
Overexpression of Rice BSR2 Confers Disease Resistance and Induces Enlarged Flowers in Torenia fournieri Lind


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
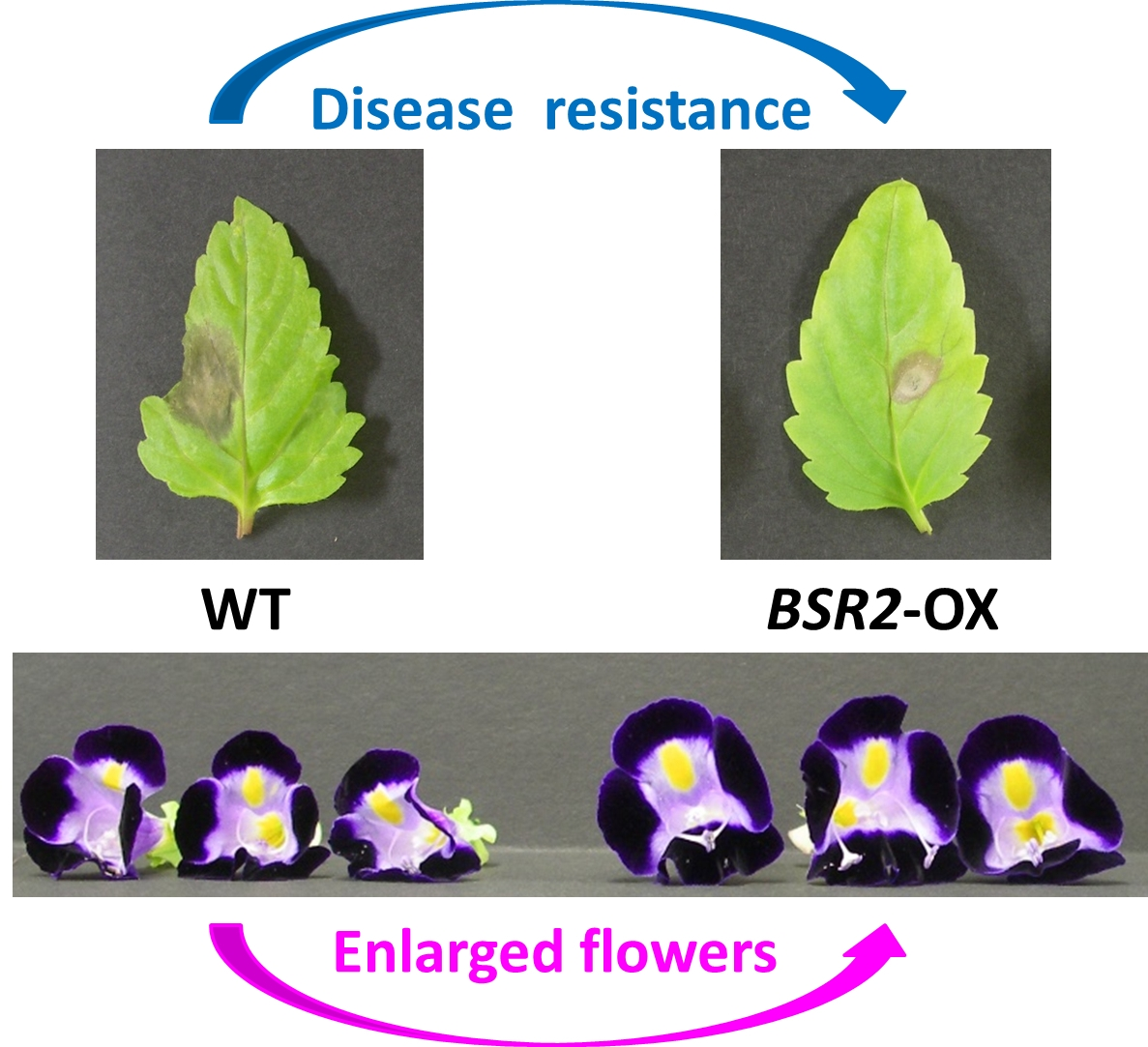
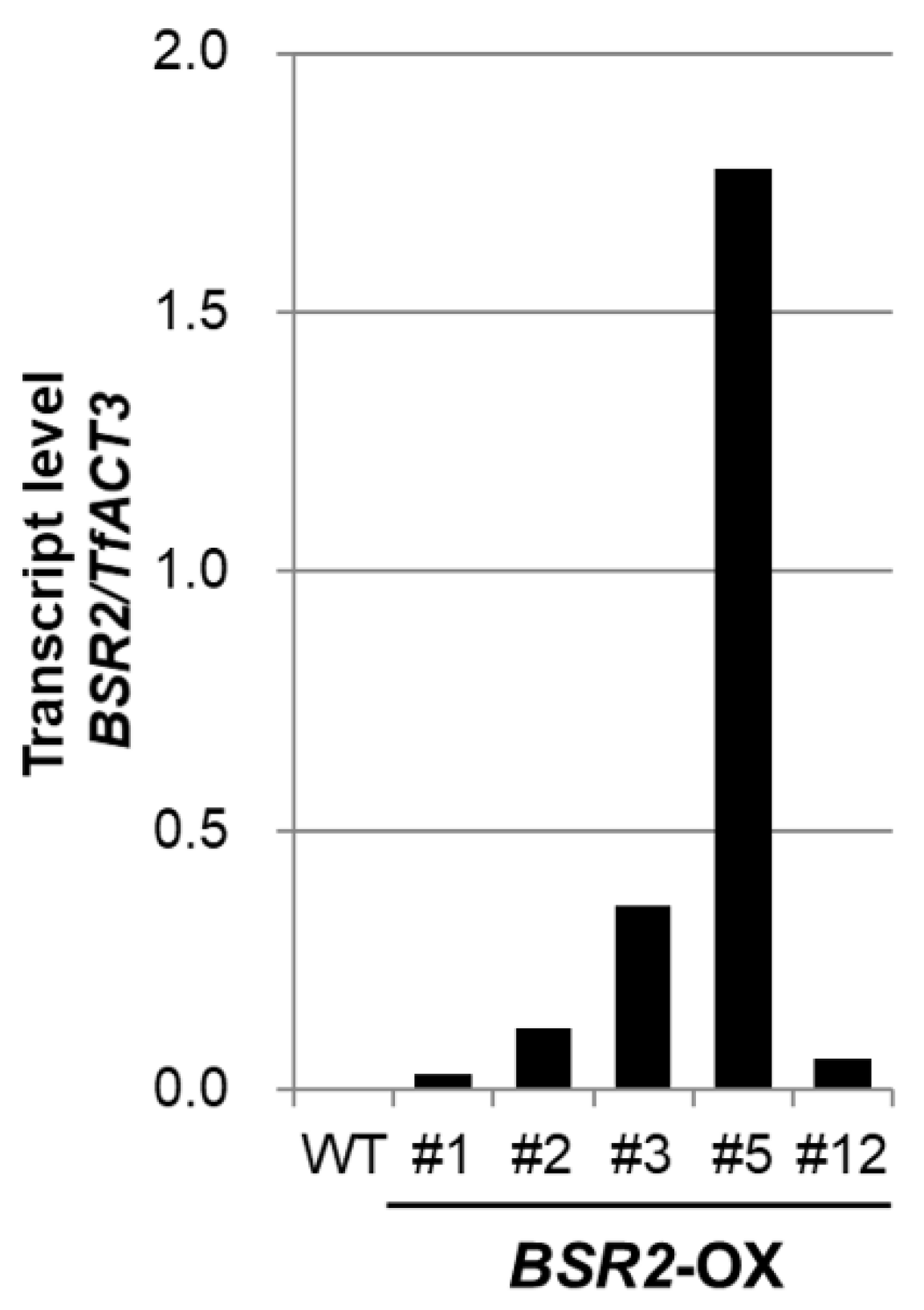
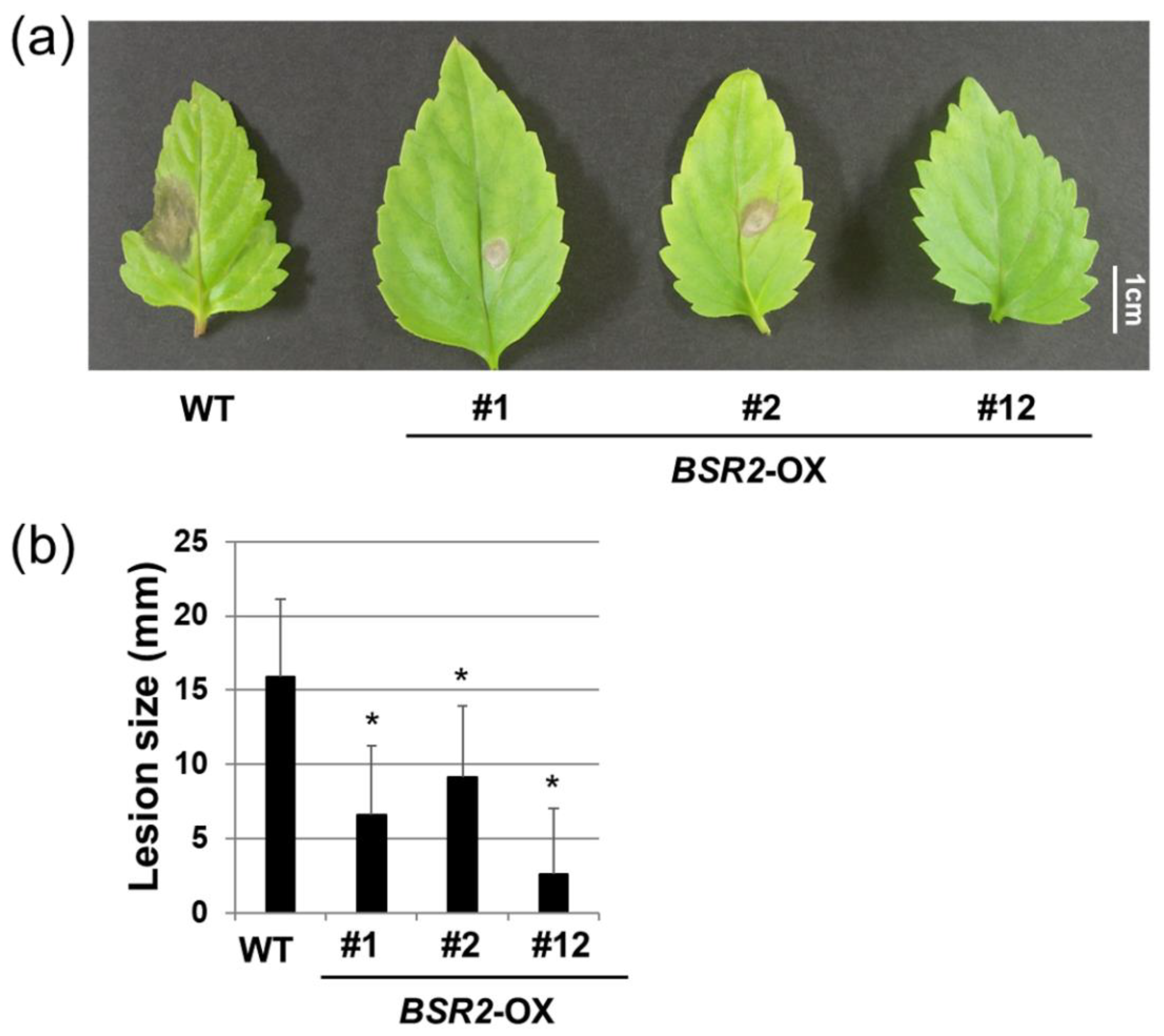
2.1. Overexpression of BSR2 Conferred Resistance against Two Fungal Pathogens in T. fournieri
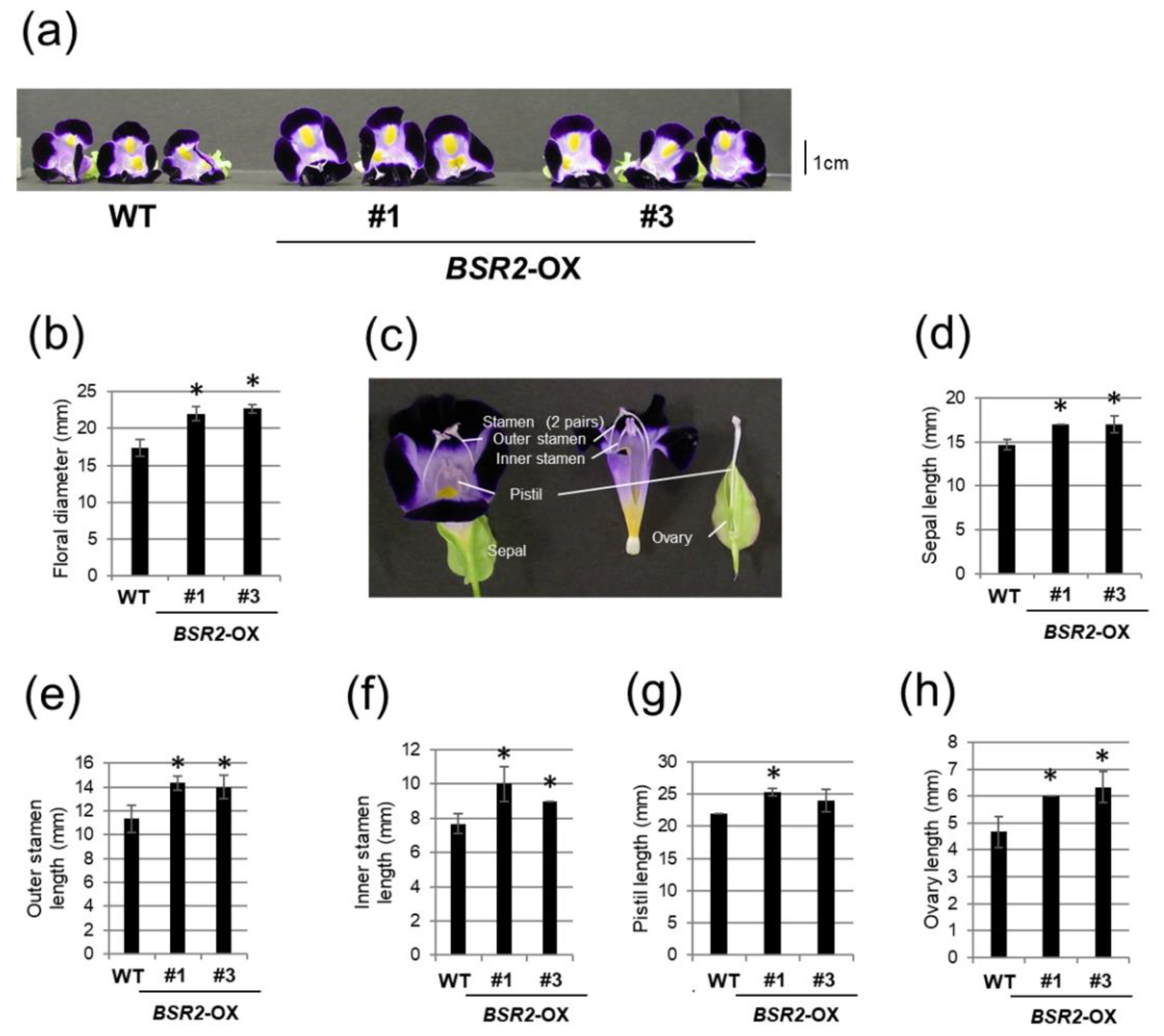
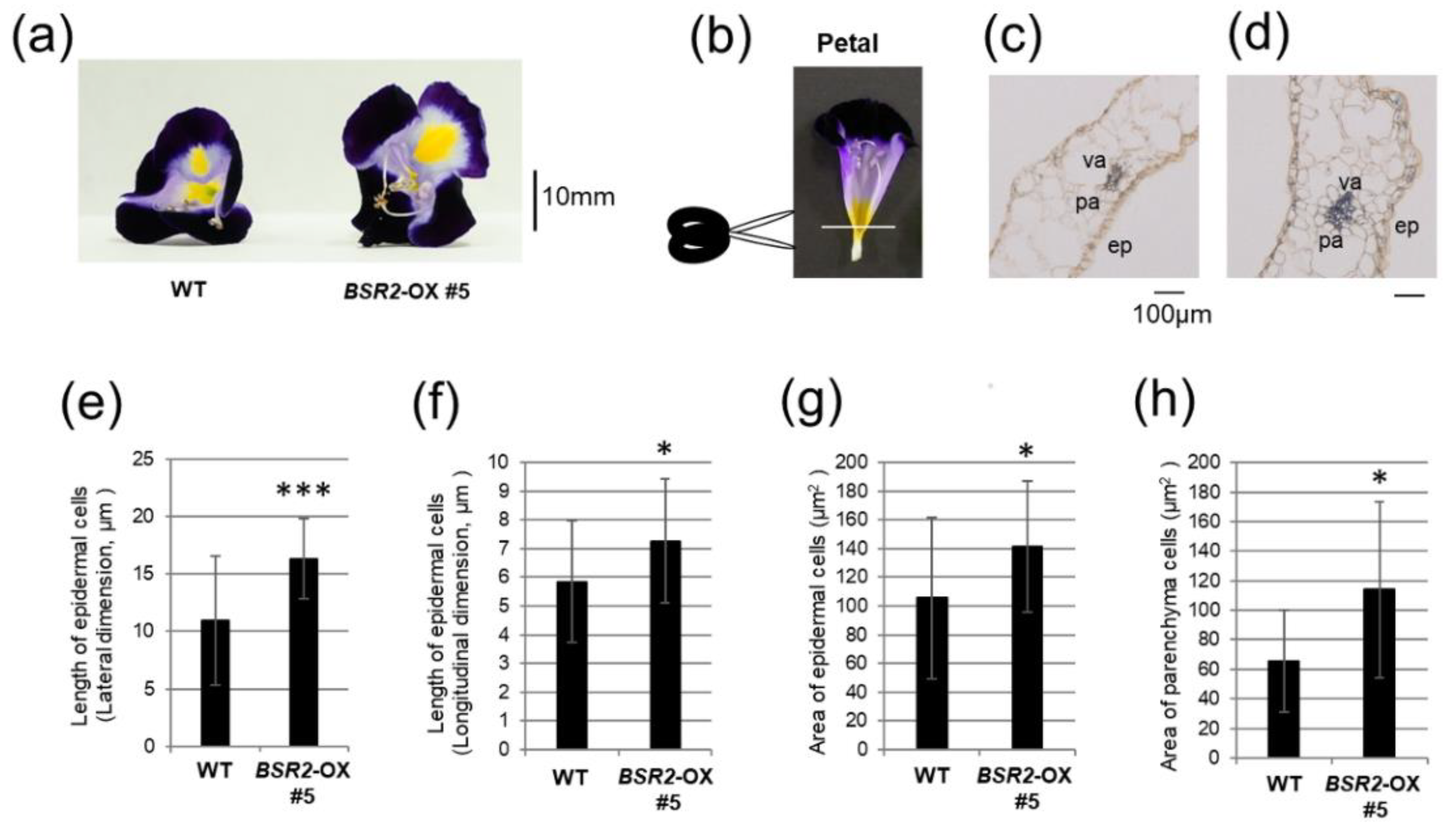
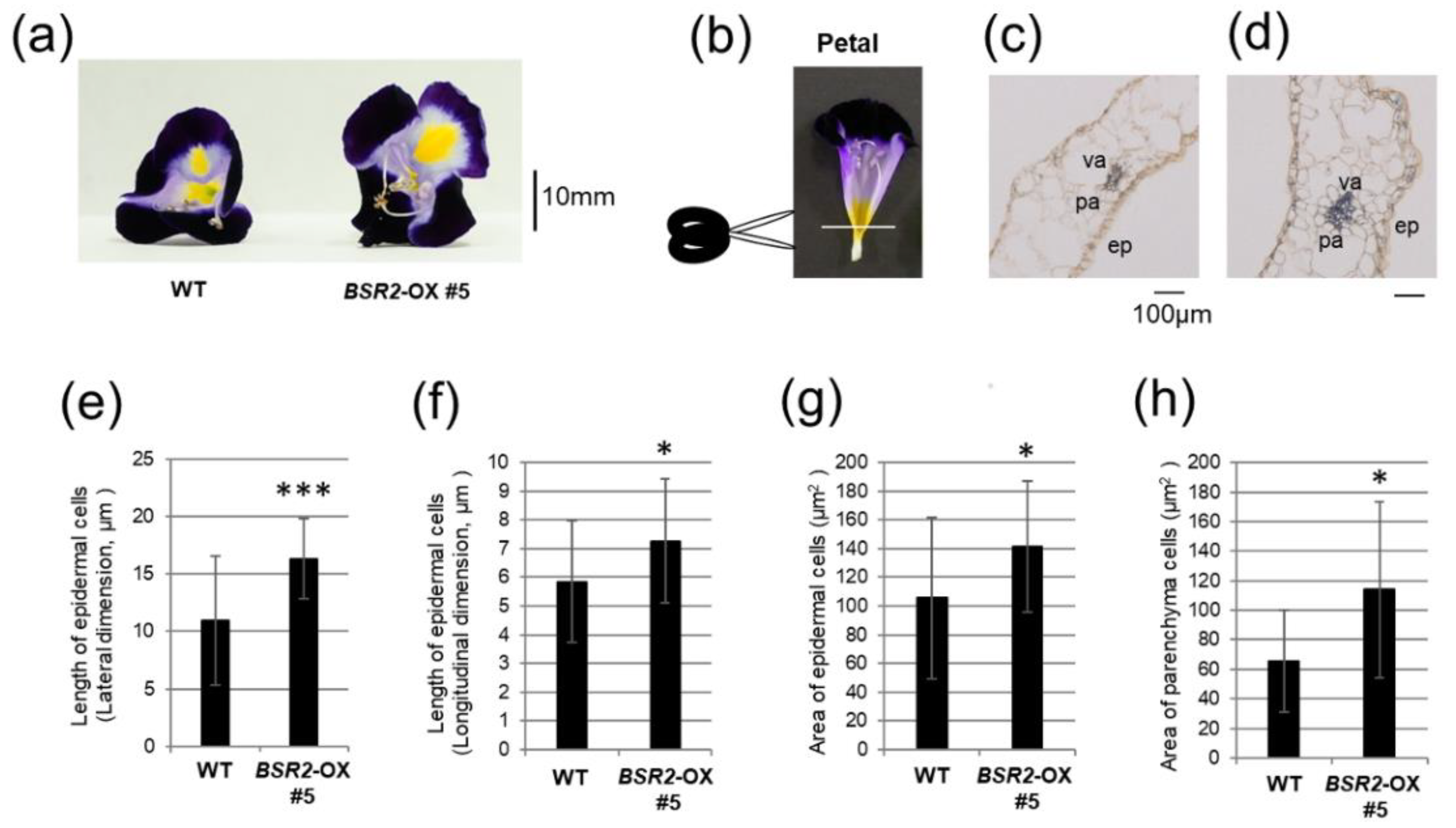
2.2. Overexpression of BSR2 Resulted in Enlarged Flowers with Enlarged Floral Organs
3. Discussion
4. Materials and Methods
4.1. Plant and Microbial Materials and Culture
4.2. T. fournieri Transformation
4.3. RNA Extraction and Quantitative Real-Time RT-PCR (qRT-PCR) Analysis
4.4. Fungal Pathogen Resistance Assay
4.5. Flow Cytometry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Molla, K.A.; Karmakar, S.; Molla, J.; Bajaj, P.; Varshney, R.K.; Datta, S.K.; Datta, K. Understanding sheath blight resistance in rice: The road behind and the road ahead. Plant Biotechnol. J. 2020, 18, 895–915. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.M.; Cartwright, D.K. Ornamental diseases incited by Rhizoctonia spp. In Rhizoctonia Species: Taxonomy, Molecular biology, Ecology, Pathology and Disease Control; Springer: Berlin/Heidelberg, Germany, 1996; pp. 303–314. [Google Scholar]
- MAEDA, S.; Dubouzet, J.G.; Kondou, Y.; Jikumaru, Y.; Seo, S.; Oda, K.; Matsui, M.; Hirochika, H.; Mori, M. The rice CYP78A gene BSR2 confers resistance to Rhizoctonia solani and affects seed size and growth in Arabidopsis and rice. Sci. Rep. 2019, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Meyerowitz, E.M. Overexpression of a gene encoding a cytochrome P450, CYP78A9, induces large and seedless fruit in Arabidopsis. Plant Cell 2000, 12, 1541–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiou, E.; Kenz, S.; Gerstung, M.; MacLean, D.; Timmer, J.; Fleck, C.; Lenhard, M. Control of plant organ size by KLUH/CYP78A5-dependent intercellular signaling. Dev. Cell 2007, 13, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Wang, Z.; Cui, R.; Li, J.; Li, Y. Maternal control of seed size by EOD3/CYP78A6 in Arabidopsis thaliana. Plant J. 2012, 70, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Yokotani, N.; Oda, K.; Mori, M. Enhanced resistance to fungal and bacterial diseases in tomato and Arabidopsis expressing BSR2 from rice. Plant Cell Rep. 2020, 39, 1493–1503. [Google Scholar] [CrossRef]
- Kikuchi, S.; Tanaka, H.; Shiba, T.; Mii, M.; Tsujimoto, H. Genome size, karyotype, meiosis and a novel extra chromosome in Torenia fournieri, T. baillonii and their hybrid. Chromosome Res. 2006, 14, 665–672. [Google Scholar] [CrossRef]
- Katsumata, T.; Fukazawa, J.; Magome, H.; Jikumaru, Y.; Kamiya, Y.; Natsume, M.; Kawaide, H.; Yamaguchi, S. Involvement of the CYP78A subfamily of cytochrome P450 monooxygenases in protonema growth and gametophore formation in the moss Physcomitrella patens. Biosci. Biotechnol. Biochem. 2011, 75, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Bak, S.; Beisson, F.; Bishop, G.; Hamberger, B.; Höfer, R.; Paquette, S.; Werck-Reichhart, D. Cytochromes p450. Arab. Book 2011, 9, e0144. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, M.; Zhang, N.; Sauvage, C.; Muños, S.; Blanca, J.; Cañizares, J.; Diez, M.J.; Schneider, R.; Mazourek, M.; McClead, J.; et al. A cytochrome P450 regulates a domestication trait in cultivated tomato. Proc. Natl. Acad. Sci. USA 2013, 110, 17125–17130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, Y.; Zhang, H.; Sun, G.; Zhang, W.; Qiu, L. Evolution and association analysis of GmCYP78A10 gene with seed size/weight and pod number in soybean. Mol. Biol. Rep. 2015, 42, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Zhao, H.; Li, Z.; Hu, S.; Song, W.; Liu, X. TaCYP78A5 regulates seed size in wheat (Triticum aestivum). J. Exp. Bot. 2016, 67, 1397–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotelo-Silveira, M.; Cucinotta, M.; Chauvin, A.L.; Chávez Montes, R.A.; Colombo, L.; Marsch-Martínez, N.; de Folter, S. Cytochrome P450 CYP78A9 is involved in Arabidopsis reproductive development. Plant Physiol. 2013, 162, 779–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaishi, H.; Matsuo, S.; Swai, E.; Ohkawa, H. CYP78A1 preferentially expressed in developing inflorescences of Zea mays encoded a cytochrome P450-dependent lauric acid 12-monooxygenase. Biosci. Biotechnol. Biochem. 2000, 64, 1696–1701. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chakrabarti, M.; Taitano, N.K.; Okazaki, Y.; Saito, K.; Al-Abdallat, A.M.; van der Knaap, E. Differential expression of SIKLUH controlling fruit and seed weight is associated with changes in lipid metabolism and photosynthesis-related genes. J. Exp. Bot. 2021, 72, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.T.; Cao, X.Q.; Shi, S.C.; Zhao, N.; Li, D.D.; Fang, P.H.; Chen, X.; Qi, W.C.; Zhang, Z. Comparative RNA-Seq analysis reveals a critical role for brassinosteroids in rose (Rosa hybrida) petal defense against Botrytis cinerea infection. BMC Genet. 2018, 19, 62. [Google Scholar] [CrossRef]
- Sasaki, K.; Yamaguchi, H.; Kasajima, I.; Narumi, T.; Ohtsubo, N. Generation of Novel Floral Traits Using a Combination of Floral Organ-Specific Promoters and a Chimeric Repressor in Torenia fournieri Lind. Plant Cell Physiol. 2016, 57, 1319–1331. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Sasaki, K.; Shikata, M.; Aida, R.; Ohtsubo, N. Trehalose drastically extends the in vitro vegetative culture period and facilitates maintenance of Torenia fournieri plants. Plant Biotechnol. 2011, 28, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Aida, R.; Shibata, M. Agrobacterium-Mediated Transformation of Torenia (Torenia fournieri). Breed. Sci. 1995, 45, 71–74. [Google Scholar] [CrossRef]
- Aida, R. A protocol for transformation of Torenia. Methods Mol. Biol. 2012, 847, 267–274. [Google Scholar] [PubMed]
- Mishiba, K.; Ando, T.; Mii, M.; Watanabe, H.; Kokubun, H.; Hashimoto, G.; Marchesi, E. Nuclear DNA content as an index character discriminating taxa in the genus Petunia sensu Jussieu (Solanaceae). Ann. Bot. 2000, 85, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Shimizu, A.; Degi, K.; Morishita, T. Effects of dose and dose rate of gamma ray irradiation on mutation induction and nuclear DNA content in chrysanthemum. Breed. Sci. 2008, 58, 331–335. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, S.; Sasaki, K.; Kaku, H.; Kanda, Y.; Ohtsubo, N.; Mori, M. Overexpression of Rice BSR2 Confers Disease Resistance and Induces Enlarged Flowers in Torenia fournieri Lind. Int. J. Mol. Sci. 2022, 23, 4735. https://doi.org/10.3390/ijms23094735
Maeda S, Sasaki K, Kaku H, Kanda Y, Ohtsubo N, Mori M. Overexpression of Rice BSR2 Confers Disease Resistance and Induces Enlarged Flowers in Torenia fournieri Lind. International Journal of Molecular Sciences. 2022; 23(9):4735. https://doi.org/10.3390/ijms23094735
Chicago/Turabian StyleMaeda, Satoru, Katsutomo Sasaki, Hisatoshi Kaku, Yasukazu Kanda, Norihiro Ohtsubo, and Masaki Mori. 2022. "Overexpression of Rice BSR2 Confers Disease Resistance and Induces Enlarged Flowers in Torenia fournieri Lind" International Journal of Molecular Sciences 23, no. 9: 4735. https://doi.org/10.3390/ijms23094735
APA StyleMaeda, S., Sasaki, K., Kaku, H., Kanda, Y., Ohtsubo, N., & Mori, M. (2022). Overexpression of Rice BSR2 Confers Disease Resistance and Induces Enlarged Flowers in Torenia fournieri Lind. International Journal of Molecular Sciences, 23(9), 4735. https://doi.org/10.3390/ijms23094735