
Effects of Heavy Metals on Stomata in Plants: A Review


Abstract
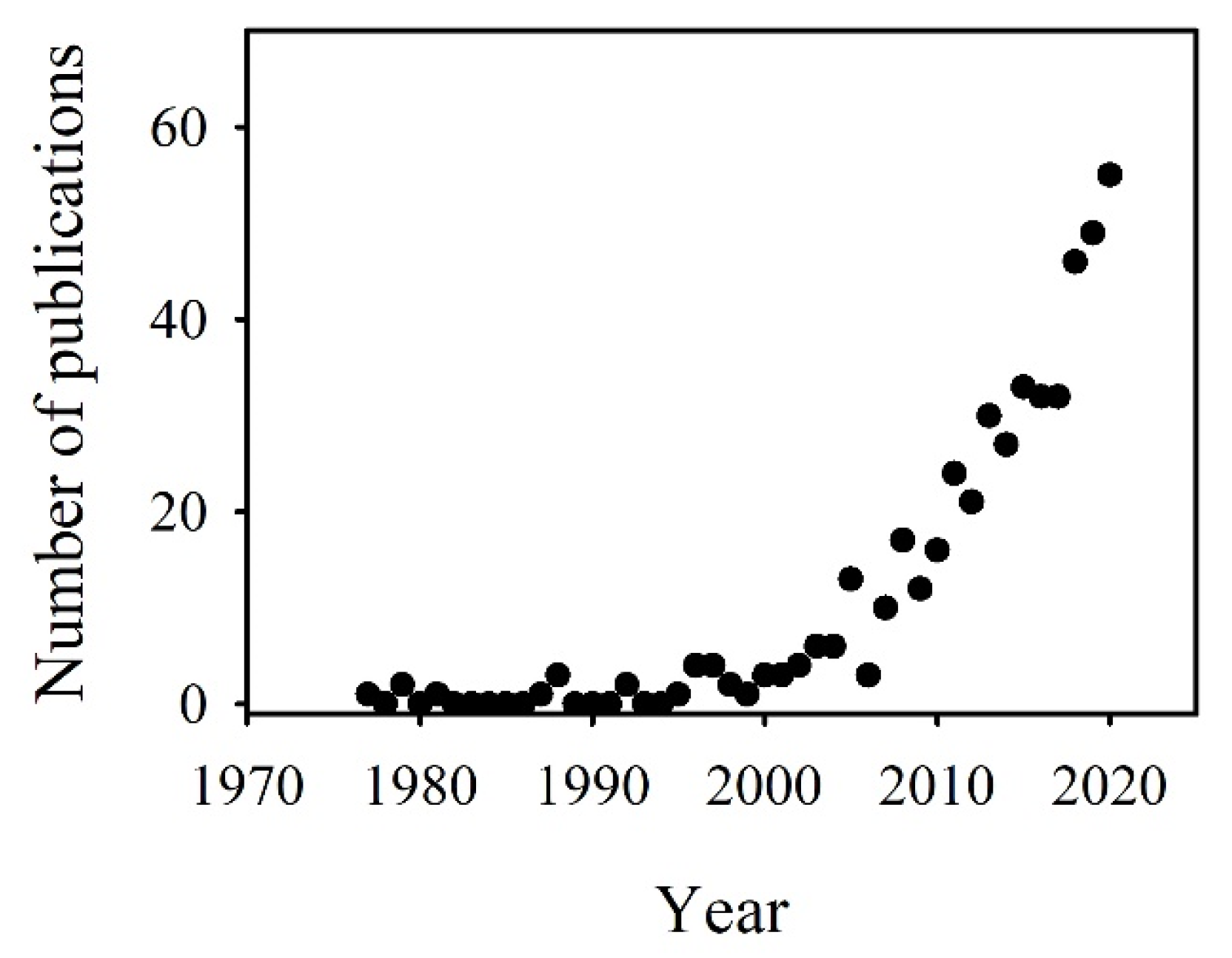
1. Introduction
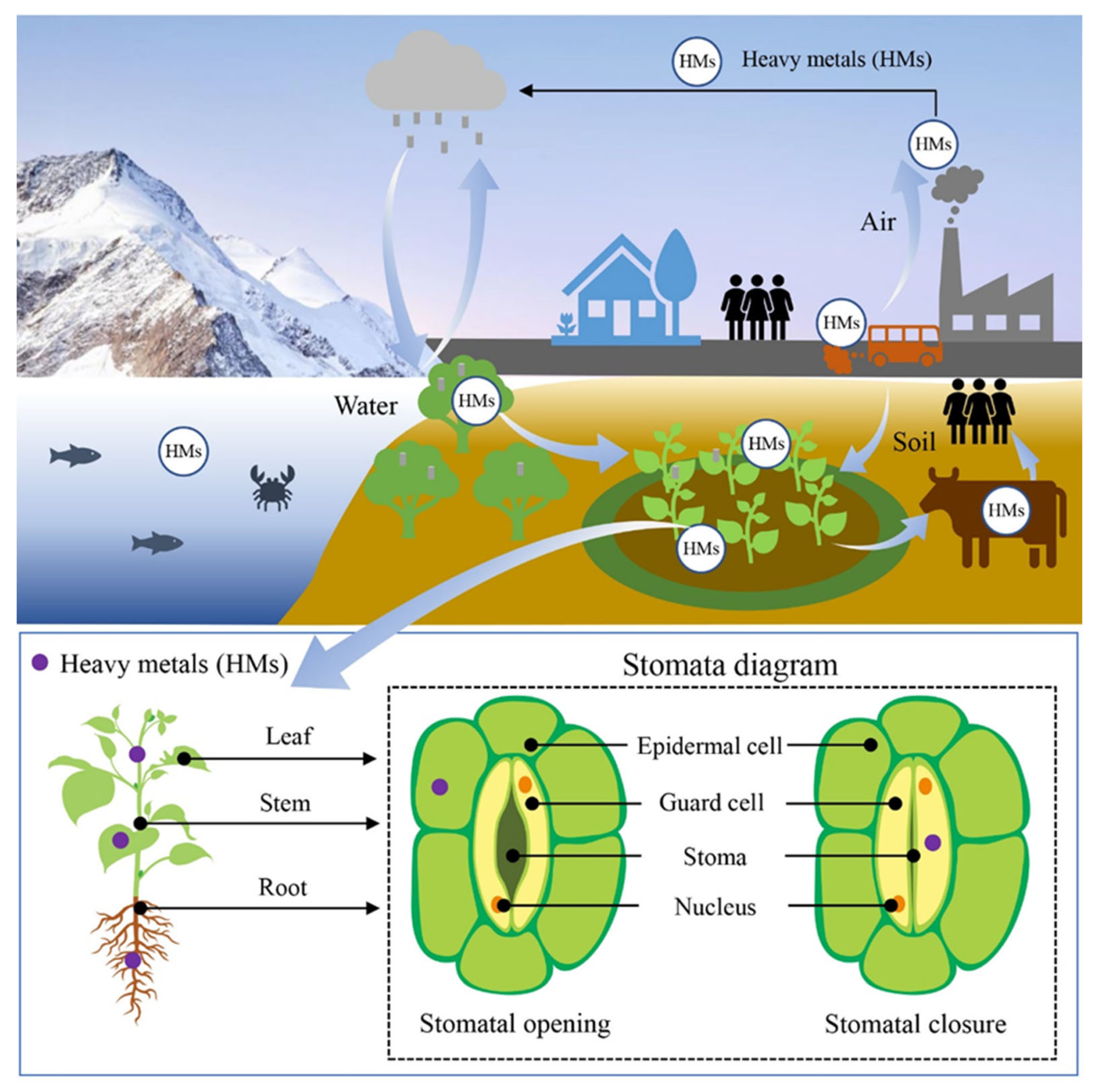
2. Structure and Function of Stomata
3. Role of Stomata in Heavy-Metal-Induced Plant Damage
4. Toxic Effects and Mechanisms of Heavy Metals on Plant Stomata
4.1. Effect of Heavy Metals on Stomatal Behavior

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heavy Metal | Processing Time and Concentration | Plant Species | Plant Type | Toxic Effect | References |
|---|---|---|---|---|---|
| Cd | 50, 100 mg/kg; 3 months | Pennisetum sp. | C3 | Stomatal closure | [27] |
| 0.6 Nm/L; 7 days | Brassica juncea | C3 | Stomatal closure | [30] | |
| 15 mg/L; 10 days | Monochoria hastata | C3 | Stomatal closure | [37] | |
| 24 mg/kg; 40 days | Brassica campestris ssp. Brassica juncea Czernajew | C3 | Decreased stomatal conductance | [50] | |
| 0.2 Μm/L; 12 days | Arachis hypogaea cv | C3 | Pores become smaller | [51] | |
| 50 mg/kg; 10 days | Glycine max | C3 | Decreased of number stomata | [33] | |
| 5, 10, 30, 50 mg/kg; 7 days | Zea mays L. | C4 | Increased stomatal conductance (high) Decreased stomatal conductance (low) | [52,53] | |
| 100 mg/kg; 28 days | Calendula officinalis | C4 | Decreased stomatal conductance | [8] | |
| 10 μM/L; 8 h | Beta vulgar | C3 | Reduced stomatal aperture and size | [54] | |
| 25, 50 μM/L; 28 days | Bacopa monniera | Decreased stomatal conductance | [49] | ||
| 42 mg/kg; 10 days | Hordeum vulgare L. | C3 | Decreased stomatal conductance | [55] | |
| 100 mg/kg; 30, 60, 90 days | Triticum aestivum L. | C3 | Decreased stomatal conductance | [54] | |
| 10, 50, 100 μM/L; 21 days | Vigna radiata | C3 | Decreased stomatal conductance | [56] | |
| 25, 100 mg/kg; 60, 90 days | Cicer arietinum L. | C3 | Decreased stomatal conductance | [38] | |
| 10, 50, 100 μM/L; 20 days | Cucumis sativus L. | C3 | Decreased stomatal conductance | [57] | |
| 1, 10, 100 μM/L; 15 days | Hordeum vulgare | C3 | Reducing the density and number of stomata | [58] | |
| 250 μM/L; 90 days | Schinus molle | Smaller stomata size | [59] | ||
| 7 μM/L; 40 days | Eichhornia crassipes | C3 | Increased stomatal density | [15] | |
| 30 mg/kg; 42 days | Melissa officinalis L. | Reduced stomatal size and reduced stomatal index | [58] | ||
| 32 μM/L; 15 days | Calophyllum brasiliense | Stomatal closure | [40] | ||
| 100 μM/L; 15 days | Vigna unguiculata (L.) | Decreased stomatal conductance | [41] | ||
| Pb | 30.2 mg/kg | Brassica rapa spp. pekinensis | C3 | Stomatal closure | [60] |
| 700 μM/L; 21 days | Leucaena leucocephala (Lam.) | C3 | Decreased stomatal conductance | [11] | |
| 1000; mg/kg | Triticum aestivum L. | C3 | Decreased stomatal conductance | [30] | |
| 0.5, 1, 2, 4 mg/L; 20 days | Eichhornia crassipes | C3 | Decreased stomatal conductance | [61] | |
| 500 μM/L; 7 days | Nicotiana tabacum L. | C3 | Stomatal closure | [34] | |
| 500, 1000 mg/kg; 30 days | Plantago asiatica L | C3 | Decreased stomatal conductance | [62] | |
| 0.5, 1, 2, 4, 8 μM/L; 30 days | Panicum aquanticum Poir | C3 | Increased stomatal density and Decreased stomatal size | [63] | |
| 40 mg/dm; 14 days | Glycine max (Linn.) Merr | C3 | Stomatal closure | [36] | |
| 1400 mg/kg; 7 days | Ligustrum lucidum Ait. | Decreased stomatal conductance | [47] | ||
| 300 mg; 21 days | Lactuca sativa L. | Decreased stomatal conductance | [64] | ||
| Cu | different concentrations in the air | Gochnatia arequipensis Sandwith | C3 | High stomatal density, small pores. | [65] |
| 25 mg/L; 10 days | Vigna mungo (L.) | C3 | Stomatal restriction | [33] | |
| 10, 100 mg/kg; 20 days | Solanum lycopersicum L. | C3 | Guard cells are destroyed, stomatal closure | [46] | |
| 2, 20, 200 μM/L; 80 days | Billbergia zebrina Lindl. | C3 | Change in stomata density | [15] | |
| 50 μM/L; 14 days | Coriandrum sativum | C3 | Decreased stomatal conductance | [66] | |
| Zn | 1 nM; 25 days | Populus × euramericana | C3 | Changing the number, density of stomata | [64] |
| 400 μg; 6 months | Cajanus cajan (Linn.) Huth | C3 | Decreased stomatal conductance | [12] | |
| 5 μM/L; 20 days | Datura species | C3 | Stomatal closure | [67] | |
| Al | 10 mM; 98 days | Quercus glauca Thumb | C3 | Decreased stomatal conductance | [48] |
| Cr | 150 μM; 20 days | Zea mays L. | C4 | Decreased stomatal conductance | [68] |
| Ba | 5000 μM; 20 days | Glycine max | C3 | Stomatal closure | [17] |
| Ni | 100 ppm; 2 years | Arundo donax L. | C3 | Stomatal conductance declined; increased stomatal resistance | [69] |
| Hg | 1–1000 nM; 49 days | Picea asperata Mast | C3 | Stomatal closure | [41] |
| Sb | 2000 mg/kg; 60 days | Acorus calamus L. | C3 | Decreased stomatal conductance | [47] |
| Ti | 20 mg/L | Quercus ilex subsp ballota | C3 | Decreased stomatal conductance | [70] |
| As | 25 μM; 8 days | Glycine max | C3 | Decreased stomatal conductance | [33,52] |
| Mn | 150 mg/kg; 15 days | Brassica juncea (L.) Czern. | C3 | Stomatal closure | [3] |
| Fe | 100 μM/L; 12 days | Arachis hypogaea cv | C3 | Caused small and abundant stomata on the leaf surface | [52] |
| Ag | 17.7 μM/L; 21 days | Salix miyabeana | C3 | Decreased stomatal conductance | [71] |
4.2. The Effect of Heavy Metals on Stomatal Conductance
4.3. The Effect of Heavy Metals on the Amount and Density of Stomata
4.4. The Effect of Heavy Metals on Stomatal Guard Cells
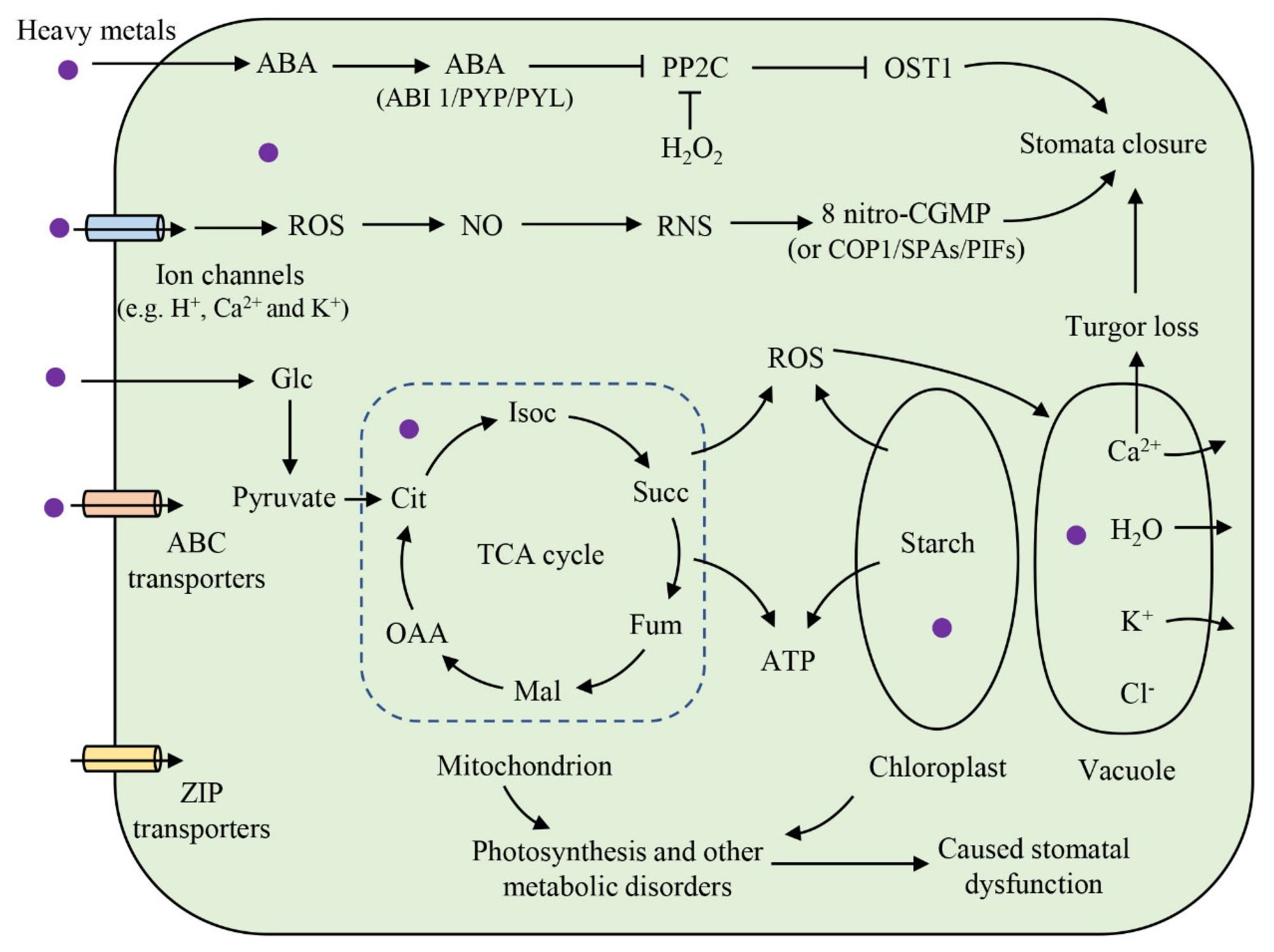
4.5. The Mechanisms of Heavy-Metal-Induced Stomatal Damage
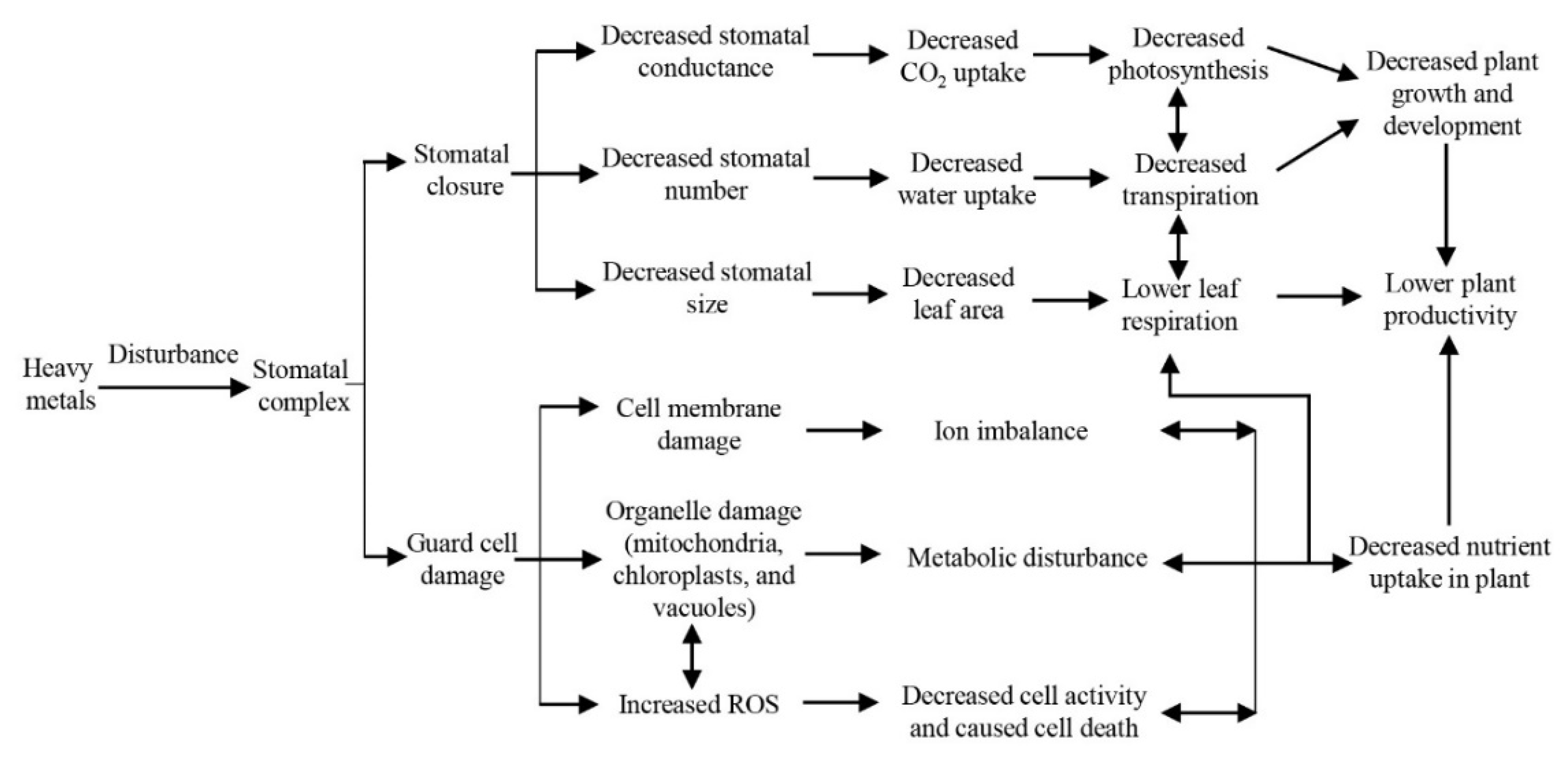
5. Ecological Damage of Whole Plants in Relation to Stomata Response
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barceló, J. Poschenrieder, C. Plant water relations as affected by heavy metal stress: A review. J. Plant Nutr. 1990, 13, 1–37. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Hussain, A.; Nazir, F.; Fariduddin, Q. 24-epibrassinolide and spermidine alleviate Mn stress via the modulation of root morphology, stomatal behavior, photosynthetic attributes and antioxidant defense in Brassica juncea. Physiol. Mol. Biol. Plants 2019, 25, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Farooq, A.; Nadeem, M.; Abbas, G.; Shabbir, A.; Khalid, M.S.; Javeed, H.M.R.; Saeed, M.F.; Akram, A.; Younis, A.; Akhtar, G. Cadmium Partitioning, Physiological and Oxidative Stress Responses in Marigold (Calendula calypso) Grown on Contaminated Soil: Implications for Phytoremediation. Bull. Environ. Contam. Toxicol. 2020, 105, 270–276. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Chen, L. Growth and physiological responses of Pennisetum sp. to cadmium stress under three different soils. Environ. Sci. Pollut. Res. Int. 2021, 28, 14867–14881. [Google Scholar] [CrossRef]
- Casson, S.A.; Hetherington, A.M. Environmental regulation of stomatal development. Curr. Opin. Plant Biol. 2010, 13, 90–95. [Google Scholar] [CrossRef]
- Berry, J.A.; Beerling, D.J.; Franks, P.J. Stomata: Key players in the earth system, past and present. Curr. Opin. Plant Biol. 2010, 13, 233–240. [Google Scholar] [CrossRef]
- Lawson, T.; Vialet-Chabrand, S. Speedy stomata, photosynthesis and plant water use efficiency. New Phytol. 2019, 221, 93–98. [Google Scholar] [CrossRef]
- Henry, C.; John, G.P.; Pan, R.; Bartlett, M.K.; Fletcher, L.R.; Scoffoni, C.; Sack, L. A stomatal safety-efficiency trade-off constrains responses to leaf dehydration. Nat. Commun. 2019, 10, 3398. [Google Scholar] [CrossRef]
- Ali, G.; Srivastava, P.S.; Iqbal, M. Influence of Cadmium and Zinc on Growth and Photosynthesis of Bacopa monniera Cultivated in vitro. Biol. Plant. 2000, 43, 599–601. [Google Scholar] [CrossRef]
- Alkhatib, R.; Mheidat, M.; Abdo, N.; Tadros, M.; Al-Eitan, L.; Al-Hadid, K. Effect of lead on the physiological, biochemical and ultrastructural properties of Leucaena leucocephala. Plant Biol. 2019, 21, 1132–1139. [Google Scholar] [CrossRef]
- Khudsar, T.; Arshi, A.; Siddiqi, T.O.; Mahmooduzzafar; Iqbal, M. Zinc-Induced Changes in Growth Characters, Foliar Properties, and Zn-Accumulation Capacity of Pigeon Pea at Different Stages of Plant Growth. J. Plant Nutr. 2008, 31, 281–306. [Google Scholar] [CrossRef]
- Chen, Z.H.; Chen, G.; Dai, F.; Wang, Y.; Hills, A.; Ruan, Y.L.; Zhang, G.; Franks, P.J.; Nevo, E.; Blatt, M.R. Molecular Evolution of Grass Stomata. Trends Plant Sci. 2017, 22, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Zwieniecki, M.A.; Haaning, K.S.; Boyce, C.K.; Jensen, K.H. Stomatal design principles in synthetic and real leaves. J. R. Soc. Interface 2016, 13, 20160535. [Google Scholar] [CrossRef]
- Shimada, T.; Sugano, S.S.; Hara-Nishimura, I. Positive and negative peptide signals control stomatal density. Cell Mol. Life Sci. CMLS 2011, 68, 2081–2088. [Google Scholar] [CrossRef]
- Von Groll, U.; Berger, D.; Altmann, T. The subtilisin-like serine protease SDD1 mediates cell-to-cell signaling during arabidopsis stomatal development. Plant Cell 2002, 14, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Suwa, R.; Jayachandran, K.; Nguyen, N.T.; Boulenouar, A.; Fujita, K.; Saneoka, H. Barium toxicity effects in soybean plants. Arch. Environ. Contam. Toxicol. 2008, 55, 397–403. [Google Scholar] [CrossRef]
- Martins, J.P.R.; Martins, A.D.; Pires, M.F.; Braga Junior, R.A.; Reis, R.O.; Dias, G.d.M.G.; Pasqual, M. Anatomical and physiological responses of Billbergia zebrina (Bromeliaceae) to copper excess in a controlled microenvironment. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 126, 43–57. [Google Scholar] [CrossRef]
- Wong, S.C.; Cowan, I.R.; Farquhar, G.D. Stomatal conductance correlates with photosynthetic capacity. Nature 1979, 282, 424–426. [Google Scholar] [CrossRef]
- Sussmilch, F.C.; Schultz, J.; Hedrich, R.; Roelfsema, M.R.G. Acquiring Control: The Evolution of Stomatal Signalling Pathways. Trends Plant Sci. 2019, 24, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Serna, L. Stomatal development in Arabidopsis and grasses: Differences and commonalities. Int. J. Dev. Biol. 2011, 55, 5–10. [Google Scholar] [CrossRef]
- Vatén, A.; Bergmann, D.C. Mechanisms of stomatal development: An evolutionary view. Evodevo 2012, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A.; Szarejko, I. Open or close the gate—Stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Dubeaux, G.; Hsu, P.K.; Ceciliato, P.H.O.; Swink, K.J.; Rappel, W.J.; Schroeder, J.I. Deep dive into CO2-dependent molecular mechanisms driving stomatal responses in plants. Plant Physiol. 2021, 187, 2032–2042. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of Stomatal Closure in Plants Exposed to Drought and Cold Stress. Adv. Exp. Med. Biol. 2018, 1081, 215–232. [Google Scholar]
- Driesen, E.; Van den Ende, W.; De Proft, M.; Saeys, W. Influence of Environmental Factors Light, CO2, Temperature, and Relative Humidity on Stomatal Opening and Development: A Review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Yang, J.; Li, C.; Kong, D.; Guo, F.; Wei, H. Light-Mediated Signaling and Metabolic Changes Coordinate Stomatal Opening and Closure. Front. Plant Sci. 2020, 11, 601478. [Google Scholar] [CrossRef]
- Zou, M.; Guo, M.; Zhou, Z.; Wang, B.; Pan, Q.; Li, J.; Zhou, J.M.; Li, J. MPK3- and MPK6-mediated VLN3 phosphorylation regulates actin dynamics during stomatal immunity in Arabidopsis. Nat. Commun. 2021, 12, 6474. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Jing, Y.; Zhang, L.; Kong, D. Phytohormones and their crosstalk in regulating stomatal development and patterning. J. Exp. Bot. 2021, 72, 2356–2370. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Sami, F.; Siddiqui, H.; Hayat, S.J. Supplementation of Salicylic Acid and Citric Acid for Alleviation of Cadmium Toxicity to Brassica juncea. J. Plant Growth Regul. 2019, 39, 641–655. [Google Scholar] [CrossRef]
- Lamoreaux, R.J.; Chaney, W.Q.R. Growth and Water Movement in Silver Maple Seedlings Affected by Cadmium. J. Environ. Quality 1977, 6, 201–205. [Google Scholar] [CrossRef]
- Kanwal, A.; Farhan, M.; Sharif, F.; Hayyat, M.U.; Shahzad, L.; Ghafoor, G.Z. Effect of industrial wastewater on wheat germination, growth, yield, nutrients and bioaccumulation of lead. Sci. Rep. 2020, 10, 11361. [Google Scholar] [CrossRef] [PubMed]
- Gálusová, T.; Piršelová, B.; Rybanský, L.; Krasylenko, Y.; Mészáros, P.; Blehová, A.; Bardáčová, M.; Moravčíková, J.; Matušíková, I. Plasticity of Soybean Stomatal Responses to Arsenic and Cadmium at the Whole Plant Level. Pol. J. Environ. Stud. 2020, 29, 3569–3580. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, R.; Maruthavanan, J.; Ghoshroy, S.; Steiner, R.; Sterling, T.; Creamer, R. Physiological and ultrastructural effects of lead on tobacco. Biol. Plant. 2012, 56, 711–716. [Google Scholar] [CrossRef]
- Bibi, M.; Hussain, M. Effect of copper and lead on photosynthesis and plant pigments in black gram [Vigna mungo (L.) Hepper]. Bull. Environ. Contam. Toxicol. 2005, 74, 1126–1133. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Chwil, M. Lead-Induced Histological and Ultrastructural Changes in the Leaves of Soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr. 2005, 51, 203–212. [Google Scholar] [CrossRef]
- Baruah, S.; Bora, M.S.; Sharma, P.; Deb, P.; Sarma, K.P. Understanding of the Distribution, Translocation, Bioaccumulation, and Ultrastructural Changes of Monochoria hastata Plant Exposed to Cadmium. Water Air Soil Pollut. 2016, 228, 17. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Ahmad, A. Cumulative effects of proline salicylic acid on the cadmium-induced changes in Cicer arietinum L. Fresenius Environ. Bull. 2014, 23, 330–340. [Google Scholar]
- Kaur, R.; Yadav, P.; Thukral, A.K.; Sharma, A.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Castasterone and Citric Acid Supplementation Alleviates Cadmium Toxicity by Modifying Antioxidants and Organic Acids in Brassica juncea. J. Plant Growth Regul. 2017, 37, 286–299. [Google Scholar] [CrossRef]
- Pereira, A.S.; Cortez, P.A.; de Almeida, A.F.; Prasad, M.N.V.; Franca, M.G.C.; da Cunha, M.; de Jesus, R.M.; Mangabeira, P.A.O. Morphology, ultrastructure, and element uptake in Calophyllum brasiliense Cambess. (Calophyllaceae J. Agardh) seedlings under cad-mium exposure. Environ. Sci. Pollut. Res. Int. 2017, 24, 15576–15588. [Google Scholar] [CrossRef]
- Sadeghipour, O. Cadmium Toxicity Alleviates by Seed Priming with Proline or Glycine Betaine in Cowpea (Vigna unguiculata (L.) Walp.). Egypt. J. Agron. 2020, 42, 161–168. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, J.; Zou, J.; Jiang, Z.; Wang, J.; Wu, H.; Jiang, W.; Liu, D. Cadmium uptake and accumulation and its toxic effects on leaves in Hordeum vulgare. Fresenius Environ. Bull. 2015, 24, 4504–4511. [Google Scholar]
- Godbold, D.L.; Plantarum, A. Inhibition of photosynthesis and transpiration in relation to mercury-induced root daage in spruce seedlings. Physiol. Plant. 1988, 74, 270–275. [Google Scholar] [CrossRef]
- Nazir, F.; Fariduddin, Q.; Hussain, A.; Khan, T.A. Brassinosteroid and hydrogen peroxide improve photosynthetic machiery, stomatal movement, root morphology and cell viability and reduce Cu- triggered oxidative burst in tomato. Ecotoxicol. Environ. Saf. 2021, 207, 111081. [Google Scholar] [CrossRef]
- Nazir, F.; Hussain, A.; Fariduddin, Q. Interactive role of epibrassinolide and hydrogen peroxide in regulating stomatal physiology, root morphology, photosynthetic and growth traits in Solanum lycopersicum L. under nickel stress. Environ. Exp. Bot. 2019, 162, 479–495. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, C.; Zhu, P.; Liu, F. Effects of Antimony Stress on Photosynthesis and Growth of Acorus calamus. Front. Plant Sci. 2018, 9, 579. [Google Scholar] [CrossRef] [PubMed]
- Akaya, M.; Takenaka, C.J. Effects of aluminum stress on photosynthesis of Quercus glauca Thumb. Plant Soil 2001, 237, 137–146. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Gunsé, B.; Barceló, J. Influence of cadmium on water relations, stomatal resistance, and abscisic Acid content in expanding bean leaves. Plant Physiol. 1989, 90, 1365–1371. [Google Scholar] [CrossRef]
- Perfus-Barbeoch, L.; Leonhardt, N.; Vavasseur, A.; Forestier, C. Heavy metal toxicity: Cadmium permeates through calcium channels and disturbs the plant water status. Plant J. Cell Mol. Biol. 2002, 32, 539–548. [Google Scholar] [CrossRef]
- Xin, C.; Wang, J.; Shi, Y.; Zhao, M.Q.; Chi, G.Y. Effects of cadmium on growth and photosynthetic activities in pakchoi and mustard. Bot. Stud. 2011, 52, 41–46. [Google Scholar]
- Shi, G.; Sun, L.; Wang, X.; Liu, C. Leaf responses to iron nutrition and low cadmium in peanut: Anatomical properties in relation to gas exchange. Plant Soil 2013, 375, 99–111. [Google Scholar] [CrossRef]
- Vezza, M.E.; Llanes, A.; Travaglia, C.; Agostini, E.; Talano, M.A. Arsenic stress effects on root water absorption in soybean plants: Physiological and morphological aspects. Plant Physiol. Biochem. PPB 2018, 123, 8–17. [Google Scholar] [CrossRef]
- Jia, J.; Gao, Y.; Lu, Y.; Shi, K.; Li, Z.; Wang, S. Trace metal effects on gross primary productivity and its associative environmental risk assessment in a subtropical lake, China. Environ. Pollut. 2020, 259, 113848. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, A.; Lidon, F.C.; Matos, M.D.C.; Ramalho, J.C.; Yordanov, I. Photosynthetic Performance and Content of Some Nutrients in Cadmium- and Copper-Treated Barley Plants. J. Plant Nutr. 2020, 25, 2343–2360. [Google Scholar] [CrossRef]
- Samiullah; Khan, N.A. Physiological basis for reduced photosynthesis and growth of cadmium treated wheat cultivars differing in yield potential. J. Food Agric. Environ. 2007, 5, 375–377. [Google Scholar]
- Wahid, A.; Ghani, A.; Javed, F. Effect of cadmium on photosynthesis, nutrition and growth of mungbean. Agron. Sustain. Dev. 2008, 28, 273–280. [Google Scholar] [CrossRef]
- Sun, S.; Li, M.; Zuo, J.; Jiang, W.; Liu, D. Cadmium effects on mineral accumulation, antioxidant defence system and gas exchange in cucumber. Zemdirb.-Agric. 2015, 102, 193–200. [Google Scholar] [CrossRef]
- Kilic, S.; Kilic, M. Effects of cadmium-induced stress on essential oil production, morphology and physiology of lemon balm (Melissa officinalis L. Lamiaceae). Appl. Ecol. Environ. Res. 2017, 15, 1653–1669. [Google Scholar] [CrossRef]
- Gao, P.P.; Xue, P.Y.; Dong, J.W.; Zhang, X.M.; Sun, H.X.; Geng, L.P.; Luo, S.X.; Zhao, J.J.; Liu, W.J. Contribution of PM2.5-Pb in atmospheric fallout to Pb accumulation in Chinese cabbage leaves via stomata. J. Hazard. Mater. 2020, 407, 124356. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.J.; Castro, E.M.; Oliveira, C.D.; Pires, M.F.; Pereira, M.P.; Ramos, S.J.; Faquin, V. Lead tolerance of water hyacinth (Eichhornia crassipes Mart.-Pontederiaceae) as defined by anatomical and physiological traits. An. Da Acad. Bras. De Ciências 2014, 86, 1423–1433. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Z.; Zhang, Y.; Wei, Y.; Jiang, Z. Effects of lead stress on the growth, physiology, and cellular structure of privet seedlings. PLoS ONE 2018, 13, e0191139. [Google Scholar] [CrossRef] [PubMed]
- Kosobrukhov, A.; Knyazeva, I.; Mudrik, V. Plantago major plants responses to increase content of lead in soil: Growth and photosynthesis. Plant Growth Regul. 2004, 42, 145–151. [Google Scholar] [CrossRef]
- Pires-Lira, M.F.; de Castro, E.M.; Lira, J.M.S.; de Oliveira, C.; Pereira, F.J.; Pereira, M.P. Potential of Panicum aquanticum Poir. (Poaceae) for the phytoremediation of aquatic environments contaminated by lead. Ecotoxicol. Environ. Saf. 2020, 193, 110336. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Zhang, T.; Dumat, C.; Sobanska, S.; Dappe, V.; Shahid, M.; Xian, Y.; Li, X.; Li, S. Airborne foliar transfer of particular metals in Lactuca sativa L.: Translocation, phytotoxicity, and bioaccessibility. Environ. Sci. Pollut. Res. Int. 2019, 26, 20064–20078. [Google Scholar] [CrossRef] [PubMed]
- Machacal, D.A.A.; Valdivia, B.R.M.; Vargas, F.F.R.; Oviedo, R.D.N. Toxicidad del cobre sobre la morfología de estomas de Gochnatia arequipensis Sandwith (Asteraceae) de dos localidades de Arequipa, Perú. Idesia (Arica) 2019, 37, 81–87. [Google Scholar] [CrossRef]
- Vaillant, N.; Monnet, F.; Hitmi, A.; Sallanon, H.; Coudret, A. Comparative study of responses in four Datura species to a zinc stress. Chemosphere 2005, 59, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Baccio, D.D.; Minnocci, A.; Sebastiani, L. Leaf structural modifications in Populus × euramericana subjected to Zn excess. Biol. Plant. 2010, 54, 502–508. [Google Scholar] [CrossRef]
- Anjum, S.A. Chromium Toxicity Induced Alterations in Growth, Photosynthesis, Gas Exchange Attributes and Yield Formation in Maize. Pak. J. Agric. Sci. 2016, 53, 751–757. [Google Scholar]
- Papazoglou, E.G.; Serelis, K.G.; Bouranis, D.L. Impact of high cadmium nickel soil concentration on selected physiological parameters of Arundo donax L. Eur. J. Soil Biol. 2007, 43, 207–215. [Google Scholar] [CrossRef]
- Dominguez, M.T.; Maranon, T.; Murillo, J.M.; Redondo-Gomez, S. Response of Holm oak (Quercus ilex subsp. ballota) and mastic shrub (Pistacia lentiscus L.) seedlings to high concentrations of Cd and Tl in the rhizosphere. Chemosphere 2011, 83, 1166–1174. [Google Scholar] [CrossRef]
- Nissim, W.G.; Pitre, F.E.; Kadri, H.; Desjardins, D.; Labrecque, M. Early response of willow to increasing silver concentration exposure. Int. J. Phytoremediation 2014, 16, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Qi, L.; Jiang, W.; Liu, D. Cadmium accumulation and its effects on growth and gas exchange in four Populus cultivars. Acta Biol. Cracoviensia. Ser. Bot. 2007, 49, 7–14. [Google Scholar]
- Jia, Z.; Sinicae, C. Photosynthetic Responses of the Riparian Salix variegata to Cadmium Stress in Three Gorges Reservoir Region. Entia Silvae Sin. 2012, 48, 152–158. [Google Scholar]
- Stancheva, I.; Geneva, M.; Markovska, Y.; Tzvetkova, N.; Mitova, I.; Todorova, M.; Petrov, P. A comparative study on plant morphology, gas exchange parameters, and antioxidant response of Ocimum basilicum L. and Origanum vulgare L. grown on industrially polluted soil. Turk. J. Biol. 2014, 38, 89–102. [Google Scholar] [CrossRef]
- Davies, F.T.; Puryear, J.D.; Newton, R.J.; Egilla, J.N.; Grossi, J.A.S. Mycorrhizal fungi increase chromium uptake by sunflower plants: Influence on tissue mineral concentration, growth, and gas exchange. J. Plant Nutr. 2002, 25, 2389–2407. [Google Scholar] [CrossRef]
- Zaouali, W.; Mahmoudi, H.; Salah, I.B.; Mejri, F.; Casabianca, H.; Hosni, K.; Ouerghi, Z. Copper-induced changes in growth, photosynthesis, antioxidative system activities and lipid metabolism of cilantro (Coriandrum sativum L.). Biologia 2020, 75, 367–380. [Google Scholar] [CrossRef]
- Fox, J.-P.; Capen, J.D.; Zhang, W.; Ma, X.; Rossi, L. Effects of cerium oxide nanoparticles and cadmium on corn (Zea mays L.) seedlings physiology and root anatomy. NanoImpact 2020, 20, 100264. [Google Scholar] [CrossRef]
- Pereira, F.J.; de Castro, E.M.; Pires, M.F.; Oliveira, C.D.; Pasqual, M. Anatomical and physiological modifications in water hyacinth under cadmium contamination. J. Appl. Bot. Food Qual. 2017, 90, 10–17. [Google Scholar]
- Orcen, N. Stomatal Parameters and Growth Responses of Tomatal Nicotiana and Atriplex to Cd, Pb and Cd-Pb-Contaminated Soil. Fresenius Environ. Bull. 2017, 26, 6340–6345. [Google Scholar]
- Greger, M.; Johansson, M. Cadmium effects on leaf transpiration of sugar beet (Beta vulgaris). Physiol. Plant. 1992, 86, 465–473. [Google Scholar] [CrossRef]
- Nazir, A.; Rafique, F.; Ahmed, K.; Khan, S.A.; Khan, N.; Akbar, M.; Zafar, M. Evaluation of heavy metals effects on morpho-anatomical alterations of wheat (Triticum aestivum L.) seedlings. Microsc. Res. Tech. 2021, 84, 2517–2529. [Google Scholar] [CrossRef]
- Nazir, F.; Hussain, A.; Fariduddin, Q. Hydrogen peroxide modulate photosynthesis and antioxidant systems in tomato (Solanum lycopersicum L.) plants under copper stress. Chemosphere 2019, 230, 544–558. [Google Scholar] [CrossRef]
- Rucińska-Sobkowiak, R. Water relations in plants subjected to heavy metal stresses. Acta Physiol. Plant. 2016, 38, 257. [Google Scholar] [CrossRef]
- Kastori, R.; Petrovi, M.; Petrovi, N. Effect of excess lead, cadmium, copper, and zinc on water relations in sunflower. J. Plant Nutr. 1992, 15, 2427–2439. [Google Scholar] [CrossRef]
- Eleftheriou, E.P.; Michalopoulou, V.A.; Adamakis, I.D. Aberration of mitosis by hexavalent chromium in some Fabaceae members is mediated by species-specific microtubule disruption. Environ. Sci. Pollut. Res. Int. 2015, 22, 7590–7599. [Google Scholar] [CrossRef]
- Xu, P.; Liu, D.; Jiang, W. Cadmium effects on the organization of microtubular cytoskeleton in interphase and mitotic cells of Allium sativum. Biol. Plant. 2009, 53, 387–390. [Google Scholar] [CrossRef]
- Ying, R.R.; Qiu, R.L.; Tang, Y.T.; Hu, P.J.; Qiu, H.; Chen, H.R.; Shi, T.H.; Morel, J.L. Cadmium tolerance of carbon assimilation enzymes and chloroplast in Zn/Cd hyperaccumulator Picris divaricata. J. Plant Physiol. 2010, 167, 81–87. [Google Scholar] [CrossRef]
- Hills, A.; Chen, Z.H.; Amtmann, A.; Blatt, M.R.; Lew, V.L. OnGuard, a computational platform for quantitative kinetic modeling of guard cell physiology. Plant Physiol. 2012, 159, 1026–1042. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Böhmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef]
- Tran, T.A. Cadmium-induced structural disturbances in pea leaves are alleviated by nitric oxide. Turk. J. Bot. 2013, 37, 698–707. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q. Leaf plasticity in peanut (Arachis hypogaea L.) in response to heavy metal stress. Environ. Exp. Bot. 2009, 67, 112–117. [Google Scholar] [CrossRef]
- Barceló, P.D.J.; Vaschez, M.D.; Poschenrieder, C. Cadmium-Induced Structural and Ultrastructural Changes in the Vascular System of Bush Bean Stems. Bot. Acta 1988, 101, 254–261. [Google Scholar] [CrossRef]
- Gupta, P.; Bhatnagar, A.K.J.E.; Botany, E. Spatial distribution of arsenic in different leaf tissues and its effect on structure and development of stomata and trichomes in mung bean, Vigna radiata (L.) Wilczek. Environ. Exp. Bot. 2015, 109, 12–22. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, D.; Yang, W.; Liu, Y.; Zhang, L.; Gao, S. Sex-specific responses of Populus deltoides to Glomus intraradices colonization and Cd pollution. Chemosphere 2016, 155, 196–206. [Google Scholar] [CrossRef]
- Kültz, D. Molecular and evolutionary basis of the cellular stress response. Annu. Rev. Physiol. 2005, 67, 225–257. [Google Scholar] [CrossRef]
- Bhaduri, A.M.; Fulekar, M. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Bio/Technol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Kalinowska, R.; Pawlik-Skowrońska, B. Response of two terrestrial green microalgae (Chlorophyta, Trebouxiophyceae) isolated from Cu-rich and unpolluted soils to copper stress. Environ. Pollut. 2010, 158, 2778–2785. [Google Scholar] [CrossRef]
- Martins, L.L.; Mourato, M.P.; Cardoso, A.I.; Pinto, A.P.; Mota, A.M.; Gonçalves, M.; Plantarum, A. Oxidative stress induced by cadmium in Nicotiana tabacum L.: Effects on growth parameters, oxidative damage and antioxidant responses in different plant parts. Acta Physiol. Plant. 2011, 33, 1375–1383. [Google Scholar] [CrossRef]
- Benzarti, S.; Hamdi, H.; Mohri, S.; Ono, Y. Response of antioxidative enzymes and apoplastic bypass transport in Thlaspi caerulescens and Raphanus sativus to cadmium stress. Int. J. Phytoremediation 2010, 12, 733–744. [Google Scholar] [CrossRef]
- Tao, L.; Min, Q.; Yang, Z.; Hui, L.; Ping, F.; Fen, T.; Tang, X. Effect of Cadmium Stress on the Growth, Antioxidative Enzymes and Lipid Peroxidation in Two Kenaf (Hibiscus cannabinus L.) Plant Seedlings. J. Integr. Agric. 2013, 12, 610–620. [Google Scholar]
- Zhang, F.Q.; Wang, Y.S.; Lou, Z.P.; Dong, J.D. Effect of heavy metal stress on antioxidative enzymes and lipid peroxidation in leaves and roots of two mangrove plant seedlings (Kandelia candel and Bruguiera gymnorrhiza). Chemosphere 2007, 67, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jin, Z.Q.; Zhao, J.; Zhang, G.; Wu, F. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. Int. J. Nat. Synth. Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Meng, D.; Zhao, Y.; Lei, C.; Zhou, X. Physiological Mechanism of Cd-tolerance of Artemisia Selengensis Grown in Dongting Lake Wetland. Asian J. Ecotoxicol. 2013, 8, 111–120. [Google Scholar]
- Kieffer, P.; Schröder, P.; Dommes, J.; Hoffmann, L.; Renaut, J.; Hausman, J.F. Proteomic and enzymatic response of poplar to cadmium stress. J. Proteom. 2009, 72, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.; Pandey, P.; Rajpoot, R.; Rani, A.; Dubey, R.S. Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings. Protoplasma 2014, 251, 1047–1065. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic signaling during abiotic stress combination in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 13810–13820. [Google Scholar] [CrossRef]
- Mahmood, T.; Khalid, S.; Abdullah, M.; Ahmed, Z.; Shah, M.K.N.; Ghafoor, A.; Du, X. Insights into Drought Stress Signaling in Plants and the Molecular Genetic Basis of Cotton Drought Tolerance. Cells 2019, 9, 105. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research Progress and Perspective on Drought Stress in Legumes: A Review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Meng, Z.; Wang, B.; Chen, M. Research Progress on the Roles of Cytokinin in Plant Response to Stress. Int. J. Mol. Sci. 2020, 21, 6574. [Google Scholar] [CrossRef]
- Koua, A.P.; Oyiga, B.C.; Baig, M.M.; Leon, J.; Ballvora, A. Breeding Driven Enrichment of Genetic Variation for Key Yield Components and Grain Starch Content Under Drought Stress in Winter Wheat. Front. Plant Sci. 2021, 12, 684205. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, J. Rift over biodiversity divides ecologists. Science 2000, 289, 1282–1283. [Google Scholar] [CrossRef]
- Mulenga, C.; Clarke, C.; Meincken, M. Physiological and Growth Responses to Pollutant-Induced Biochemical Changes in Plants: A Review. Pollution 2020, 6, 827–848. [Google Scholar]
- Drenovsky, R.E.; Grewell, B.J.; D’Antonio, C.M.; Funk, J.L.; James, J.J.; Molinari, N.; Parker, I.M.; Richards, C.L. A functional trait perspec tive on plant invasion. Ann. Bot. 2012, 110, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.H.; de Bello, F.; Boulangeat, I.; Pellet, G.; Lavorel, S.; Thuiller, W. On the importance of intraspecific variability for the quantification of functional diversity. Oikos 2012, 121, 116–126. [Google Scholar] [CrossRef]
- Li, G.; Yang, D.; Sun, S. Allometric relationships between lamina area, lamina mass and petiole mass of 93 temperate woody species vary with leaf habit, leaf form and altitude. Funct. Ecol. 2008, 22, 557–564. [Google Scholar] [CrossRef]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Steege, H.T.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- McIntyre, S.; Lavorel, S.; Landsberg, J.; Forbes, T.D.A. Disturbance response in vegetation—Towards a global perspective on functional traits. J. Veg. Sci. 2009, 10, 621–630. [Google Scholar] [CrossRef]
- Campoy, J.; Campos, I.; Plaza, C.; Calera, M.; Jiménez, N.; Bodas, V.; Calera, A. Water use efficiency and light use efficiency in garlic using a remote sensing-based approach. Agric. Water Manag. 2019, 219, 40–48. [Google Scholar] [CrossRef]
- Tesfaye, K.; Walker, S.; Tsubo, M. Radiation interception and radiation use efficiency of three grain legumes under water deficit conditions in a semi-arid environment. Eur. J. Agron. 2006, 25, 60–70. [Google Scholar] [CrossRef]
- Hamanishi, E.T.; Thomas, B.R.; Campbell, M.M. Drought induces alterations in the stomatal development program in Populus. J. Exp. Bot. 2012, 63, 4959–4971. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Y.; Wang, X.; Yang, Q.; Quan, X.; Zeng, J.; Dai, F.; Zeng, F.; Wu, F.; Zhang, G.; et al. Leaf epidermis transcriptome reveals drought-Induced hormonal signaling for stomatal regulation in wild barley. Plant Growth Regul. 2018, 87, 39–54. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.A.; Chen, Y.; Ponce, G.; Acevedo, F.E.; Lynch, J.P.; Anderson, C.T.; Ali, J.G.; Felton, G.W. Stomata-mediated interactions between plants, herbivores, and the environment. Trends Plant Sci. 2021, 27, 287–300. [Google Scholar] [CrossRef]
- Miranda, Y.H.; Negron, J. Heavy metals and gene expression in plant stomata. FASEB J. 2015, 29, 887.19. [Google Scholar]
- Chowdhury, M.R.; Ahamed, M.S.; Mas-Ud, M.A.; Islam, H.; Fatamatuzzohora, M.; Hossain, M.F.; Billah, M.; Hossain, M.S.; Matin, M.N. Stomatal development genetic expression in Arabidopsis thaliana L. Heliyon 2021, 7, e07889. [Google Scholar] [CrossRef]
- Gavassi, M.A.; Silva, G.S.; da Silva, C.d.M.S.; Thompson, A.J.; Macleod, K.; Oliveira, P.M.R.; Cavalheiro, M.F.; Domingues, D.S.; Habermann, G. NCED expression is related to increased ABA biosynthesis and stomatal closure under aluminum stress. Environ. Exp. Bot. 2021, 185, 104404. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Gao, Y.; Yuan, X.; Yuan, M.; Huang, L.; Wang, S.; Liu, C.; Duan, C. Effects of Heavy Metals on Stomata in Plants: A Review. Int. J. Mol. Sci. 2023, 24, 9302. https://doi.org/10.3390/ijms24119302
Guo Z, Gao Y, Yuan X, Yuan M, Huang L, Wang S, Liu C, Duan C. Effects of Heavy Metals on Stomata in Plants: A Review. International Journal of Molecular Sciences. 2023; 24(11):9302. https://doi.org/10.3390/ijms24119302
Chicago/Turabian StyleGuo, Zhaolai, Yuhan Gao, Xinqi Yuan, Mengxiang Yuan, Lv Huang, Sichen Wang, Chang’e Liu, and Changqun Duan. 2023. "Effects of Heavy Metals on Stomata in Plants: A Review" International Journal of Molecular Sciences 24, no. 11: 9302. https://doi.org/10.3390/ijms24119302
APA StyleGuo, Z., Gao, Y., Yuan, X., Yuan, M., Huang, L., Wang, S., Liu, C., & Duan, C. (2023). Effects of Heavy Metals on Stomata in Plants: A Review. International Journal of Molecular Sciences, 24(11), 9302. https://doi.org/10.3390/ijms24119302