
Differential Functional Contribution of BK Channel Subunits to Aldosterone-Induced Channel Activation in Vascular Smooth Muscle and Eventual Cerebral Artery Dilation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
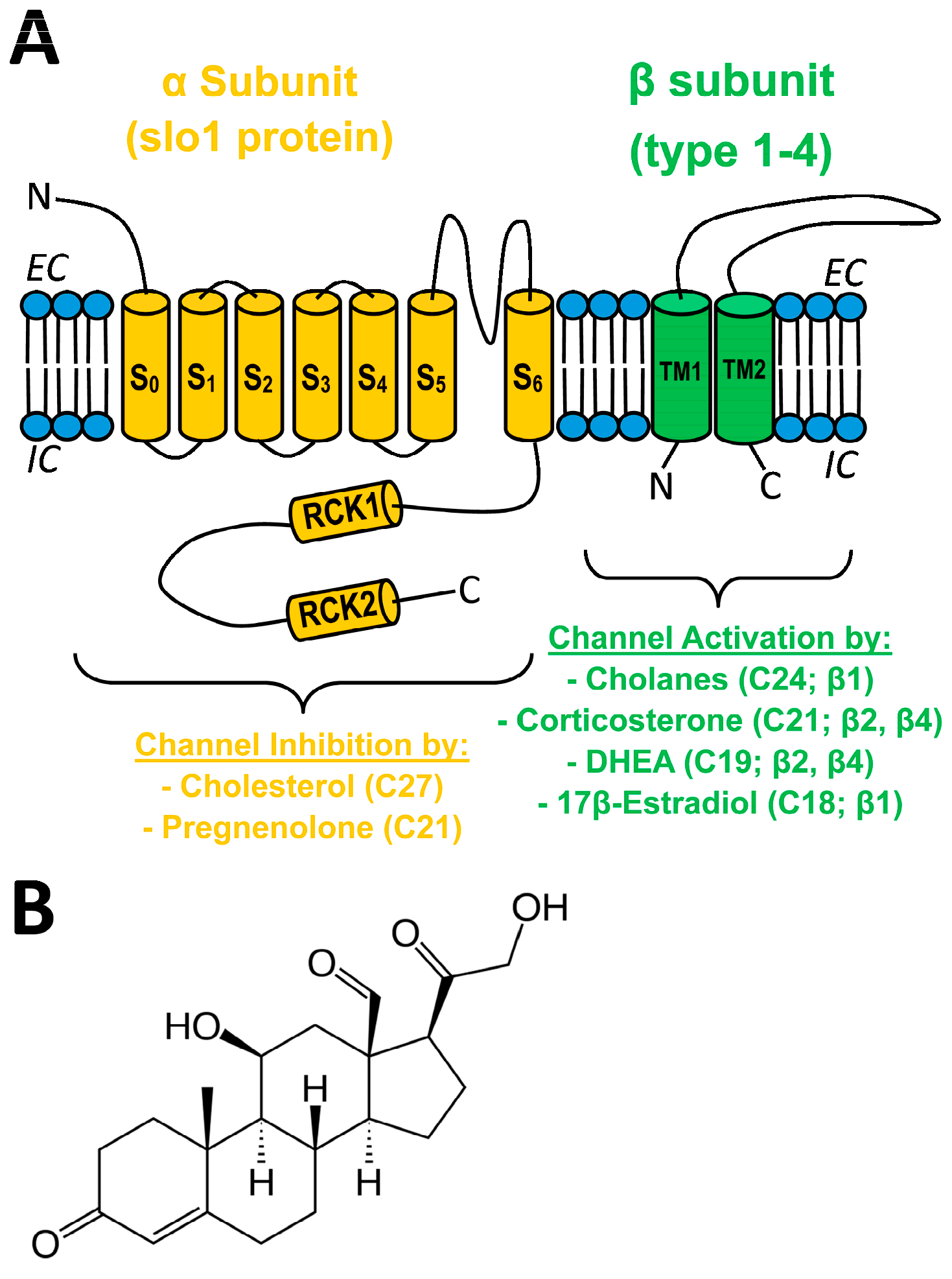
2.1. Aldosterone Binds onto BK α and β1 Subunits with Different Affinity
2.2. Activation of Cerebrovascular Smooth Muscle BK Channels by µM Aldosterone Requires BK β1 Subunits
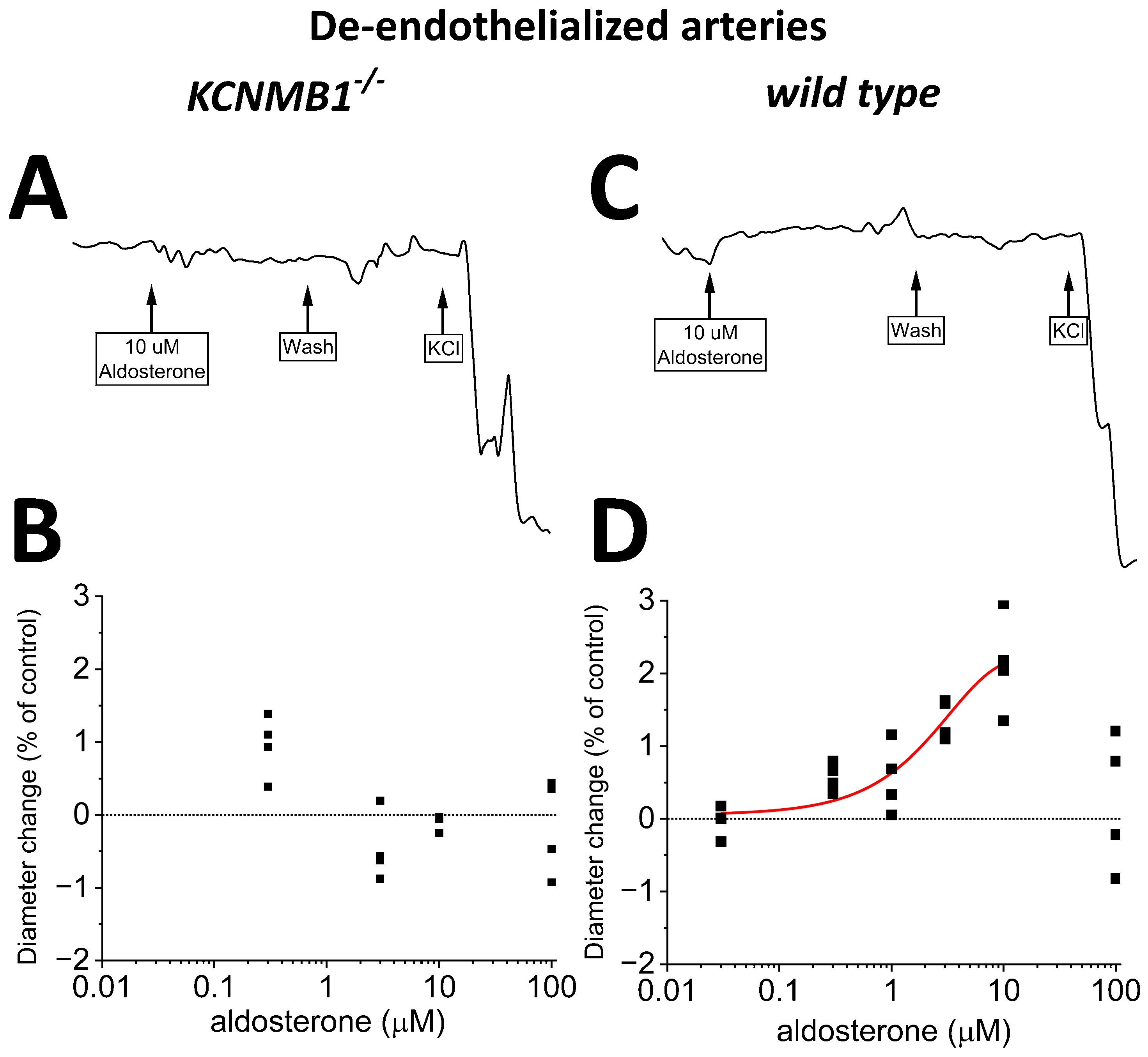
2.3. Aldosterone at Concentrations Well above Physiological Levels Mildly Dilates Middle Cerebral Arteries Independently of Circulating, Metabolic, and Endothelial Factors, an Action That Requires BK β1 Subunits
2.4. Endothelium Sustains Aldosterone-Induced Dilation of Mouse MCA
3. Discussion
4. Materials and Methods
4.1. Immunoprecipitation and Nanoscale Differential Scanning Fluorimetry
4.2. Isolation of Mouse Middle Cerebral Artery Segments and Individual Myocytes
4.3. Electrophysiology Data Acquisition and Analysis
4.4. Middle Cerebral Artery Diameter In Vitro Measurements
4.5. Chemicals
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barrantes, F.J.; Antollini, S.S.; Bouzat, C.B.; Garbus, I.; Massol, R.H. Nongenomic effects of steroids on the nicotinic acetylcholine receptor. Kidney Int. 2000, 57, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Rosenhouse-Dantsker, A.; Noskov, S.; Durdagi, S.; Logothetis, D.E.; Levitan, I. Identification of Novel Cholesterol-binding Regions in Kir2 Channels. J. Biol. Chem. 2013, 288, 31154–31164. [Google Scholar] [CrossRef] [PubMed]
- McMillan, J.E.; Bukiya, A.N.; Terrell, C.L.; Patil, S.A.; Miller, D.D.; Dopico, A.M.; Parrill, A.L. Multi-generational pharmacophore modeling for ligands to the cholane steroid-recognition site in the β1 modulatory subunit of the BKCa channel. J. Mol. Graph. Model. 2014, 54, 174–183. [Google Scholar] [CrossRef] [PubMed]
- North, K.C.; Slayden, A.V.; Mysiewicz, S.; Bukiya, A.N.; Dopico, A.M. Celastrol Dilates and Counteracts Ethanol-Induced Constriction of Cerebral Arteries. Experiment 2020, 375, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Pařízek, A.; Šimják, P.; Koucký, M.; Anderlová, K.; Krejčí, H.; Vejražková, D.; Ondřejíková, L.; Černý, A.; Kancheva, R. Steroids, steroid associated substances and gestational diabetes mellitus. Physiol. Res. 2021, 70 (Suppl. 4), S617–S634. [Google Scholar] [CrossRef]
- Slayden, A.V.; Dyer, C.L.; Ma, D.; Li, W.; Bukiya, A.N.; Parrill, A.L.; Dopico, A.M. Discovery of agonist–antagonist pairs for the modulation of Ca[2]+ and voltage-gated K+ channels of large conductance that contain beta1 subunits. Bioorganic. Med. Chem. 2022, 68, 116876. [Google Scholar] [CrossRef]
- Dopico, A.M.; Bukiya, A.N. Lipid regulation of BK channel function. Front. Physiol. 2014, 5, 312. [Google Scholar] [CrossRef]
- Bukiya, A.N.; Dopico, A.M. Common structural features of cholesterol binding sites in crystallized soluble proteins. J. Lipid Res. 2017, 58, 1044–1054. [Google Scholar] [CrossRef]
- Jiang, Q.-X.; Levitan, I. Principles of cholesterol regulation of ion channels. In Cholesterol; Bukiya, A.N., Dopico, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 169–204. [Google Scholar]
- Dopico, A.M.; Bukiya, A.N.; Jaggar, J.H. Calcium- and voltage-gated BK channels in vascular smooth muscle. Pflügers Arch. -Eur. J. Physiol. 2018, 470, 1271–1289. [Google Scholar] [CrossRef]
- Salkoff, L.; Butler, A.; Ferreira, G.; Santi, C.; Wei, A. High-conductance potassium channels of the SLO family. Nat. Rev. Neurosci. 2006, 7, 921–931. [Google Scholar] [CrossRef]
- Latorre, R.; Castillo, K.; Carrasquel-Ursulaez, W.; Sepulveda, R.V.; Gonzalez-Nilo, F.; Gonzalez, C.; Alvarez, O. Molecular Determinants of BK Channel Functional Diversity and Functioning. Physiol. Rev. 2017, 97, 39–87. [Google Scholar] [CrossRef]
- Orio, P.; Rojas, P.; Ferreira, G.; Latorre, R. New disguises for an old channel: MaxiK channel beta-subunits. News Physiol. Sci. 2002, 17, 156–161. [Google Scholar] [CrossRef]
- Gonzalez-Perez, V.; Lingle, C.J. Regulation of BK Channels by Beta and Gamma Subunits. Annu. Rev. Physiol. 2019, 81, 113–137. [Google Scholar] [CrossRef]
- Dopico, A.M.; Walsh, J.V.; Singer, J.J. Natural Bile Acids and Synthetic Analogues Modulate Large Conductance Ca2+-activated K+ (BKCa) Channel Activity in Smooth Muscle Cells. J. Gen. Physiol. 2002, 119, 251–273. [Google Scholar] [CrossRef]
- Bukiya, A.N.; Vaithianathan, T.; Kuntamallappanavar, G.; Asuncion-Chin, M.; Dopico, A.M. Smooth Muscle Cholesterol Enables BK β1 Subunit-Mediated Channel Inhibition and Subsequent Vasoconstriction Evoked by Alcohol. Arter. Thromb. Vasc. Biol. 2011, 31, 2410–2423. [Google Scholar] [CrossRef]
- Granados, S.T.; Castillo, K.; Bravo-Moraga, F.; Sepúlveda, R.V.; Carrasquel-Ursulaez, W.; Rojas, M.; Carmona, E.; Lorenzo-Ceballos, Y.; González-Nilo, F.; González, C.; et al. The molecular nature of the 17β-Estradiol binding site in the voltage- and Ca2+-activated K+ (BK) channel β1 subunit. Sci. Rep. 2019, 9, 9965. [Google Scholar] [CrossRef] [PubMed]
- King, J.T.; Lovell, P.V.; Rishniw, M.; Kotlikoff, M.I.; Zeeman, M.L.; McCobb, D.P. Beta2 and beta4 subunits of BK channels confer differential sensitivity to acute modulation by steroid hormones. J. Neurophysiol. 2006, 95, 2878–2888. [Google Scholar] [CrossRef]
- North, K.C.; Bukiya, A.N.; Dopico, A.M. BK channel-forming slo1 proteins mediate the brain artery constriction evoked by the neurosteroid pregnenolone. Neuropharmacology 2021, 192, 108603. [Google Scholar] [CrossRef] [PubMed]
- Crowley, J.J.; Treistman, S.N.; Dopico, A.M. Cholesterol Antagonizes Ethanol Potentiation of Human Brain BKCa Channels Reconstituted into Phospholipid Bilayers. Mol. Pharmacol. 2003, 64, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; McMillan, J.; Bukiya, A.N.; Burton, B.; Parrill, A.L.; Dopico, A.M. Multiple Cholesterol Recognition/Interaction Amino Acid Consensus (CRAC) Motifs in Cytosolic C Tail of Slo1 Subunit Determine Cholesterol Sensitivity of Ca2+- and Voltage-gated K+ (BK) Channels. J. Biol. Chem. 2012, 287, 20509–20521. [Google Scholar] [CrossRef]
- Schiffrin, E.L. Effects of Aldosterone on the Vasculature. Hypertension 2006, 47, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Bader, M. Tissue Renin-Angiotensin-Aldosterone Systems: Targets for Pharmacological Therapy. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 439–465. [Google Scholar] [CrossRef]
- Dinh, Q.N.; Arumugam, T.V.; Young, M.J.; Drummond, G.R.; Sobey, C.G.; Chrissobolis, S. Aldosterone and the mineralocorticoid receptor in the cerebral circulation and stroke. Exp. Transl. Stroke Med. 2012, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Strömstedt, M.; Waterman, M.R. Messenger RNAs encoding steroidogenic enzymes are expressed in rodent brain. Mol. Brain Res. 1995, 34, 75–88. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.; Clark, C.; Fraser, R.; Gomez-Sanchez, C.; Connell, J.; Davies, E. Expression of 11beta-hydroxylase and aldosterone synthase genes in the rat brain. J. Mol. Endocrinol. 2000, 24, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Rigsby, C.S.; Burch, A.E.; Ogbi, S.; Pollock, D.M.; Dorrance, A.M. Intact female stroke-prone hypertensive rats lack responsiveness to mineralocorticoid receptor antagonists. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1754–R1763. [Google Scholar] [CrossRef]
- Ghafar, M.T.A. An overview of the classical and tissue-derived renin-angiotensin-aldosterone system and its genetic polymorphisms in essential hypertension. Steroids 2020, 163, 108701. [Google Scholar] [CrossRef]
- Takeda, R.; Matsubara, T.; Miyamori, I.; Hatakeyama, H.; Morise, T.; The Research Committee of Disorders of Adrenal Hormones in Japan. Vascular complications in patients with aldosterone producing adenoma in Japan: Comparative study with essential hypertension. J. Endocrinol. Investig. 1995, 18, 370–373. [Google Scholar] [CrossRef]
- Milliez, P.; Girerd, X.; Plouin, P.-F.; Blacher, J.; Safar, M.E.; Mourad, J.-J. Evidence for an increased rate of cardiovascular events in patients with primary aldosteronism. J. Am. Coll. Cardiol. 2005, 45, 1243–1248. [Google Scholar] [CrossRef]
- Gomez-Sanchez, E.P.; Ahmad, N.; Romero, D.G.; Gomez-Sanchez, C.E. Origin of Aldosterone in the Rat Heart. Endocrinology 2004, 145, 4796–4802. [Google Scholar] [CrossRef]
- Gasparini, S.; Melo, M.; Andrade-Franzé, G.; Geerling, J.; Menani, J.; Colombari, E. Aldosterone infusion into the 4th ventricle produces sodium appetite with baroreflex attenuation independent of renal or blood pressure changes. Brain Res. 2018, 1698, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Grimm, P.R.; Irsik, D.L.; Settles, D.C.; Holtzclaw, J.D.; Sansom, S.C. Hypertension of Kcnmb1/is linked to deficient K secretion and aldosteronism. Proc. Natl. Acad. Sci. USA 2009, 106, 11800–11805. [Google Scholar] [CrossRef] [PubMed]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S.V. Stroke risk factors, genetics, and prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef]
- Jerabek-Willemsen, M.; Wienken, C.J.; Braun, D.; Baaske, P.; Duhr, S. Molecular Interaction Studies Using Microscale Thermophoresis. ASSAY Drug Dev. Technol. 2011, 9, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Bukiya, A.N.; Liu, J.; Toro, L.; Dopico, A.M. Beta1 (KCNMB1) subunits mediate lithocholate activation of large-conductance Ca2+-activated K+ channels and dilation in small, resistance-size arteries. Mol. Pharmacol. 2007, 72, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Jaggar, J.H.; Li, A.; Parfenova, H.; Liu, J.; Umstot, E.S.; Dopico, A.M.; Leffler, C.W. Heme is a carbon monoxide receptor for large-conductance Ca2+-activated K+ channels. Circ. Res. 2005, 97, 805–812. [Google Scholar] [CrossRef]
- Seidel, S.A.I.; Dijkman, P.M.; Lea, W.A.; van den Bogaart, G.; Jerabek-Willemsen, M.; Lazic, A.; Joseph, J.S.; Srinivasan, P.; Baaske, P.; Simeonov, A.; et al. Microscale thermophoresis quantifies biomolecular interactions under previously challenging conditions. Methods 2013, 59, 301–315. [Google Scholar] [CrossRef]
- Pérez, G.J.; Bonev, A.D.; Nelson, M.T. Micromolar Ca(2+) from sparks activates Ca(2+)-sensitive K(+) channels in rat cerebral artery smooth muscle. Am. J. Physiol. Cell Physiol. 2001, 281, C1769–C1775. [Google Scholar] [CrossRef]
- Lee, R.M. Morphology of cerebral arteries. Pharmacol. Ther. 1995, 66, 149–173. [Google Scholar] [CrossRef]
- Lehecka, M.; Dashti, R.; Rinne, J.; Romani, R.; Kivisaari, R.; Niemelä, M.; Hernesniemi, J. Surgical Management of Aneurysms of the Middle Cerebral Artery. In Schmidek and Sweet Operative Neurosurgical Techniques; Saunders: Stanford, CA, USA, 2012; pp. 897–913. [Google Scholar] [CrossRef]
- Cipolla, M.J.; Curry, A.B. Middle cerebral artery function after stroke: The threshold duration of reperfusion for myogenic activity. Stroke 2002, 33, 2094–2099. [Google Scholar] [CrossRef]
- Delgado, M.G.; Bogousslavsky, J. Superficial Middle Cerebral Artery Territory Infarction. Front. Neurol. Neurosci. 2012, 30, 111–114. [Google Scholar] [CrossRef]
- Krafft, P.R.; Bailey, E.L.; Lekic, T.; Rolland, W.B.; Altay, O.; Tang, J.; Wardlaw, J.M.; Zhang, J.H.; Sudlow, C.L.M. Etiology of Stroke and Choice of Models. Int. J. Stroke 2012, 7, 398–406. [Google Scholar] [CrossRef]
- Márquez-Rodas, I.; Xavier, F.E.; Arroyo-Villa, I.; Longo, F.; Aras-Lopez, R.; Blanco-Rivero, J.; Ferrer, M.; Balfagón, G. Increased expression in calcitonin-like receptor induced by aldosterone in cerebral arteries from spontaneously hypertensive rats does not correlate with functional role of CGRP receptor. Regul. Pept. 2008, 146, 125–130. [Google Scholar] [CrossRef]
- Kobayashi, S.; Mochida, Y.; Ishioka, K.; Oka, M.; Maesato, K.; Moriya, H.; Hidaka, S.; Ohtake, T. The effects of blood pressure and the renin–angiotensin–aldosterone system on regional cerebral blood flow and cognitive impairment in dialysis patients. Hypertens. Res. 2014, 37, 636–641. [Google Scholar] [CrossRef]
- Pires, P.W.; McClain, J.L.; Hayoz, S.F.; Dorrance, A.M. Mineralocorticoid receptor antagonism prevents obesity-induced cerebral artery remodeling and reduces white matter injury in rats. Microcirculation 2018, 25, e12460. [Google Scholar] [CrossRef]
- Hajjar, I.; Hart, M.; Mack, W.; Lipsitz, L.A. Aldosterone, cognitive function, and cerebral hemodynamics in hypertension and antihypertensive therapy. Am. J. Hypertens. 2015, 28, 319–325. [Google Scholar] [CrossRef]
- Bukiya, A.N.; Leo, M.D.; Jaggar, J.H.; Dopico, A.M. Cholesterol activates BK channels by increasing KCNMB1 protein levels in the plasmalemma. J. Biol. Chem. 2021, 296, 100381. [Google Scholar] [CrossRef]
- Atkovska, K.; Klingler, J.; Oberwinkler, J.; Keller, S.; Hub, J.S. Rationalizing Steroid Interactions with Lipid Membranes: Conformations, Partitioning, and Kinetics. ACS Cent. Sci. 2018, 4, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Dopico, A.M.; Bukiya, A.N.; Singh, A.K. Large conductance, calcium- and voltage-gated potassium (BK) channels: Regulation by cholesterol. Pharmacol. Ther. 2012, 135, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Rushmer, R.F. Structure and Function of the Cardiovascular System. In Organ Physiology; Saunders: Philadelphia, PA, USA, 1989; pp. 5–22. [Google Scholar] [CrossRef]
- Valverde, M.A.; Rojas, P.; Amigo, J.; Cosmelli, D.; Orio, P.; Bahamonde, M.I.; Mann, G.E.; Vergara, C.; Latorre, R. Acute activation of Maxi-K channels (hSlo) by estradiol binding to the beta subunit. Science 1999, 285, 1929–1931. [Google Scholar] [CrossRef] [PubMed]
- Senisterra, G.; Chau, I.; Vedadi, M. Thermal Denaturation Assays in Chemical Biology. ASSAY Drug Dev. Technol. 2012, 10, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.E.; Funder, J.W.; Kattoula, S.R. Evolution of hormone selectivity in glucocorticoid and mineralocorticoid receptors. J. Steroid Biochem. Mol. Biol. 2013, 137, 57–70. [Google Scholar] [CrossRef] [PubMed]
- McClain, J.L.; Dorrance, A.M. Temporary mineralocorticoid receptor antagonism during the development of hypertension improves cerebral artery dilation. Exp. Biol. Med. 2014, 239, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, C.; Struthers, A. Aldosterone induces acute endothelial dysfunction in vivo in humans: Evidence for an aldosterone-induced vasculopathy. Clin. Sci. 2002, 103, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Bukiya, A.N.; McMillan, J.; Parrill, A.L.; Dopico, A.M. Structural determinants of monohydroxylated bile acids to activate beta 1 subunit-containing BK channels. J. Lipid Res. 2008, 49, 2441–2451. [Google Scholar] [CrossRef]
- Thorsen, T.; Tangen, M.; Støa, K. Concentration of endogenous oestradiol as related to oestradiol receptor sites in breast tumor cytosol. Eur. J. Cancer Clin. Oncol. 1982, 18, 333–337. [Google Scholar] [CrossRef]
- Weber, K.T. Aldosterone in Congestive Heart Failure. N. Engl. J. Med. 2001, 345, 1689–1697. [Google Scholar] [CrossRef]
- Zhang, Y.D.; Ding, X.J.; Dai, H.Y.; Peng, W.S.; Guo, N.F.; Zhang, Y.; Zhou, Q.L.; Chen, X.L. SB-216763, a GSK-3β inhibitor, protects against aldosterone-induced cardiac, and renal injury by activating autophagy. J. Cell. Biochem. 2018, 119, 5934–5943. [Google Scholar] [CrossRef]
- Dopico, A.M.; Anantharam, V.; Treistman, S.N. Ethanol increases the activity of Ca(++)-dependent K+ (mslo) channels: Functional interaction with cytosolic Ca++. J. Pharmacol. Exp. Ther. 1998, 284, 258–268. [Google Scholar]
- Liu, P.; Xi, Q.; Ahmed, A.; Jaggar, J.H.; Dopico, A.M. Essential role for smooth muscle BK channels in alcohol-induced cerebrovascular constriction. Proc. Natl. Acad. Sci. USA 2004, 101, 18217–18222. [Google Scholar] [CrossRef]
- Bukiya, A.N.; McMillan, J.E.; Fedinec, A.L.; Patil, S.A.; Miller, D.D.; Leffler, C.W.; Parrill, A.L.; Dopico, A.M. Cerebrovascular Dilation via Selective Targeting of the Cholane Steroid-Recognition Site in the BK Channel β1-Subunit by a Novel Nonsteroidal Agent. Mol. Pharmacol. 2013, 83, 1030–1044. [Google Scholar] [CrossRef] [PubMed]
- Kuntamallappanavar, G.; Bisen, S.; Bukiya, A.N.; Dopico, A.M. Differential distribution and functional impact of BK channel beta1 subunits across mesenteric, coronary, and different cerebral arteries of the rat. Pflügers Arch. -Eur. J. Physiol. 2017, 469, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Mysiewicz, S.; North, K.; Moreira, L.; Odum, S.J.; Bukiya, A.; Dopico, A.M. Interspecies and regional variability of alcohol action on large cerebral arteries; regulation by KCNMB1 proteins. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2023, 324, R480–R496. [Google Scholar] [CrossRef]
- Ahmed, A.; Waters, C.M.; Leffler, C.W.; Jaggar, J.H. Ionic mechanisms mediating the myogenic response in newborn porcine cerebral arteries. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H2061–H2069. [Google Scholar] [CrossRef] [PubMed]
- Clifford, P.S.; Ferguson, B.S.; Jasperse, J.L.; Hill, M.A. Arteriolar vasodilation involves actin depolymerization. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H423–H428. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mysiewicz, S.C.; Hawks, S.M.; Bukiya, A.N.; Dopico, A.M. Differential Functional Contribution of BK Channel Subunits to Aldosterone-Induced Channel Activation in Vascular Smooth Muscle and Eventual Cerebral Artery Dilation. Int. J. Mol. Sci. 2023, 24, 8704. https://doi.org/10.3390/ijms24108704
Mysiewicz SC, Hawks SM, Bukiya AN, Dopico AM. Differential Functional Contribution of BK Channel Subunits to Aldosterone-Induced Channel Activation in Vascular Smooth Muscle and Eventual Cerebral Artery Dilation. International Journal of Molecular Sciences. 2023; 24(10):8704. https://doi.org/10.3390/ijms24108704
Chicago/Turabian StyleMysiewicz, Steven C., Sydney M. Hawks, Anna N. Bukiya, and Alex M. Dopico. 2023. "Differential Functional Contribution of BK Channel Subunits to Aldosterone-Induced Channel Activation in Vascular Smooth Muscle and Eventual Cerebral Artery Dilation" International Journal of Molecular Sciences 24, no. 10: 8704. https://doi.org/10.3390/ijms24108704
APA StyleMysiewicz, S. C., Hawks, S. M., Bukiya, A. N., & Dopico, A. M. (2023). Differential Functional Contribution of BK Channel Subunits to Aldosterone-Induced Channel Activation in Vascular Smooth Muscle and Eventual Cerebral Artery Dilation. International Journal of Molecular Sciences, 24(10), 8704. https://doi.org/10.3390/ijms24108704