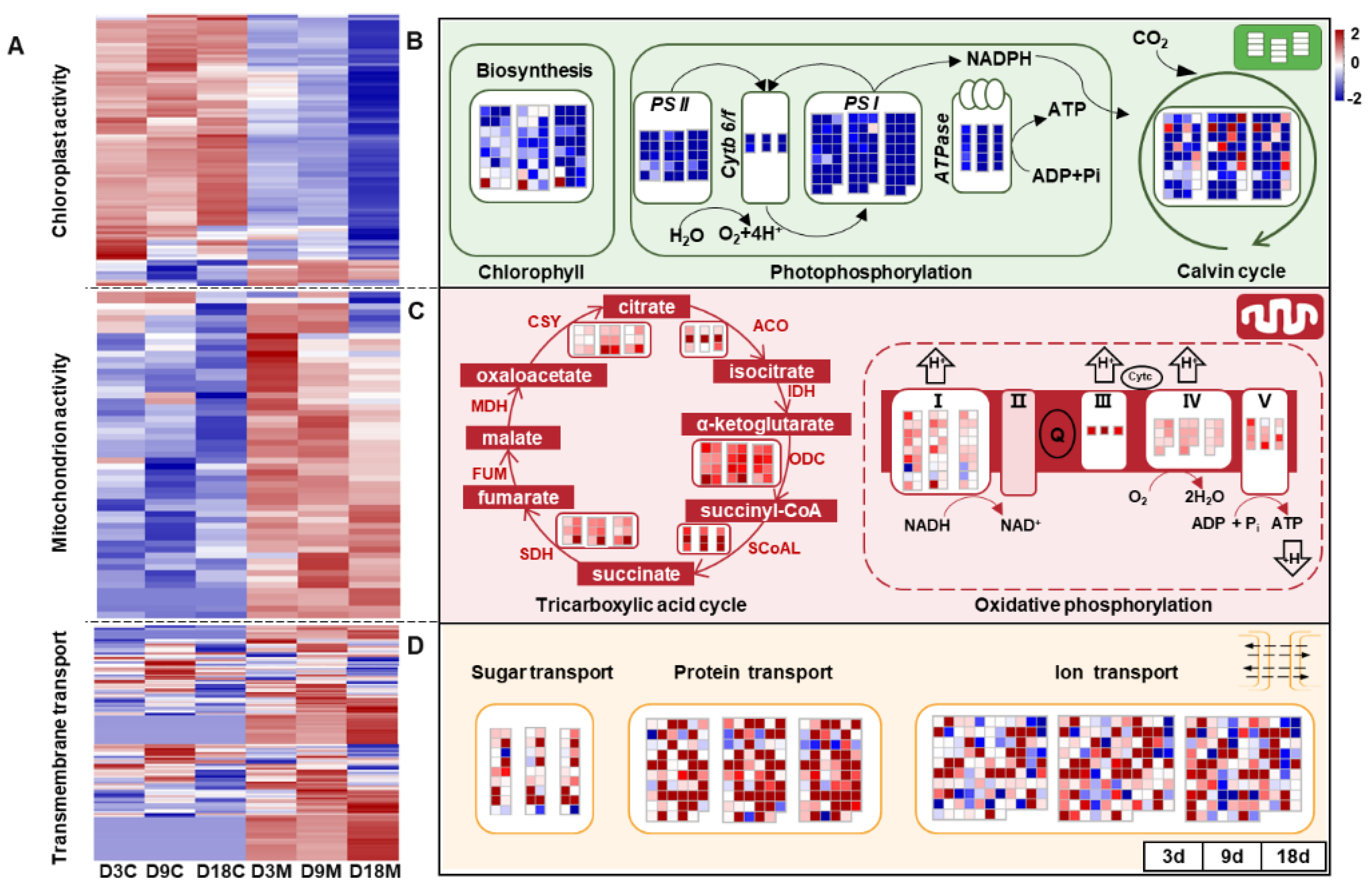
2.5. Modification of Energy Metabolism in Galls by A. pallida
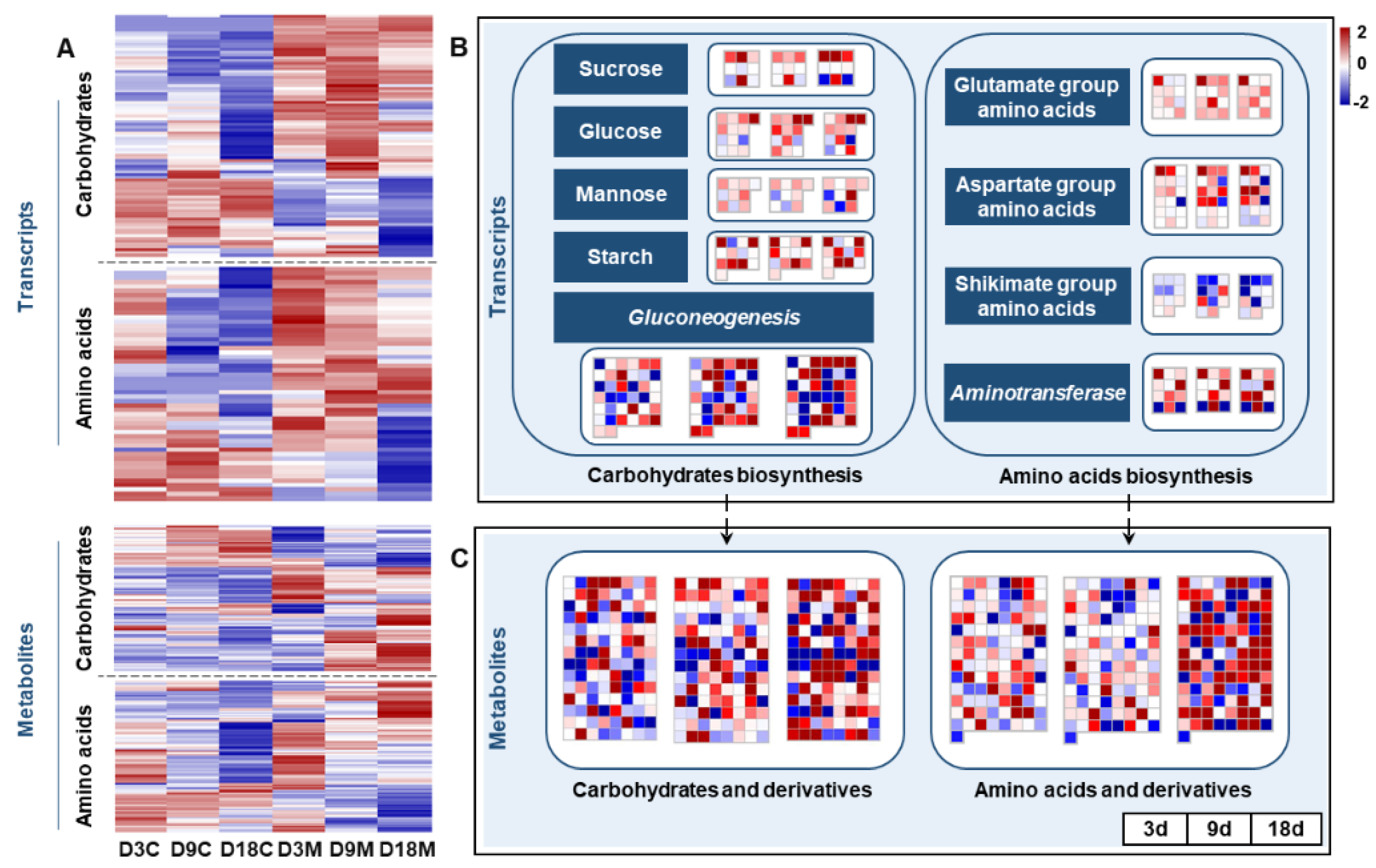
2.6. Changes in Nutrient Metabolism in Galls Induced by A. pallida
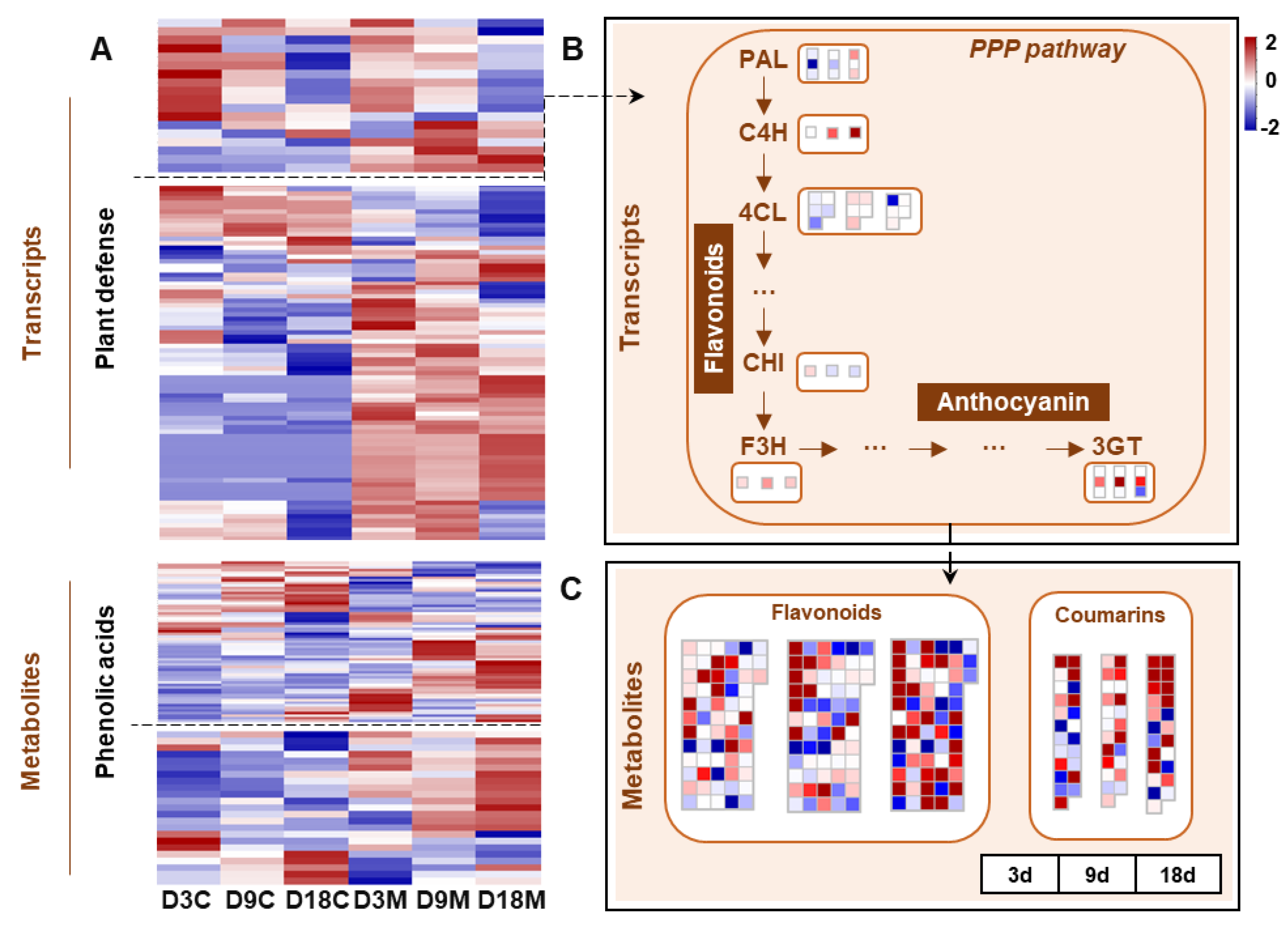
2.7. Defensive Responses of L. barbarum to A. pallida
3. Discussion
4. Materials and Methods
4.1. Plant and Mite Resources and Their Rearing Conditions
4.2. Preparation of Experimental Samples
4.3. Morphological and Anatomical Analysis
4.4. RNA Extraction, Transcriptome Sequencing and Analysis
4.5. Metabolic Analysis by Liquid Chromatography Coupled to Tandem Mass Spectrometry (LC–MS/MS)
4.6. Phytohormone Quantification by LC–MS/MS
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giron, D.; Huguet, E.; Stone, G.N.; Body, M. Insect-induced effects on plants and possible effectors used by galling and leaf-mining insects to manipulate their host-plant. J. Insect Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Mani, M.S. What are galls? In Ecology of Plant Galls; Weisbach, W.W., Vanoye, P., Eds.; Springer: Dordrecht, The Netherlands, 1964; pp. 1–34. [Google Scholar] [CrossRef]
- Gelvin, S.B. Integration of Agrobacterium T-DNA into the plant genome. Annu. Rev. Genet. 2017, 51, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Tooker, J.F.; Helms, A.M. Phytohormone dynamics associated with gall insects, and their potential role in the evolution of the gall-inducing habit. J. Chem. Ecol. 2014, 40, 742–753. [Google Scholar] [CrossRef]
- Zhao, C.; Escalante, L.N.; Chen, H.; Benatti, T.R.; Qu, J.; Chellapilla, S.; Waterhouse, R.M.; Wheeler, D.; Andersson, M.N.; Bao, R.; et al. A massive expansion of effector genes underlies gall-formation in the wheat pest Mayetiola destructor. Curr. Biol. 2015, 25, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Rehill, B.J.; Schultz, J.C. Hormaphis hamamelidis fundatrices benefit by manipulating phenolic metabolism of their host. J. Chem. Ecol. 2012, 38, 496–498. [Google Scholar] [CrossRef]
- Sedlarević, A.; Morina, F.; Toševski, I.; Gašić, U.; Natić, M.; Jović, J.; Krstić, O.; Veljović-Jovanović, S. Comparative analysis of phenolic profiles of ovipositional fluid of Rhinusa pilosa (Mecinini, Curculionidae) and its host plant Linaria vulgaris (Plantaginaceae). Arthropod Plant Interact. 2016, 10, 311–322. [Google Scholar] [CrossRef]
- Gatjens-Boniche, O. The mechanism of plant gall induction by insects: Revealing clues, facts, and consequences in a cross-kingdom complex interaction. Rev. Biol. Trop. 2019, 67, 1359–1382. [Google Scholar] [CrossRef]
- Oates, C.N.; Denby, K.J.; Myburg, A.A.; Slippers, B.; Naidoo, S. Insect gallers and their plant hosts: From omics data to systems biology. Int. J. Mol. Sci. 2016, 17, 1891. [Google Scholar] [CrossRef]
- Takeda, S.; Hirano, T.; Ohshima, I.; Sato, M.H. Recent progress regarding the molecular aspects of insect gall formation. Int. J. Mol. Sci. 2021, 22, 9424. [Google Scholar] [CrossRef]
- Tokuda, M.; Jikumaru, Y.; Matsukura, K.; Takebayashi, Y.; Kumashiro, S.; Matsumura, M.; Kamiya, Y. Phytohormones related to host plant manipulation by a gall-inducing leafhopper. PLoS ONE 2013, 8, e62350. [Google Scholar] [CrossRef]
- Tanaka, Y.; Okada, K.; Asami, T.; Suzuki, Y. Phytohormones in Japanese mugwort gall induction by a gall-inducing gall midge. Biosci. Biotechnol. Biochem. 2013, 77, 1942–1948. [Google Scholar] [CrossRef] [PubMed]
- Tooker, J.F.; De Moraes, C.M. A gall-inducing caterpillar species increases essential fatty acid content of its host plant without concomitant increases in phytohormone levels. Mol. Plant-Microbe Interact. 2009, 22, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, R.G.S.; Castro, A.C.; Isaias, R.M.S. Unique histochemical gradients in a photosynthesis-deficient plant gall. S. Afr. J. Bot. 2014, 92, 97–104. [Google Scholar] [CrossRef]
- Lu, Q.; Chen, X.M.; Yang, Z.X.; Bashir, N.H.; Liu, J.; Cui, Y.Z.; Shao, S.X.; Chen, M.S.; Chen, H. Molecular and histologic adaptation of horned gall induced by the aphid Schlechtendalia chinensis (Pemphigidae). Int. J. Mol. Sci. 2021, 22, 5166. [Google Scholar] [CrossRef] [PubMed]
- Westphal, E.; Bronner, R.; Ret, M.L. Changes in leaves of susceptible and resistant Solanum dulcamara infested by the gall mite Eriophyes cladophthirus (Acarina, Eriophyoidea). Can. J. Bot. 1981, 59, 875–882. [Google Scholar] [CrossRef]
- Martinson, E.O.; Hackett, J.D.; Machado, C.A.; Arnold, A.E. Metatranscriptome analysis of fig flowers provides insights into potential mechanisms for mutualism stability and gall induction. PLoS ONE 2015, 10, e0130745. [Google Scholar] [CrossRef]
- Bragança, G.P.; Oliveira, D.C.; Isaias, R.M.d.S. Compartmentalization of metabolites and enzymatic mediation in nutritive cells of Cecidomyiidae galls on Piper Arboreum Aubl. (Piperaceae). J. Plant Stud. 2016, 6, 11–22. [Google Scholar] [CrossRef]
- Hall, C.R.; Carroll, A.R.; Kitching, R.L. A meta-analysis of the effects of galling insects on host plant secondary metabolites. Arthropod Plant Interact. 2016, 11, 463–473. [Google Scholar] [CrossRef]
- Allison, S.D.; Schultz, J.C. Biochemical responses of chestnut oak to a galling cynipid. J. Chem. Ecol. 2005, 31, 151–166. [Google Scholar] [CrossRef]
- Dorleku, W.P.; Gbewonyo, W.S.K. Levels of polyphenol oxidase activity in leaves of Milicia seedlings is indicative of the resistance to Phytolyma lata-induced gall disease. Agric. For. Entomol. 2021, 23, 518–526. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, M.S.; Liu, X. Changes in phytohormones and fatty acids in wheat and rice seedlings in response to Hessian fly (Diptera: Cecidomyiidae) infestation. J. Econ. Entomol. 2011, 104, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Rauf, A.; Khan, H. The relevance of folkloric usage of plant galls as medicines: Finding the scientific rationale. Biomed. Pharmacother. 2018, 97, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Favery, B.; Dubreuil, G.; Chen, M.S.; Giron, D.; Abad, P. Gall-inducing parasites: Convergent and conserved strategies of plant manipulation by insects and nematodes. Annu. Rev. Phytopathol. 2020, 58, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Martinson, E.O.; Werren, J.H.; Egan, S.P. Tissue-specific gene expression shows a cynipid wasp repurposes oak host gene networks to create a complex and novel parasite-specific organ. Mol. Ecol. 2022, 31, 3228–3240. [Google Scholar] [CrossRef] [PubMed]
- Bentur, J.S.; Rawat, N.; Divya, D.; Sinha, D.K.; Agarrwal, R.; Atray, I.; Nair, S. Rice-gall midge interactions: Battle for survival. J. Insect Physiol. 2016, 84, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Thomas, D.d.B.; Body, M.; Glevarec, G.; Reichelt, M.; Unsicker, S.; Bruneau, M.; Renou, J.P.; Huguet, E.; Dubreuil, G.; et al. Leaf-mining by Phyllonorycter blancardella reprograms the host-leaf transcriptome to modulate phytohormones associated with nutrient mobilization and plant defense. J. Insect Physiol. 2016, 84, 114–127. [Google Scholar] [CrossRef]
- Takei, M.; Yoshida, S.; Kawai, T.; Hasegawa, M.; Suzuki, Y. Adaptive significance of gall formation for a gall-inducing aphids on Japanese elm trees. J. Insect Physiol. 2015, 72, 43–51. [Google Scholar] [CrossRef]
- Jankiewicz, L.S.; Guzicka, M.; Marasek, C.A. Anatomy and ultrastructure of galls induced by Neuroterus quercusbaccarum (Hymenoptera: Cynipidae) on oak leaves (Quercus robur). Insects 2021, 12, 850. [Google Scholar] [CrossRef]
- Martini, V.C.; Moreira, A.S.F.P.; Kuster, V.C.; Oliveira, D.C. Galling insects as phenotype manipulators of cell wall composition during the development of galls induced on leaves of Aspidosperma tomentosum (Apocynaceae). S. Afr. J. Bot. 2019, 127, 226–233. [Google Scholar] [CrossRef]
- Qi, Y.; Duan, C.; Ren, L.; Wu, H. Growth dynamics of galls and chemical defence response of Pinus thunbergii Parl. to the pine needle gall midge, Thecodiplosis japonensis Uchida & Inouye (Diptera: Cecidomyiidae). Sci. Rep. 2020, 10, 12289–12300. [Google Scholar] [CrossRef]
- Liu, P.; Yang, Z.X.; Chen, X.M.; Foottit, R.G. The effect of the gall-forming aphid Schlechtendalia chinensis (Hemiptera: Aphididae) on leaf wing ontogenesis in Rhus chinensis (Sapindales: Anacardiaceae). Ann. Entomol. Soc. Am. 2014, 107, 242–250. [Google Scholar] [CrossRef]
- Nabity, P.D.; Haus, M.J.; Berenbaum, M.R.; DeLucia, E.H. Leaf-galling phylloxera on grapes reprograms host metabolism and morphology. Proc. Natl. Acad. Sci. USA 2013, 110, 16663–16668. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Yoza, M.; Amano, T.; Ohshima, I.; Hirano, T.; Sato, M.H.; Sakamoto, T.; Kimura, S. Comparative transcriptome analysis of galls from four different host plants suggests the molecular mechanism of gall development. PLoS ONE 2019, 14, e0223686. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Vishyakov, A.E.; Dodueva, I.T.; Osipova, M.A.; Sukhareva, S.I.; Shavarda, A.L. Gallogenesis induced by eryophyoid mites (Acariformes: Eriophyoidea). Entomol. Rev. 2016, 95, 1137–1143. [Google Scholar] [CrossRef]
- Nuzzaci, G.; Lillo, E.d. Perspectives on eriophyoid mite research. Entomologica 1996, 30, 73–91. [Google Scholar] [CrossRef]
- Petanovic, R.; Kielkiewicz, M. Plant-eriophyoid mite interactions: Cellular biochemistry and metabolic responses induced in mite-injured plants. Part I. Exp. Appl. Acarol. 2010, 51, 61–80. [Google Scholar] [CrossRef]
- Petanovic, R.; Kielkiewicz, M. Plant-eriophyoid mite interactions: Specific and unspecific morphological alterations. Part II. Exp. Appl. Acarol. 2010, 51, 81–91. [Google Scholar] [CrossRef]
- Veldtman, R.; McGeoch, M.A. Gall-forming insect species richness along a non-scleromorphic vegetation rainfall gradient in South Africa: The importance of plant community composition. Austral Ecol. 2003, 28, 1–13. [Google Scholar] [CrossRef]
- Body, M.J.A.; Zinkgraf, M.S.; Whitham, T.G.; Lin, C.H.; Richardson, R.A.; Appel, H.M.; Schultz, J.C. Heritable phytohormone profiles of poplar genotypes vary in resistance to a galling aphid. Mol. Plant Microbe Interact. 2019, 32, 654–672. [Google Scholar] [CrossRef]
- Huang, M.Y.; Huang, W.D.; Chou, H.M.; Chen, C.C.; Chen, P.J.; Chang, Y.T.; Yang, C.M. Structural, biochemical, and physiological characterization of photosynthesis in leaf-derived cup-shaped galls on Litsea acuminata. BMC Plant Biol. 2015, 15, 61–72. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Tanaka, H.; Hasegawa, M.; Tokuda, M.; Asami, T.; Suzuki, Y. Phytohormones and willow gall induction by a gall-inducing sawfly. New Phytol. 2012, 196, 586–595. [Google Scholar] [CrossRef]
- Liu, S.; Yang, M.K.; Li, J.L.; Guo, K.; Qiao, H.L.; Xu, R.; Xu, C.Q.; Chen, J. Overwintering status and species identification of gall mite in major wolfberry production areas. Chin. J. Chin. Mater. Med. 2019, 44, 2208–2212. [Google Scholar] [CrossRef]
- Li, J.L.; Liu, S.; Guo, K.; Zhang, F.; Qiao, H.L.; Chen, J.M.; Yang, M.K.; Zhu, X.; Xu, R.; Xu, C.Q.; et al. Plant-mediated competition facilitates a phoretic association between a gall mite and a psyllid vector. Exp. Appl. Acarol. 2018, 76, 325–337. [Google Scholar] [CrossRef]
- Nasareen, P.N.M.; Vardhanan, Y.S.; Ramani, N. Damage assessment of the gall mite Aceria pongamiae Keifer 1966 (Acari: Eriophyidae) on Pongamia pinnata (L.) Pierre. In Prospects in Bioscience: Addressing the Issues; Sabu, A., Augustine, A., Eds.; Springer: New Delhi, India, 2012; pp. 325–333. [Google Scholar] [CrossRef]
- Carneiro, R.G.S.; Isaias, R.M.S.; Moreira, A.; Oliveira, D.C. Reacquisition of new meristematic sites determines the development of a new organ, the Cecidomyiidae gall on Copaifera langsdorffii Desf. (Fabaceae). Front. Plant Sci. 2017, 8, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, C. The Arabidopsis cell division cycle. Arab. Book 2009, 7, e0120. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed]
- Frebort, I.; Kowalska, M.; Hluska, T.; Frebortova, J.; Galuszka, P. Evolution of cytokinin biosynthesis and degradation. J. Exp. Bot. 2011, 62, 2431–2452. [Google Scholar] [CrossRef]
- Siqueira, J.A.; Hardoim, P.; Ferreira, P.C.G.; Nunes-Nesi, A.; Hemerly, A.S. Unraveling interfaces between energy metabolism and cell cycle in plants. Trends Plant Sci. 2018, 23, 731–747. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.M.; Hosseini, P.; Alkharouf, N.W.; Hussein, E.H.; Gamal El-Din Ael, K.; Aly, M.A.; Matthews, B.F. Analysis of gene expression in soybean (Glycine max) roots in response to the root knot nematode Meloidogyne incognita using microarrays and KEGG pathways. BMC Genom. 2011, 12, 220. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Moreira, A.; Isaias, R.M.S.; Martini, V.; Rezende, U.C. Sink status and photosynthetic rate of the leaflet galls induced by Bystracoccus mataybae (Eriococcidae) on Matayba guianensis (Sapindaceae). Front. Plant Sci. 2017, 8, 1249–1260. [Google Scholar] [CrossRef]
- Hirano, T.; Kimura, S.; Sakamoto, T.; Okamoto, A.; Nakayama, T.; Matsuura, T.; Ikeda, Y.; Takeda, S.; Suzuki, Y.; Ohshima, I.; et al. Reprogramming of the developmental program of Rhus javanica during initial stage of gall induction by Schlechtendalia chinensis. Front. Plant Sci. 2020, 11, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, L.; Connor, E.F. Exogenous phytohormones and the induction of plant galls by insects. Arthropod Plant Interact. 2014, 8, 339–348. [Google Scholar] [CrossRef]
- Mapes, C.C.; Davies, P.J. Indole-3-acetic acid and ball gall development on Solidago altissima. New Phytol. 2001, 151, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Mapes, C.C.; Davies, P.J. Cytokinins in the ball gall of Solidago altissima and in the gall forming larvae of Eurosta solidaginis. New Phytol. 2001, 151, 203–212. [Google Scholar] [CrossRef]
- Suzuki, H.; Yokokura, J.; Ito, T.; Arai, R.; Yokoyama, C.; Toshima, H.; Nagata, S.; Asami, T.; Suzuki, Y. Biosynthetic pathway of the phytohormone auxin in insects and screening of its inhibitors. Insect Biochem. Mol. Biol. 2014, 53, 66–72. [Google Scholar] [CrossRef]
- Andreas, P.; Kisiala, A.; Emery, R.J.N.; De Clerck-Floate, R.; Tooker, J.F.; Price, P.W.; Miller Iii, D.G.; Chen, M.S.; Connor, E.F. Cytokinins are abundant and widespread among insect species. Plants 2020, 9, 208. [Google Scholar] [CrossRef]
- Ponce, G.E.; Fuse, M.; Chan, A.; Connor, E.F. The localization of phytohormones within the gall-inducing insect Eurosta solidaginis (Diptera: Tephritidae). Arthropod Plant Interact. 2021, 15, 375–385. [Google Scholar] [CrossRef]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.d.P.; Abrahão, J.; et al. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Dsouza, M.R.; Ravishankar, B.E. Nutritional sink formation in galls of Ficus glomerata Roxb. (Moraceae) by the insect Pauropsylla depressa (Psyllidae, Hemiptera). Trop. Ecol. 2014, 55, 129–136. [Google Scholar]
- Ikai, N.; Hijii, N. Manipulation of tannins in oaks by galling cynipids. J. For. Res. 2017, 12, 316–319. [Google Scholar] [CrossRef]
- Wasson, A.P.; Pellerone, F.I.; Mathesius, U. Silencing the flavonoid pathway in Medicago truncatula inhibits root nodule formation and prevents auxin transport regulation by rhizobia. Plant Cell 2006, 18, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Imbert, M.P.; Wilson, L.A. Stimulatory and inhibitory effects of scopoletin on IAA oxidase preparations from sweet potato. Phytochemistry 1970, 9, 1787–1794. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323–338. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Schwacke, R.; Ponce-Soto, G.Y.; Krause, K.; Bolger, A.M.; Arsova, B.; Hallab, A.; Gruden, K.; Stitt, M.; Bolger, M.E.; Usadel, B. MapMan4: A refined protein classification and annotation framework applicable to multi-omics data analysis. Mol. Plant 2019, 12, 879–892. [Google Scholar] [CrossRef]
- Niu, Q.F.; Zong, Y.; Qian, M.J.; Yang, F.X.; Teng, Y.W. Simultaneous quantitative determination of major plant hormones in pear flowers and fruit by UPLC/ESI-MS/MS. Anal. Methods 2014, 6, 1766–1773. [Google Scholar] [CrossRef]
- Simura, J.; Antoniadi, I.; Siroka, J.; Tarkowska, D.; Strnad, M.; Ljung, K.; Novak, O. Plant hormonomics: Multiple phytohormone profiling by targeted metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Total DEGs | Upregulated | Downregulated | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 3 d | 9 d | 18 d | 3 d | 9 d | 18 d | 3 d | 9 d | 18 d | |
| Photosynthesis | 152 | 159 | 185 | 7 | 12 | 13 | 145 | 147 | 172 |
| Cellular respiration | 69 | 70 | 65 | 68 | 68 | 61 | 1 | 2 | 4 |
| Vesicle trafficking | 35 | 63 | 61 | 35 | 63 | 58 | 0 | 0 | 3 |
| Protein translocation | 31 | 46 | 39 | 29 | 38 | 25 | 2 | 8 | 14 |
| Phytohormone action | 23 | 44 | 53 | 18 | 25 | 28 | 5 | 19 | 25 |
| Cell division | 50 | 72 | 77 | 46 | 62 | 57 | 4 | 10 | 20 |
| Cytoskeleton organization | 25 | 35 | 26 | 24 | 33 | 21 | 1 | 2 | 5 |
| Cell wall organization | 14 | 25 | 26 | 9 | 16 | 9 | 5 | 9 | 17 |
| Carbohydrate metabolism | 28 | 45 | 45 | 21 | 36 | 32 | 7 | 9 | 13 |
| Amino acid metabolism | 24 | 41 | 44 | 14 | 27 | 25 | 10 | 14 | 19 |
| Protein biosynthesis | 184 | 277 | 199 | 180 | 222 | 139 | 4 | 55 | 60 |
| Lipid metabolism | 54 | 77 | 95 | 39 | 53 | 60 | 15 | 24 | 35 |
| Secondary metabolism | 19 | 21 | 29 | 11 | 13 | 16 | 8 | 8 | 13 |
| Type | Total DMs | Upregulated | Downregulated | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 3 d | 9 d | 18 d | 3 d | 9 d | 18 d | 3 d | 9 d | 18 d | |
| Carbohydrates | 63 | 58 | 83 | 32 | 36 | 56 | 31 | 22 | 27 |
| Amino acids | 52 | 41 | 86 | 30 | 19 | 60 | 22 | 22 | 26 |
| Fatty acids | 36 | 42 | 53 | 25 | 27 | 34 | 11 | 15 | 19 |
| Lipids | 78 | 87 | 111 | 47 | 69 | 82 | 31 | 18 | 29 |
| Alkaloids | 117 | 101 | 169 | 46 | 43 | 105 | 71 | 58 | 64 |
| Organic acids | 47 | 54 | 76 | 26 | 31 | 54 | 21 | 23 | 22 |
| Terpenes | 58 | 52 | 77 | 29 | 18 | 38 | 29 | 34 | 39 |
| Phenolics | 60 | 61 | 92 | 31 | 31 | 56 | 29 | 30 | 36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Li, H.; Qiao, H.; Guo, K.; Xu, R.; Wei, H.; Wei, J.; Liu, S.; Xu, C. Integrated Transcriptome and Metabolome Dynamic Analysis of Galls Induced by the Gall Mite Aceria pallida on Lycium barbarum Reveals the Molecular Mechanism Underlying Gall Formation and Development. Int. J. Mol. Sci. 2023, 24, 9839. https://doi.org/10.3390/ijms24129839
Yang M, Li H, Qiao H, Guo K, Xu R, Wei H, Wei J, Liu S, Xu C. Integrated Transcriptome and Metabolome Dynamic Analysis of Galls Induced by the Gall Mite Aceria pallida on Lycium barbarum Reveals the Molecular Mechanism Underlying Gall Formation and Development. International Journal of Molecular Sciences. 2023; 24(12):9839. https://doi.org/10.3390/ijms24129839
Chicago/Turabian StyleYang, Mengke, Huanle Li, Haili Qiao, Kun Guo, Rong Xu, Hongshuang Wei, Jianhe Wei, Sai Liu, and Changqing Xu. 2023. "Integrated Transcriptome and Metabolome Dynamic Analysis of Galls Induced by the Gall Mite Aceria pallida on Lycium barbarum Reveals the Molecular Mechanism Underlying Gall Formation and Development" International Journal of Molecular Sciences 24, no. 12: 9839. https://doi.org/10.3390/ijms24129839
APA StyleYang, M., Li, H., Qiao, H., Guo, K., Xu, R., Wei, H., Wei, J., Liu, S., & Xu, C. (2023). Integrated Transcriptome and Metabolome Dynamic Analysis of Galls Induced by the Gall Mite Aceria pallida on Lycium barbarum Reveals the Molecular Mechanism Underlying Gall Formation and Development. International Journal of Molecular Sciences, 24(12), 9839. https://doi.org/10.3390/ijms24129839