
Protein Phosphatase 2A Improves Cardiac Functional Response to Ischemia and Sepsis

 , and
, and 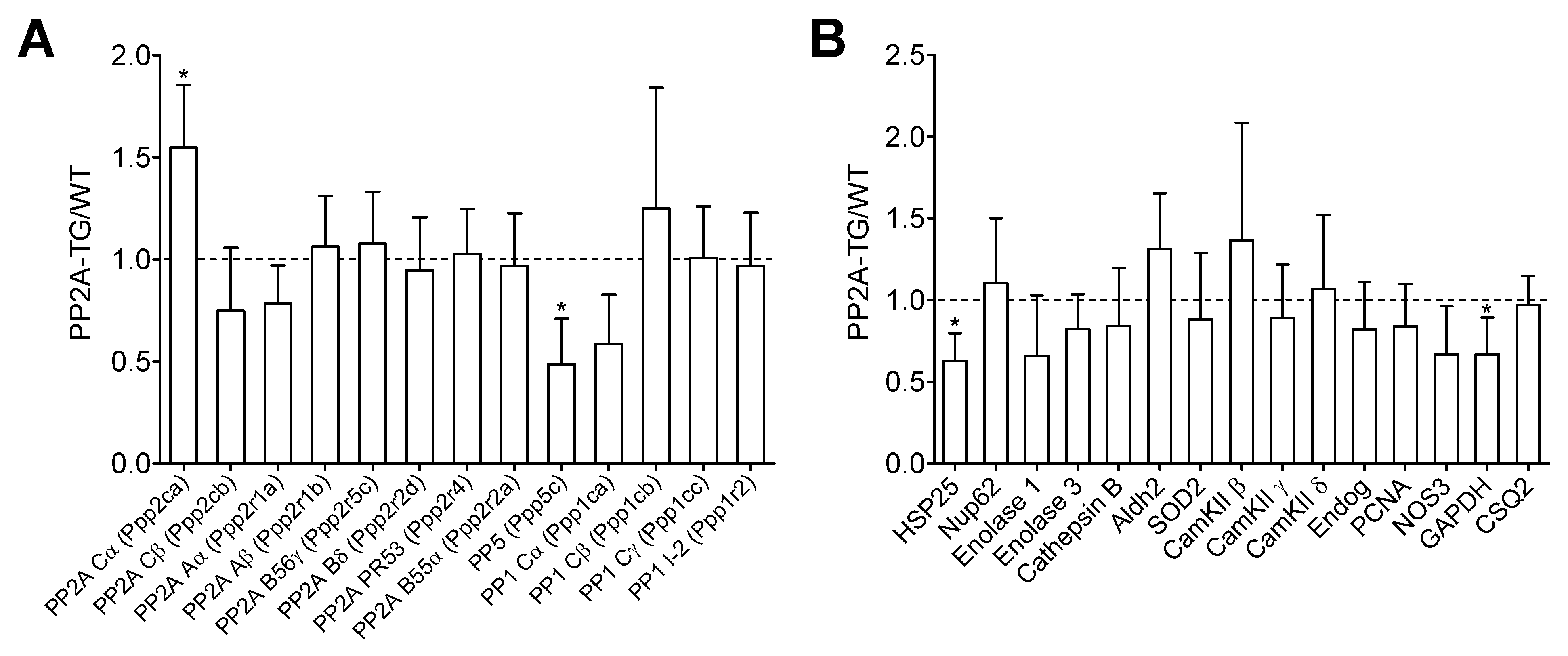{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Differential Gene Expression under Basal Conditions in PP2A-TG Hearts
2.2. Role of PP2A in Sepsis
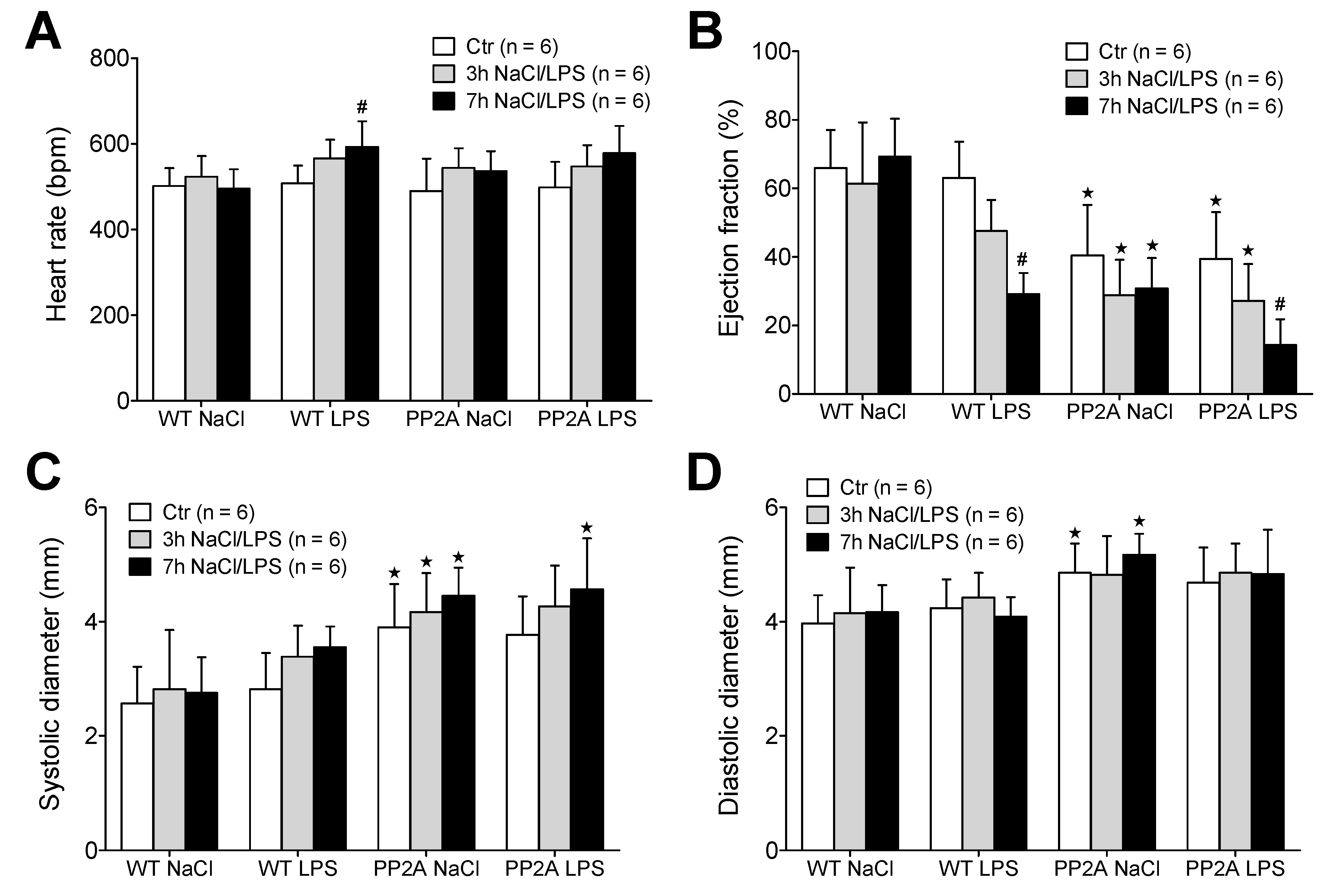
2.2.1. LPS and Echocardiography
2.2.2. LPS and Working Heart
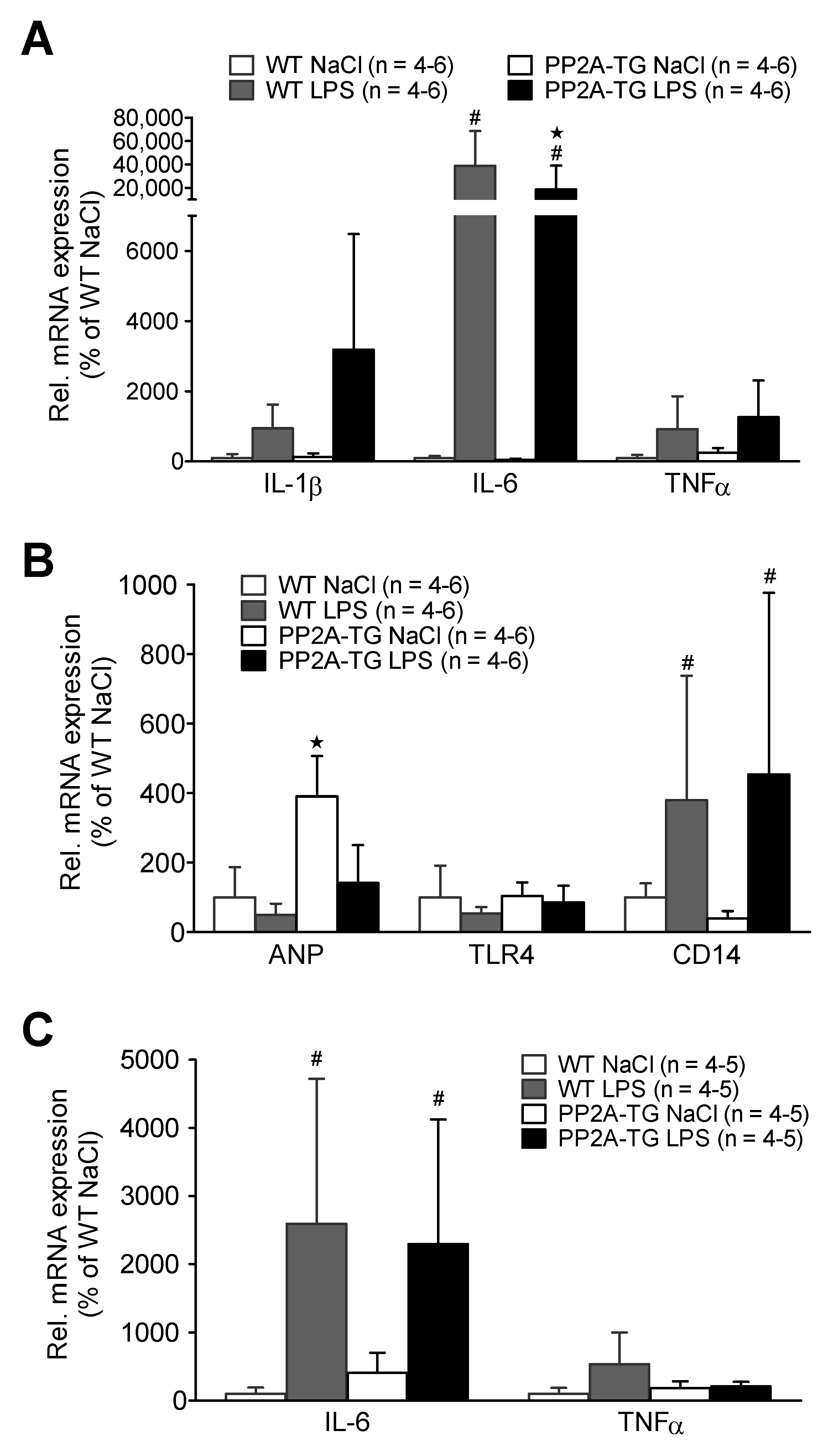
2.2.3. LPS and Gene Expression
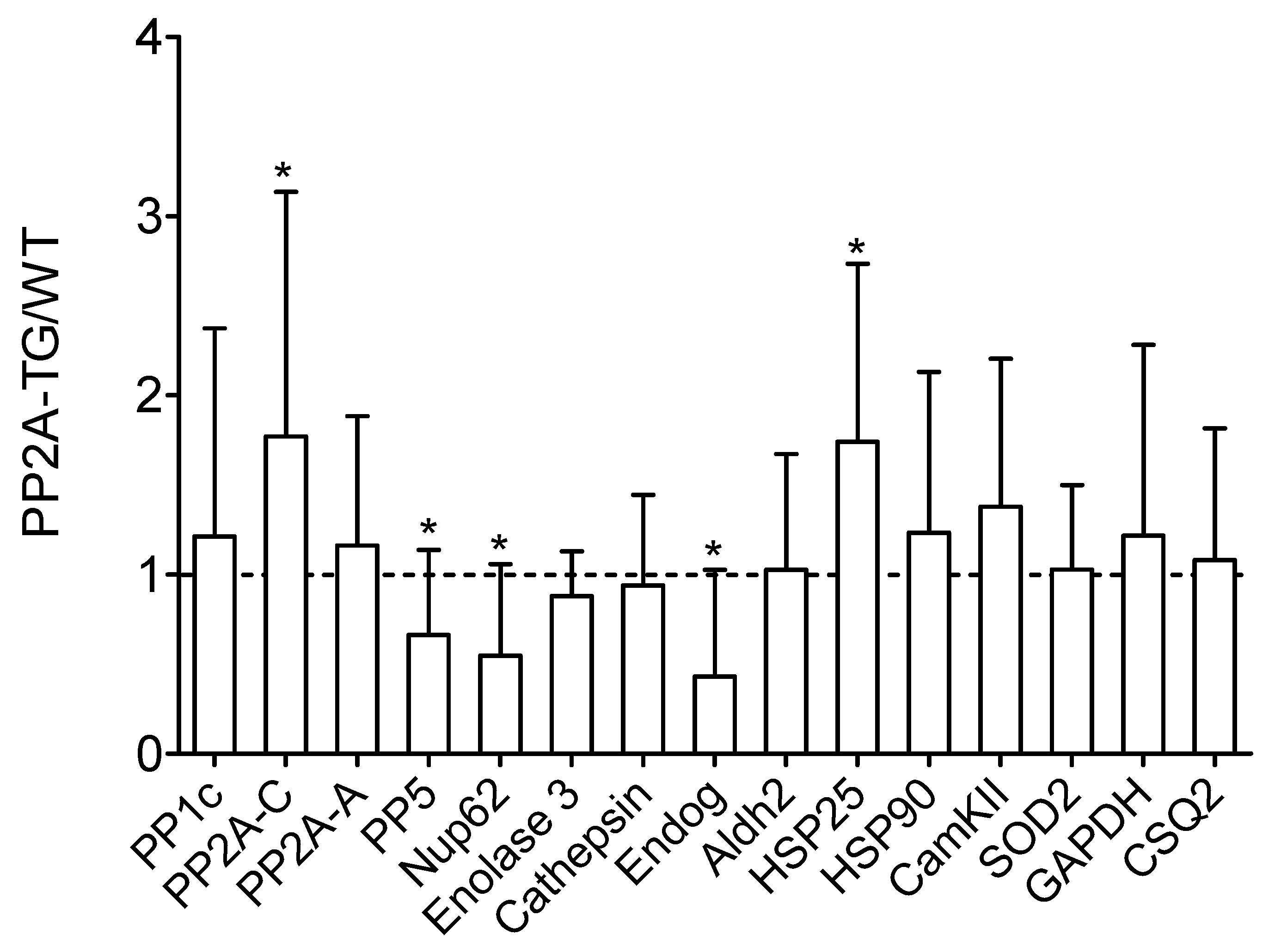
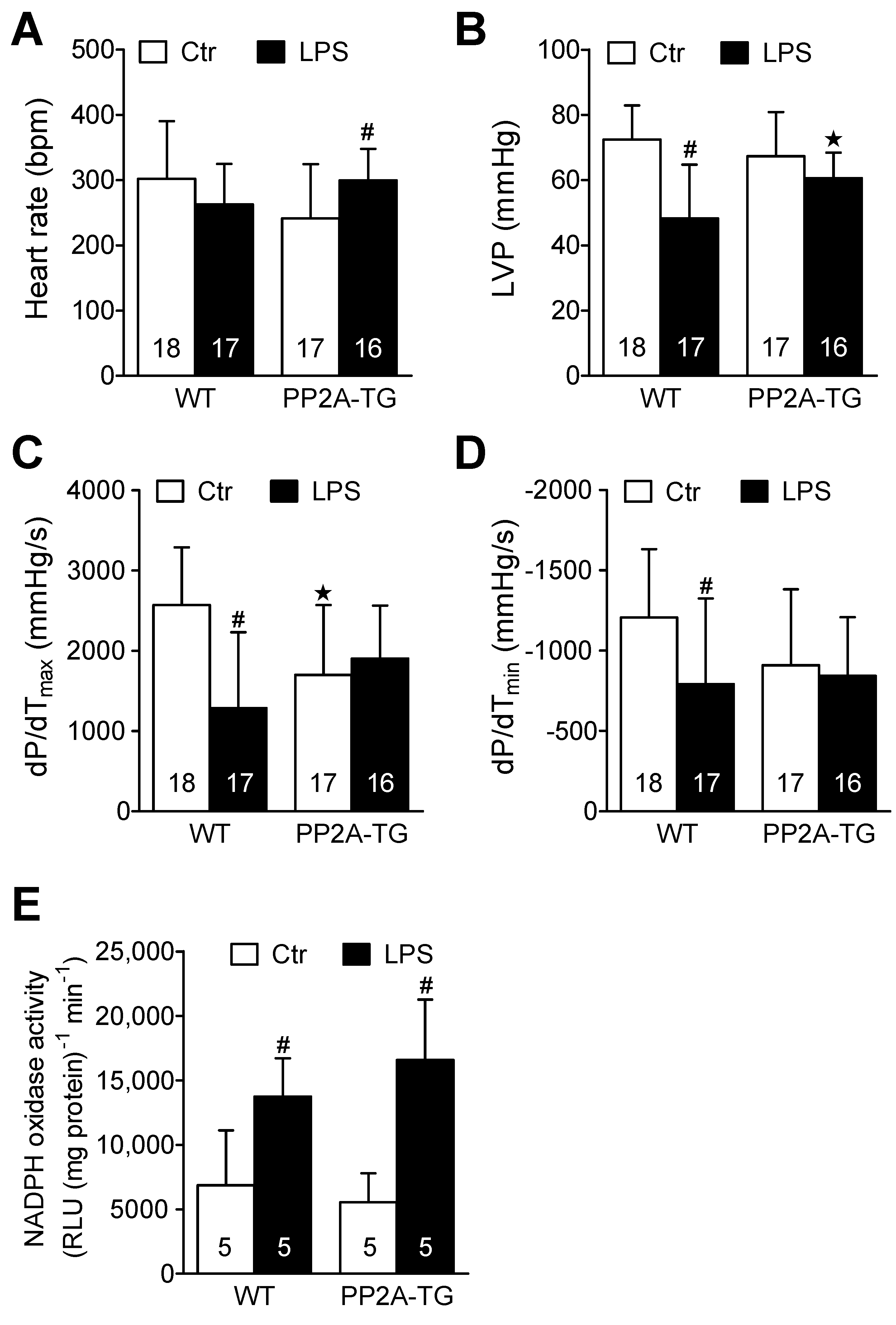
2.3. Role of PP2A in Ischemia/Reperfusion
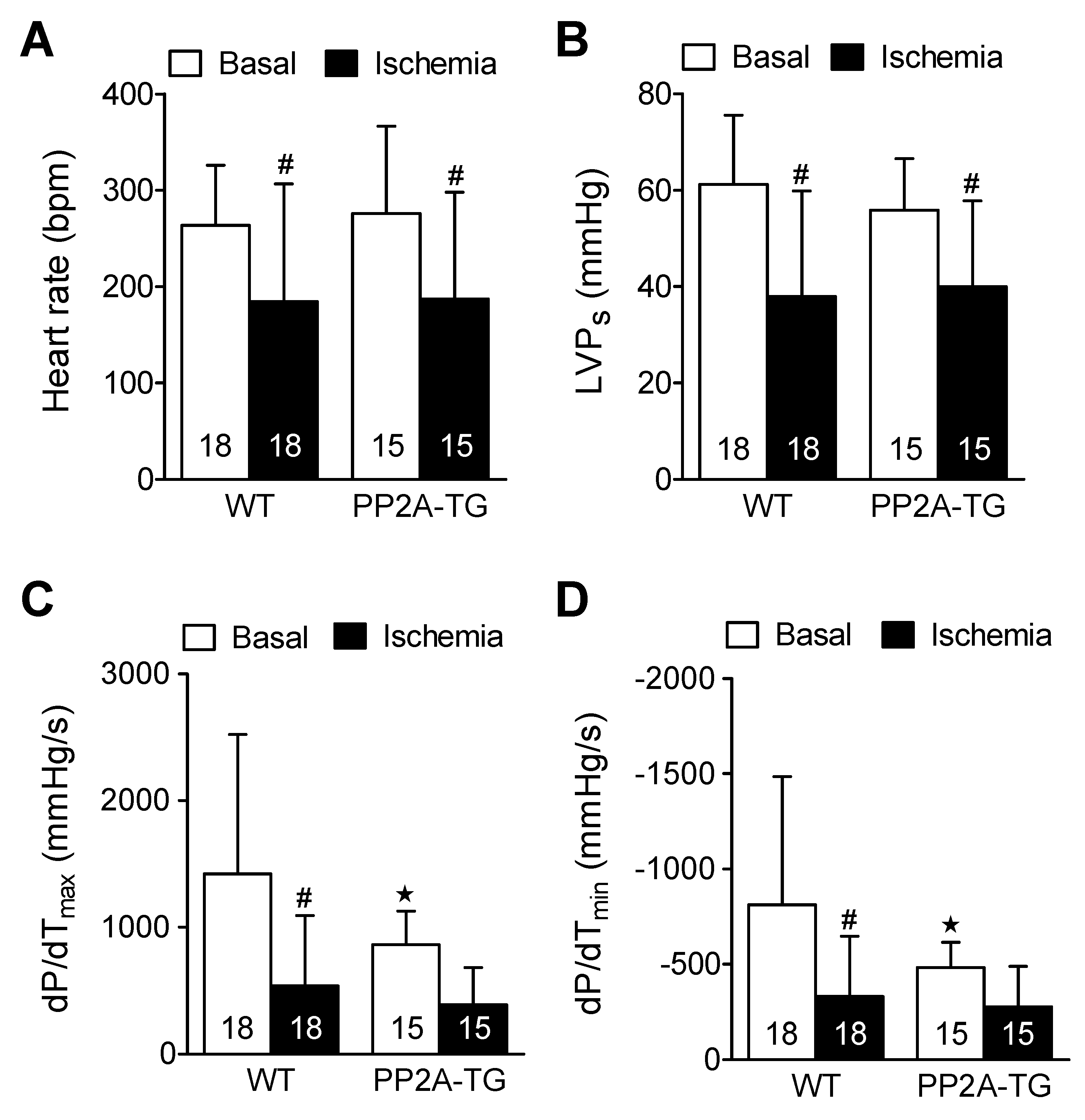
2.3.1. Cardiac Function In Vitro
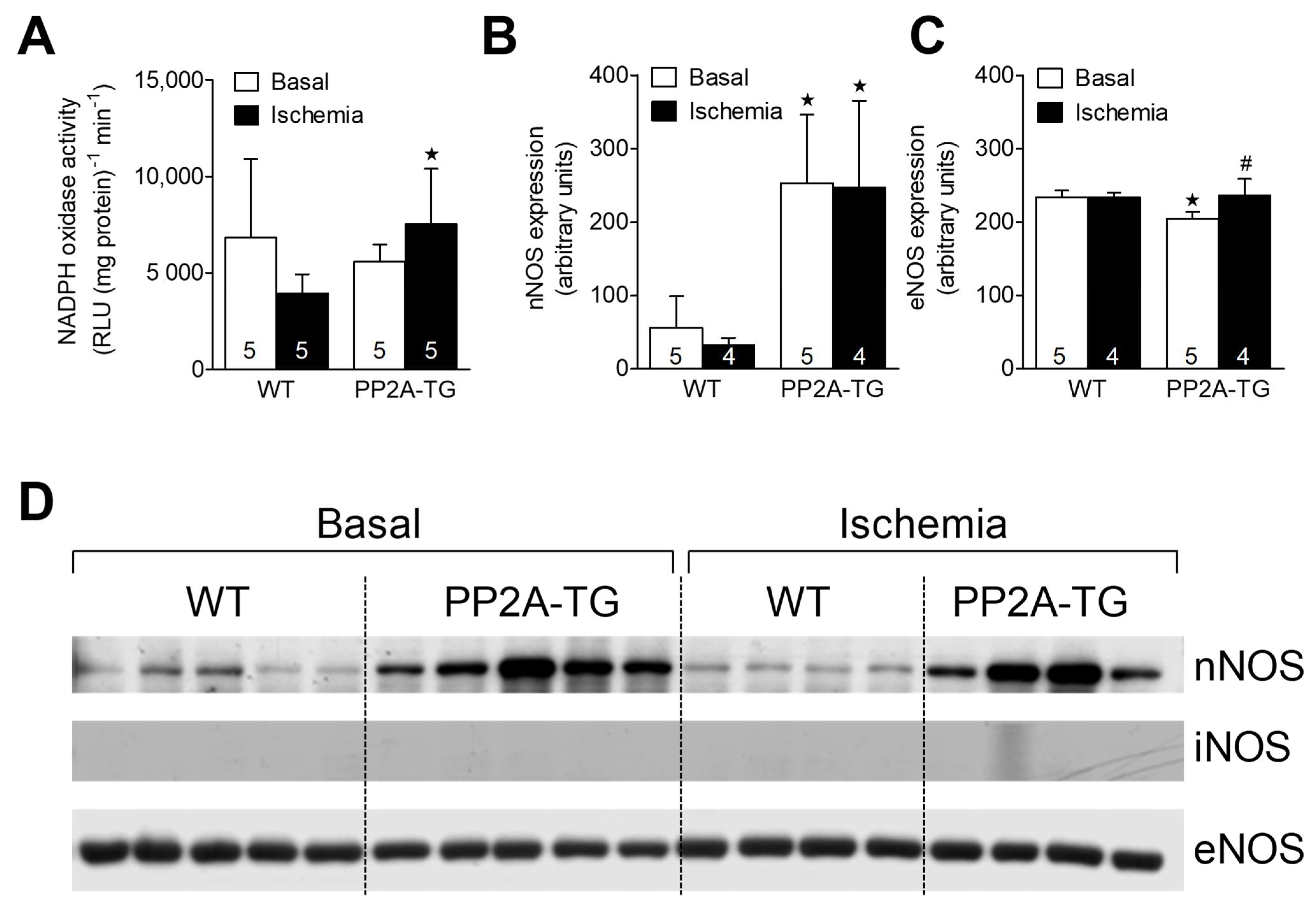
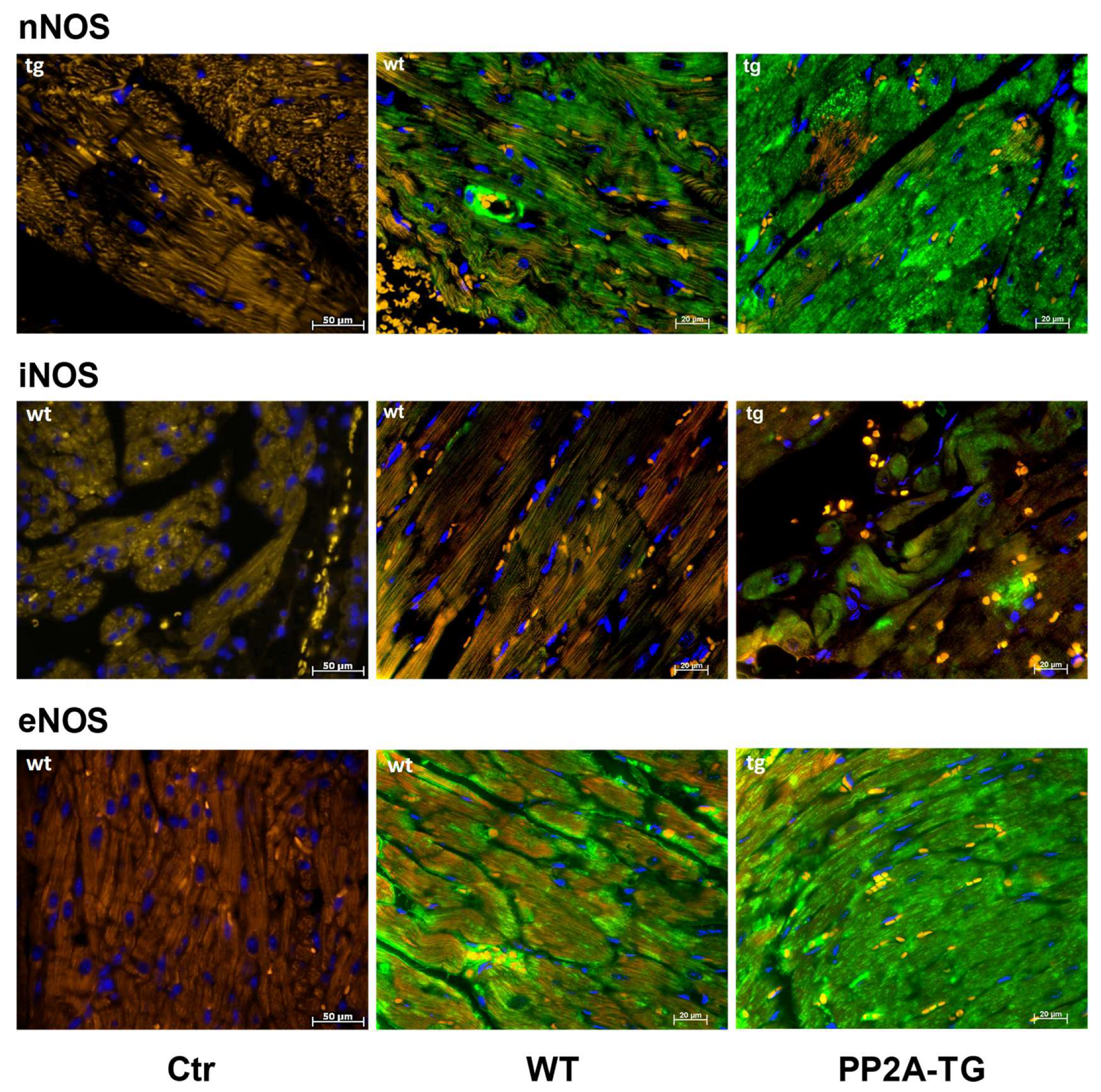
2.3.2. NADPH Oxidase and Nitric Oxide Synthases
3. Discussion
4. Materials and Methods
4.1. Transgenic Mice
4.2. RNA Extraction and cDNA Synthesis
4.3. Gene Chip Analysis
4.4. Real Time PCR
4.5. Western Blot Analysis
4.6. Echocardiography
4.7. Work-Performing Heart Preparations
4.8. NADPH Oxidase Activity
4.9. Immunohistochemistry
4.10. Reagents
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heijman, J.; Dewenter, M.; El-Armouche, A.; Dobrev, D. Function and regulation of serine/threonine phosphatases in the healthy and diseased heart. J. Mol. Cell. Cardiol. 2013, 64, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, E.R.; Mohler, P.J. Roles and regulation of protein phosphatase 2A (PP2A) in the heart. J. Mol. Cell. Cardiol. 2016, 101, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Reynhout, S.; Janssens, V. Physiologic functions of PP2A: Lessons from genetically modified mice. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Sergienko, N.M.; Donner, D.G.; Delbridge, L.M.D.; McMullen, J.R.; Weeks, K.L. Protein phosphatase 2A in the healthy and failing heart: New insights and therapeutic opportunities. Cell. Signal. 2022, 91, 110213. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- L’Heureux, M.; Sternberg, M.; Brath, L.; Turlington, J.; Kashiouris, M.G. Sepsis-Induced Cardiomyopathy: A Comprehensive Review. Curr. Cardiol. Rep. 2020, 22, 35. [Google Scholar] [CrossRef]
- Wasyluk, W.; Nowicka-Stążka, P.; Zwolak, A. Heart Metabolism in Sepsis-Induced Cardiomyopathy-Unusual Metabolic Dysfunction of the Heart. Int. J. Environ. Res. Public Health 2021, 18, 7598. [Google Scholar] [CrossRef]
- Annane, D.; Bellissant, E.; Cavaillon, J.-M. Septic shock. Lancet 2005, 365, 63–78. [Google Scholar] [CrossRef]
- Page, M.J.; Kell, D.B.; Pretorius, E. The Role of Lipopolysaccharide-Induced Cell Signalling in Chronic Inflammation. Chronic Stress (Thousand Oaks) 2022, 6, 24705470221076390. [Google Scholar] [CrossRef]
- Clark, A.R.; Ohlmeyer, M. Protein phosphatase 2A as a therapeutic target in inflammation and neurodegeneration. Pharmacol. Ther. 2019, 201, 181–201. [Google Scholar] [CrossRef]
- Gergs, U.; Boknik, P.; Buchwalow, I.; Fabritz, L.; Matus, M.; Justus, I.; Hanske, G.; Schmitz, W.; Neumann, J. Overexpression of the catalytic subunit of protein phosphatase 2A impairs cardiac function. J. Biol. Chem. 2004, 279, 40827–40834. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Li, L.; Gu, P.; Jin, T.; Wen, M.; Yuan, C.; Gao, X.; Liu, C.; Zhang, Z. Profiling metabolic remodeling in PP2Acα deficiency and chronic pressure overload mouse hearts. FEBS Lett. 2015, 589, 3631–3639. [Google Scholar] [CrossRef] [PubMed]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion--from mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Elgenaidi, I.S.; Spiers, J.P. Hypoxia modulates protein phosphatase 2A through HIF-1α dependent and independent mechanisms in human aortic smooth muscle cells and ventricular cardiomyocytes. Br. J. Pharmacol. 2019, 176, 1745–1763. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-S.; Jin, H.; Sun, X.; Huang, S.; Zhang, F.-L.; Guo, Z.-N.; Yang, Y. Free Radical Damage in Ischemia-Reperfusion Injury: An Obstacle in Acute Ischemic Stroke after Revascularization Therapy. Oxid. Med. Cell. Longev. 2018, 2018, 3804979. [Google Scholar] [CrossRef] [PubMed]
- Elgenaidi, I.S.; Spiers, J.P. Regulation of the phosphoprotein phosphatase 2A system and its modulation during oxidative stress: A potential therapeutic target? Pharmacol. Ther. 2019, 198, 68–89. [Google Scholar] [CrossRef] [PubMed]
- Piran, S.; Liu, P.; Morales, A.; Hershberger, R.E. Where genome meets phenome: Rationale for integrating genetic and protein biomarkers in the diagnosis and management of dilated cardiomyopathy and heart failure. J. Am. Coll. Cardiol. 2012, 60, 283–289. [Google Scholar] [CrossRef]
- Gergs, U.; Jahn, T.; Werner, F.; Köhler, C.; Köpp, F.; Großmann, C.; Neumann, J. Overexpression of protein phosphatase 5 in the mouse heart: Reduced contractility but increased stress tolerance—Two sides of the same coin? PLoS ONE 2019, 14, e0221289. [Google Scholar] [CrossRef]
- Li, S.; Wang, L.; Berman, M.A.; Zhang, Y.; Dorf, M.E. RNAi screen in mouse astrocytes identifies phosphatases that regulate NF-kappaB signaling. Mol. Cell 2006, 24, 497–509. [Google Scholar] [CrossRef]
- Sun, L.; Pham, T.T.; Cornell, T.T.; McDonough, K.L.; McHugh, W.M.; Blatt, N.B.; Dahmer, M.K.; Shanley, T.P. Myeloid-Specific Gene Deletion of Protein Phosphatase 2A Magnifies MyD88- and TRIF-Dependent Inflammation following Endotoxin Challenge. J. Immunol. 2017, 198, 404–416. [Google Scholar] [CrossRef]
- Shanley, T.P.; Vasi, N.; Denenberg, A.; Wong, H.R. The serine/threonine phosphatase, PP2A: Endogenous regulator of inflammatory cell signaling. J. Immunol. 2001, 166, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Sipido, K.R.; Stankovicova, T.; Flameng, W.; Vanhaecke, J.; Verdonck, F. Frequency dependence of Ca2+ release from the sarcoplasmic reticulum in human ventricular myocytes from end-stage heart failure. Cardiovasc. Res. 1998, 37, 478–488. [Google Scholar] [CrossRef]
- Kirchhefer, U.; Neumann, J.; Bers, D.; Buchwalow, I.; Fabritz, L.; Hanske, G.; Justus, I.; Riemann, B.; Schmitz, W.; Jones, L. Impaired relaxation in transgenic mice overexpressing junctin. Cardiovasc. Res. 2003, 59, 369–379. [Google Scholar] [CrossRef][Green Version]
- Hoehn, M.; Zhang, Y.; Xu, J.; Gergs, U.; Boknik, P.; Werdan, K.; Neumann, J.; Ebelt, H. Overexpression of protein phosphatase 2A in a murine model of chronic myocardial infarction leads to increased adverse remodeling but restores the regulation of β-catenin by glycogen synthase kinase 3β. Int. J. Cardiol. 2015, 183, 39–46. [Google Scholar] [CrossRef]
- Loyer, X.; Heymes, C.; Samuel, J.-L. Constitutive nitric oxide synthases in the heart from hypertrophy to failure. Clin. Exp. Pharmacol. Physiol. 2008, 35, 483–488. [Google Scholar] [CrossRef]
- Neumann, J.; Eschenhagen, T.; Jones, L.R.; Linck, B.; Schmitz, W.; Scholz, H.; Zimmermann, N. Increased expression of cardiac phosphatases in patients with end-stage heart failure. J. Mol. Cell. Cardiol. 1997, 29, 265–272. [Google Scholar] [CrossRef]
- Dörner, M.-F.; Boknik, P.; Köpp, F.; Buchwalow, I.B.; Neumann, J.; Gergs, U. Mechanisms of Systolic Cardiac Dysfunction in PP2A, PP5 and PP2AxPP5 Double Transgenic Mice. Int. J. Mol. Sci. 2021, 22, 9448. [Google Scholar] [CrossRef]
- Mishra, S.; Dunkerly-Eyring, B.L.; Keceli, G.; Ranek, M.J. Phosphorylation Modifications Regulating Cardiac Protein Quality Control Mechanisms. Front. Physiol. 2020, 11, 593585. [Google Scholar] [CrossRef]
- Yoon, S.; Kook, T.; Min, H.-K.; Kwon, D.-H.; Cho, Y.K.; Kim, M.; Shin, S.; Joung, H.; Jeong, S.H.; Lee, S.; et al. PP2A negatively regulates the hypertrophic response by dephosphorylating HDAC2 S394 in the heart. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef]
- Baumann, S.; Gergs, U.; Schulz, N.; Thiery, J.; Neumann, J. Magnetic Bead-based Proteome Profiling Using MALDI-TOF Spectrometry in Cardiac Tissue from Transgenic Mice. Curr. Proteom. 2015, 11, 229–236. [Google Scholar] [CrossRef]
- Hanumanthu, B.K.J.; Nair, A.S.; Katamreddy, A.; Gilbert, J.S.; You, J.Y.; Offor, O.L.; Kushwaha, A.; Krishnan, A.; Napolitano, M.; Palaidimos, L.; et al. Sepsis-induced cardiomyopathy is associated with higher mortality rates in patients with sepsis. Acute Crit. Care 2021, 36, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Gergs, U.; Gerigk, T.; Wittschier, J.; Schmidbaur, C.T.; Röttger, C.; Mahnkopf, M.; Edler, H.; Wache, H.; Neumann, J. Influence of Serotonin 5-HT4 Receptors on Responses to Cardiac Stressors in Transgenic Mouse Models. Biomedicines 2021, 9, 569. [Google Scholar] [CrossRef] [PubMed]
- Fallach, R.; Shainberg, A.; Avlas, O.; Fainblut, M.; Chepurko, Y.; Porat, E.; Hochhauser, E. Cardiomyocyte Toll-like receptor 4 is involved in heart dysfunction following septic shock or myocardial ischemia. J. Mol. Cell. Cardiol. 2010, 48, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Hattori, K.; Suzuki, T.; Matsuda, N. Recent advances in the pathophysiology and molecular basis of sepsis-associated organ dysfunction: Novel therapeutic implications and challenges. Pharmacol. Ther. 2017, 177, 56–66. [Google Scholar] [CrossRef]
- Marshall, M.; Anilkumar, N.; Layland, J.; Walker, S.J.; Kentish, J.C.; Shah, A.M.; Cave, A.C. Protein phosphatase 2A contributes to the cardiac dysfunction induced by endotoxemia. Cardiovasc. Res. 2009, 82, 67–76. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Osaki, K.; Kanamoto, M.; Nakao, Y.; Takahashi, E.; Higuchi, T.; Kamata, H. Distinct B subunits of PP2A regulate the NF-κB signalling pathway through dephosphorylation of IKKβ, IκBα and RelA. FEBS Lett. 2017, 591, 4083–4094. [Google Scholar] [CrossRef]
- Xiong, Y.; Murphy, M.; Manavalan, T.T.; Pattabiraman, G.; Qiu, F.; Chang, H.-H.; Ho, I.-C.; Medvedev, A.E. Endotoxin Tolerance Inhibits Lyn and c-Src Phosphorylation and Association with Toll-Like Receptor 4 but Increases Expression and Activity of Protein Phosphatases. J. Innate Immun. 2016, 8, 171–184. [Google Scholar] [CrossRef]
- Gergs, U.; Kirchhefer, U.; Bergmann, F.; Künstler, B.; Mißlinger, N.; Au, B.; Mahnkopf, M.; Wache, H.; Neumann, J. Characterization of Stressed Transgenic Mice Overexpressing H2-Histamine Receptors in the Heart. J. Pharmacol. Exp. Ther. 2020, 374, 479–488. [Google Scholar] [CrossRef]
- Boknik, P.; Drzewiecki, K.; Eskandar, J.; Gergs, U.; Hofmann, B.; Treede, H.; Grote-Wessels, S.; Fabritz, L.; Kirchhof, P.; Fortmüller, L.; et al. Evidence for Arrhythmogenic Effects of A2A-Adenosine Receptors. Front. Pharmacol. 2019, 10, 1051. [Google Scholar] [CrossRef]
- Boknik, P.; Eskandar, J.; Hofmann, B.; Zimmermann, N.; Neumann, J.; Gergs, U. Role of Cardiac A2A Receptors Under Normal and Pathophysiological Conditions. Front. Pharmacol. 2020, 11, 627838. [Google Scholar] [CrossRef]
- Gündel, D.; Lai, T.H.; Dukic-Stefanovic, S.; Teodoro, R.; Deuther-Conrad, W.; Toussaint, M.; Kopka, K.; Moldovan, R.-P.; Boknik, P.; Hofmann, B.; et al. Non-Invasive Assessment of Locally Overexpressed Human Adenosine 2A Receptors in the Heart of Transgenic Mice. Int. J. Mol. Sci. 2022, 23, 1025. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, P.; Werner, F.; Jaron, M.; Bruns, T.A.; Wache, H.; Runte, J.; Boknik, P.; Kirchhefer, U.; Müller, F.U.; Buchwalow, I.B.; et al. Initial Characterization of Stressed Transgenic Mice With Cardiomyocyte-Specific Overexpression of Protein Phosphatase 2C. Front. Pharmacol. 2020, 11, 591773. [Google Scholar] [CrossRef] [PubMed]
- Nan, J.; Li, J.; Lin, Y.; Saif Ur Rahman, M.; Li, Z.; Zhu, L. The interplay between mitochondria and store-operated Ca2+ entry: Emerging insights into cardiac diseases. J. Cell. Mol. Med. 2021, 25, 9496–9512. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.; Boknik, P.; Kirchhefer, U.; Gergs, U. The role of PP5 and PP2C in cardiac health and disease. Cell. Signal. 2021, 85, 110035. [Google Scholar] [CrossRef] [PubMed]
- Weinbrenner, C.; Baines, C.P.; Liu, G.S.; Armstrong, S.C.; Ganote, C.E.; Walsh, A.H.; Honkanen, R.E.; Cohen, M.V.; Downey, J.M. Fostriecin, an inhibitor of protein phosphatase 2A, limits myocardial infarct size even when administered after onset of ischemia. Circulation 1998, 98, 899–905. [Google Scholar] [CrossRef]
- Trost, B.M.; Knopf, J.D.; Brindle, C.S. Synthetic Strategies Employed for the Construction of Fostriecin and Related Natural Products. Chem. Rev. 2016, 116, 15035–15088. [Google Scholar] [CrossRef] [PubMed]
- Reina-Couto, M.; Pereira-Terra, P.; Quelhas-Santos, J.; Silva-Pereira, C.; Albino-Teixeira, A.; Sousa, T. Inflammation in Human Heart Failure: Major Mediators and Therapeutic Targets. Front. Physiol. 2021, 12, 746494. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Steenbergen, C. Gender-based differences in mechanisms of protection in myocardial ischemia-reperfusion injury. Cardiovasc. Res. 2007, 75, 478–486. [Google Scholar] [CrossRef]
- Dawson, D.; Lygate, C.A.; Zhang, M.-H.; Hulbert, K.; Neubauer, S.; Casadei, B. nNOS Gene Deletion Exacerbates Pathological Left Ventricular Remodeling and Functional Deterioration After Myocardial Infarction. Circulation 2005, 112, 3729–3737. [Google Scholar] [CrossRef]
- Pellegrino, D.; Shiva, S.; Angelone, T.; Gladwin, M.T.; Tota, B. Nitrite exerts potent negative inotropy in the isolated heart via eNOS-independent nitric oxide generation and cGMP-PKG pathway activation. Biochim. Biophys. Acta 2009, 1787, 818–827. [Google Scholar] [CrossRef][Green Version]
- Dries, E.; Santiago, D.J.; Johnson, D.M.; Gilbert, G.; Holemans, P.; Korte, S.M.; Roderick, H.L.; Sipido, K.R. Calcium/calmodulin-dependent kinase II and nitric oxide synthase 1-dependent modulation of ryanodine receptors during β-adrenergic stimulation is restricted to the dyadic cleft. J. Physiol. 2016, 594, 5923–5939. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Neumann, J. Effects of serine/threonine protein phosphatases on ion channels in excitable membranes. Physiol. Rev. 2000, 80, 173–210. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Takahashi, M.; Shunmugavel, A.; Wallenborn, J.G.; DePaoli-Roach, A.A.; Gergs, U.; Neumann, J.; Kuppuswamy, D.; Menick, D.R.; Cooper, G. Basis for MAP4 dephosphorylation-related microtubule network densification in pressure overload cardiac hypertrophy. J. Biol. Chem. 2010, 285, 38125–38140. [Google Scholar] [CrossRef]
- Wijnker, P.J.M.; Boknik, P.; Gergs, U.; Müller, F.U.; Neumann, J.; dos Remedios, C.; Schmitz, W.; Sindermann, J.R.; Stienen, G.J.M.; van der Velden, J.; et al. Protein phosphatase 2A affects myofilament contractility in non-failing but not in failing human myocardium. J. Muscle Res. Cell Motil. 2011, 32, 221–233. [Google Scholar] [CrossRef]
- Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011; ISBN 9780309154000.
- Junho, C.V.C.; Panico, K.; Nakama, K.K.; Sonoda, M.T.; Christoffolete, M.A.; Beserra, S.S.; Roman-Campos, D.; Carneiro-Ramos, M.S. Time Course of Gene Expression Profile in Renal Ischemia and Reperfusion Injury in Mice. Transplant. Proc. 2020, 52, 2970–2976. [Google Scholar] [CrossRef]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gergs, U.; Fahrion, C.M.; Bock, P.; Fischer, M.; Wache, H.; Hauptmann, S.; Schmitz, W.; Neumann, J. Evidence for a functional role of calsequestrin 2 in mouse atrium. Acta Physiol. 2017, 219, 669–682. [Google Scholar] [CrossRef]
- Rueckschloss, U.; Villmow, M.; Klöckner, U. NADPH oxidase-derived superoxide impairs calcium transients and contraction in aged murine ventricular myocytes. Exp. Gerontol. 2010, 45, 788–796. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gergs, U.; Jahn, T.; Schulz, N.; Großmann, C.; Rueckschloss, U.; Demus, U.; Buchwalow, I.B.; Neumann, J. Protein Phosphatase 2A Improves Cardiac Functional Response to Ischemia and Sepsis. Int. J. Mol. Sci. 2022, 23, 4688. https://doi.org/10.3390/ijms23094688
Gergs U, Jahn T, Schulz N, Großmann C, Rueckschloss U, Demus U, Buchwalow IB, Neumann J. Protein Phosphatase 2A Improves Cardiac Functional Response to Ischemia and Sepsis. International Journal of Molecular Sciences. 2022; 23(9):4688. https://doi.org/10.3390/ijms23094688
Chicago/Turabian StyleGergs, Ulrich, Tina Jahn, Nico Schulz, Claudia Großmann, Uwe Rueckschloss, Uta Demus, Igor B. Buchwalow, and Joachim Neumann. 2022. "Protein Phosphatase 2A Improves Cardiac Functional Response to Ischemia and Sepsis" International Journal of Molecular Sciences 23, no. 9: 4688. https://doi.org/10.3390/ijms23094688
APA StyleGergs, U., Jahn, T., Schulz, N., Großmann, C., Rueckschloss, U., Demus, U., Buchwalow, I. B., & Neumann, J. (2022). Protein Phosphatase 2A Improves Cardiac Functional Response to Ischemia and Sepsis. International Journal of Molecular Sciences, 23(9), 4688. https://doi.org/10.3390/ijms23094688