
Monitoring Apricot (Prunus armeniaca L.) Ripening Progression through Candidate Gene Expression Analysis

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
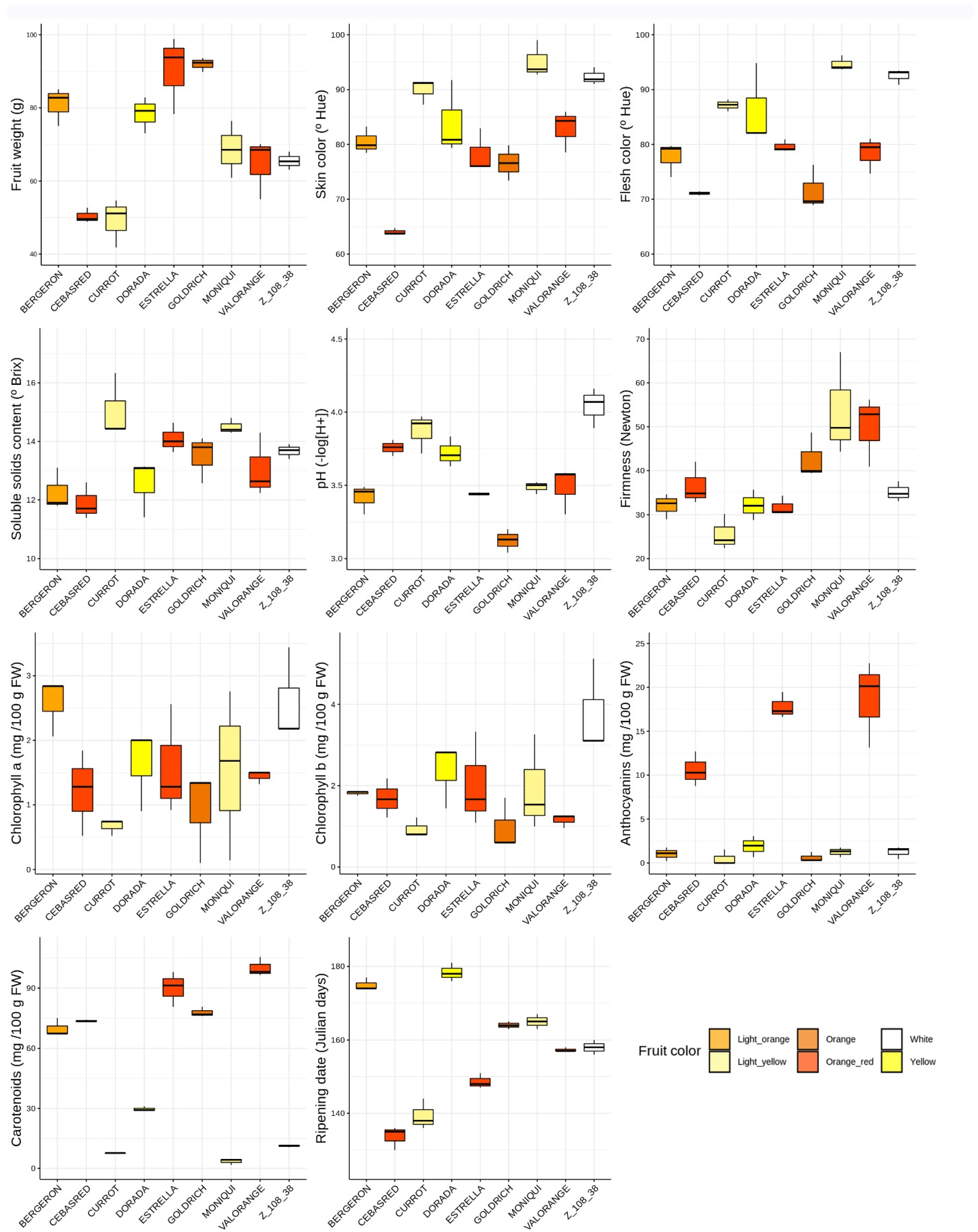
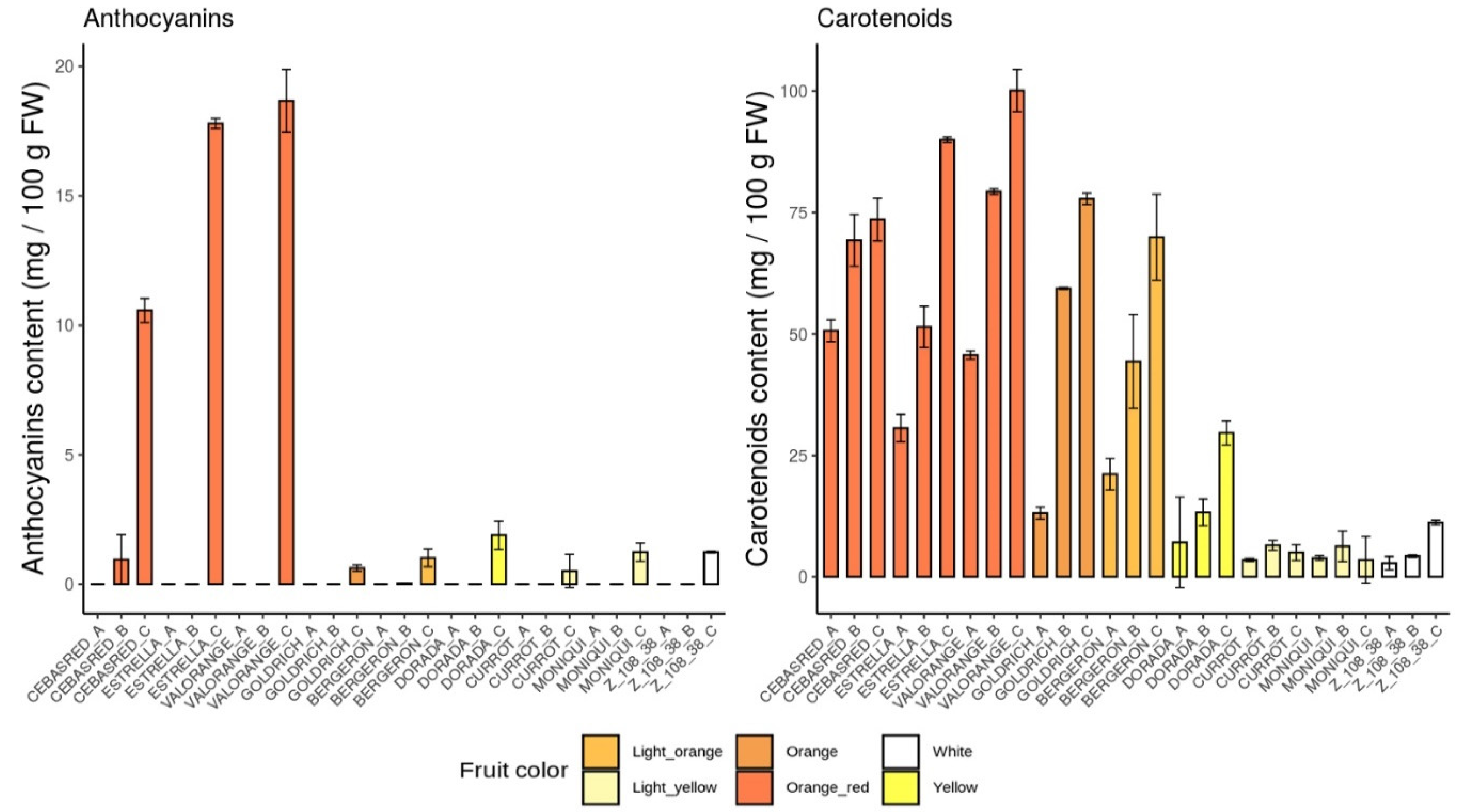
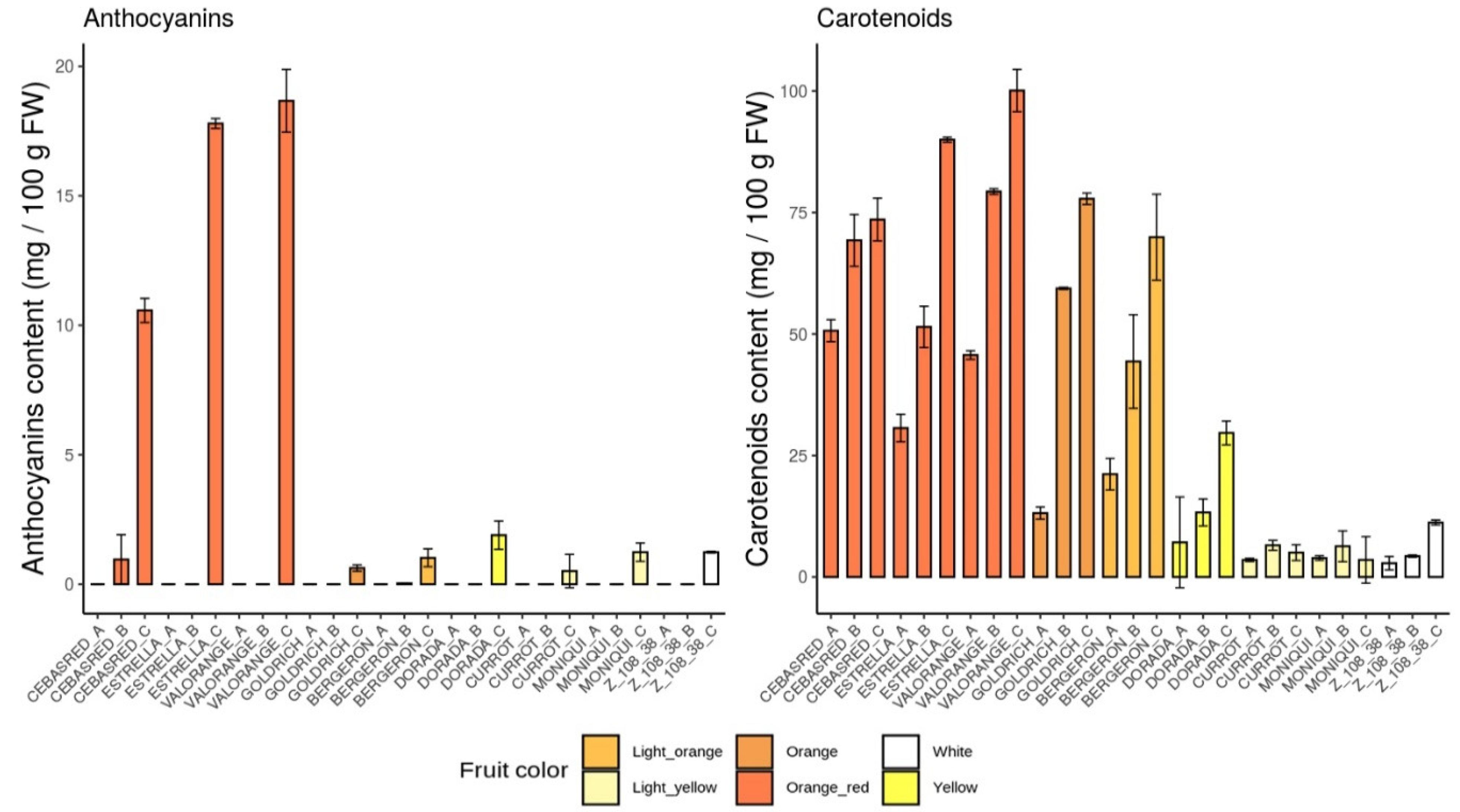
2.1. Phenotypic Evaluation
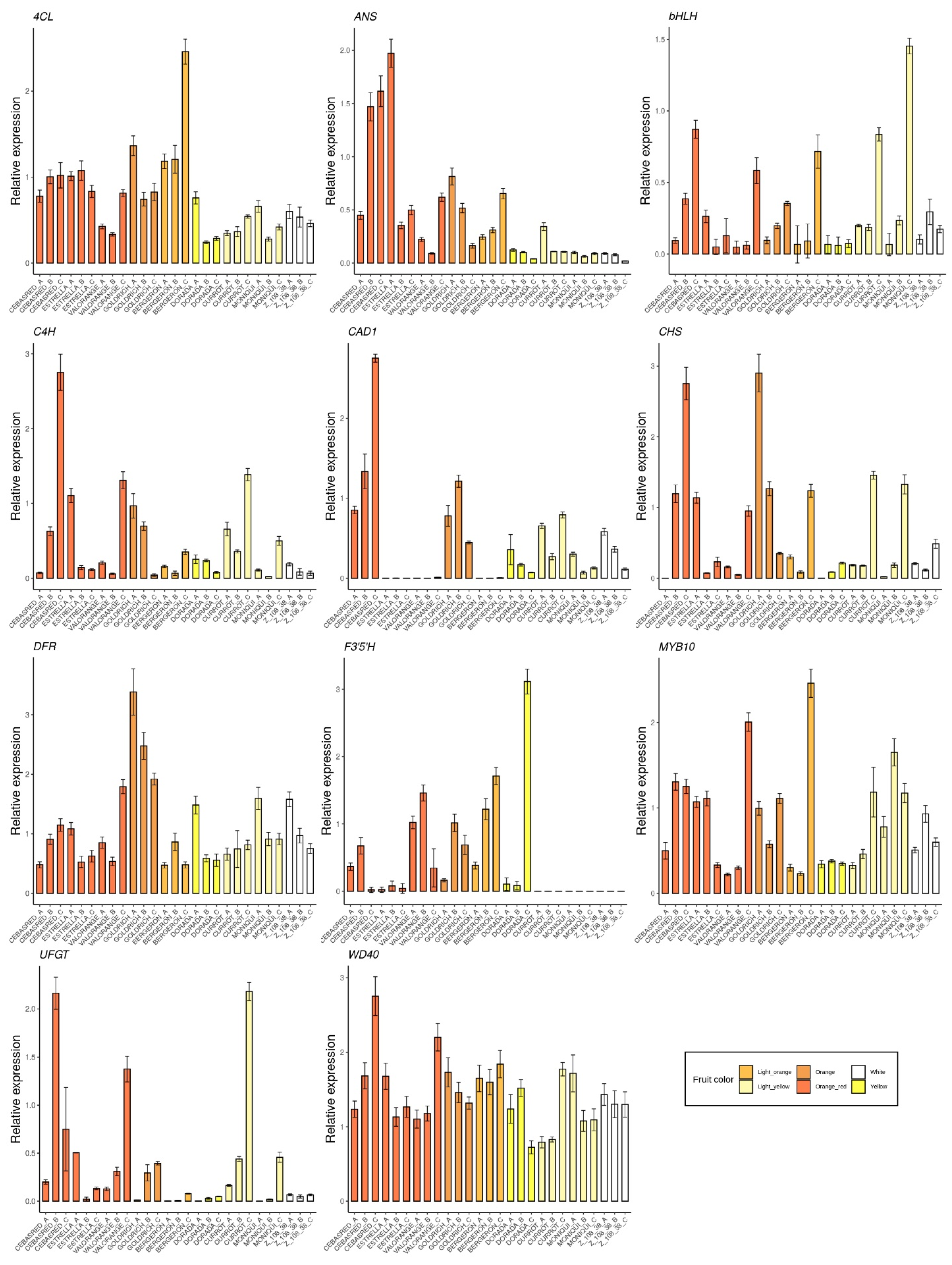
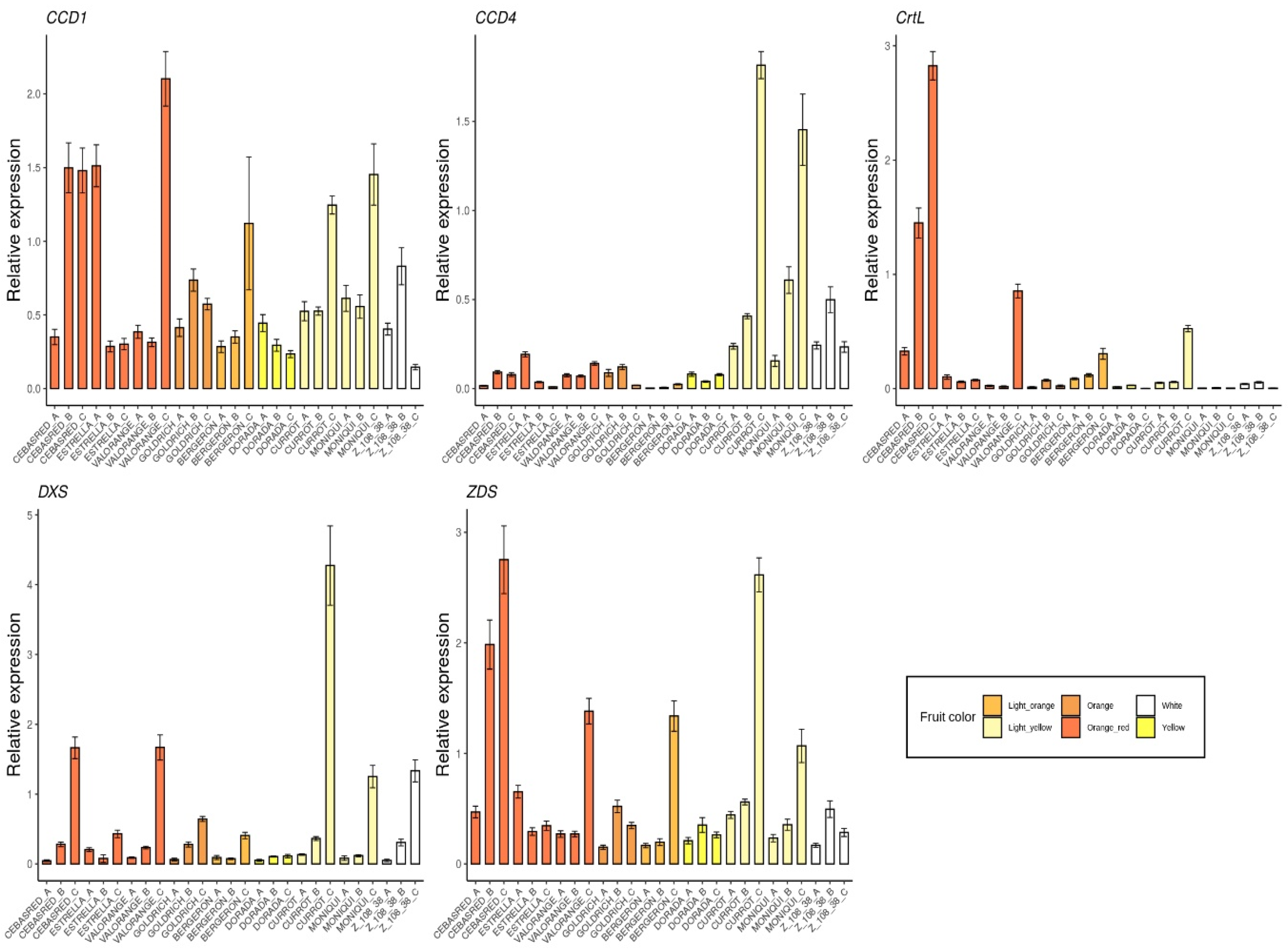
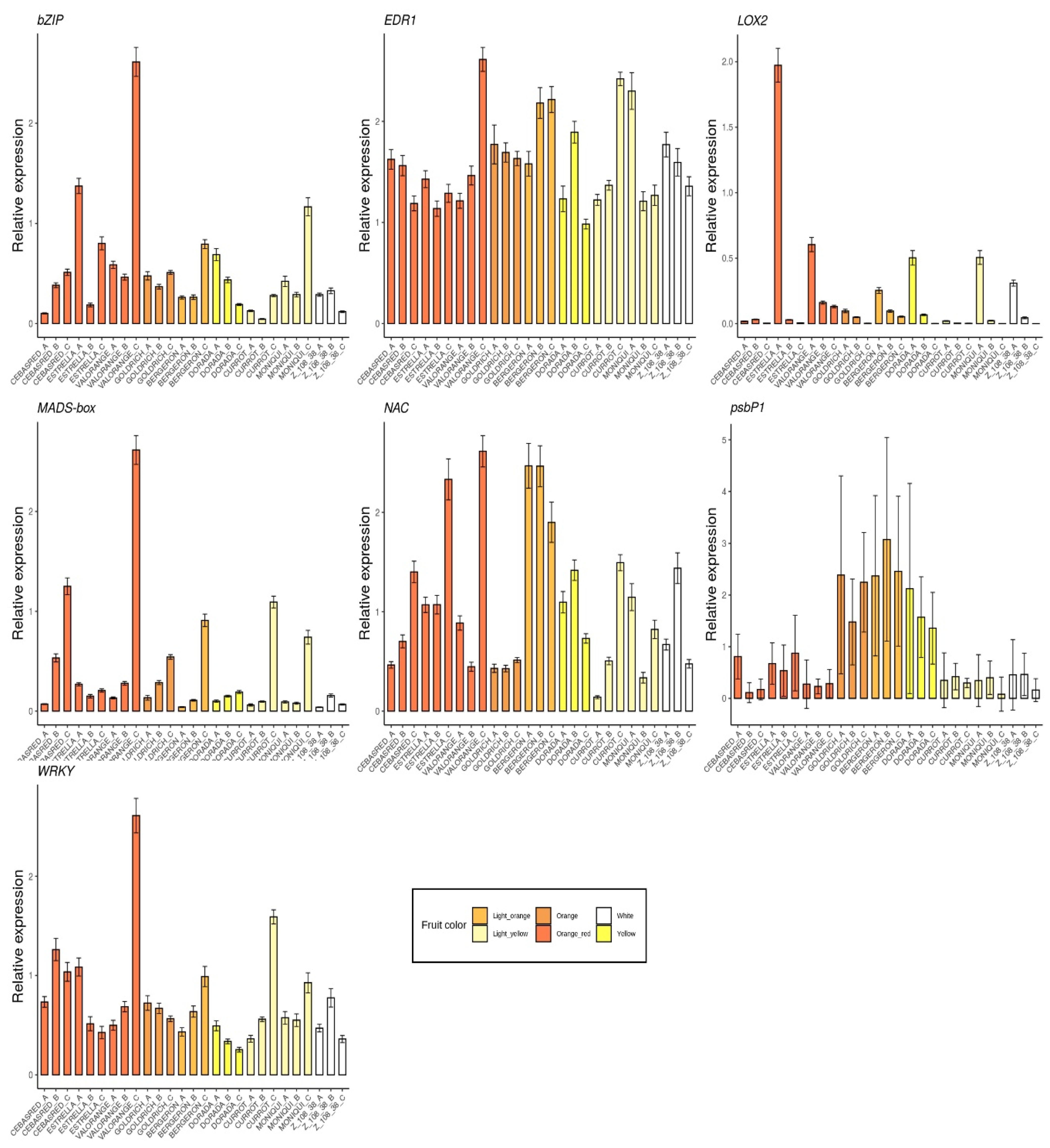
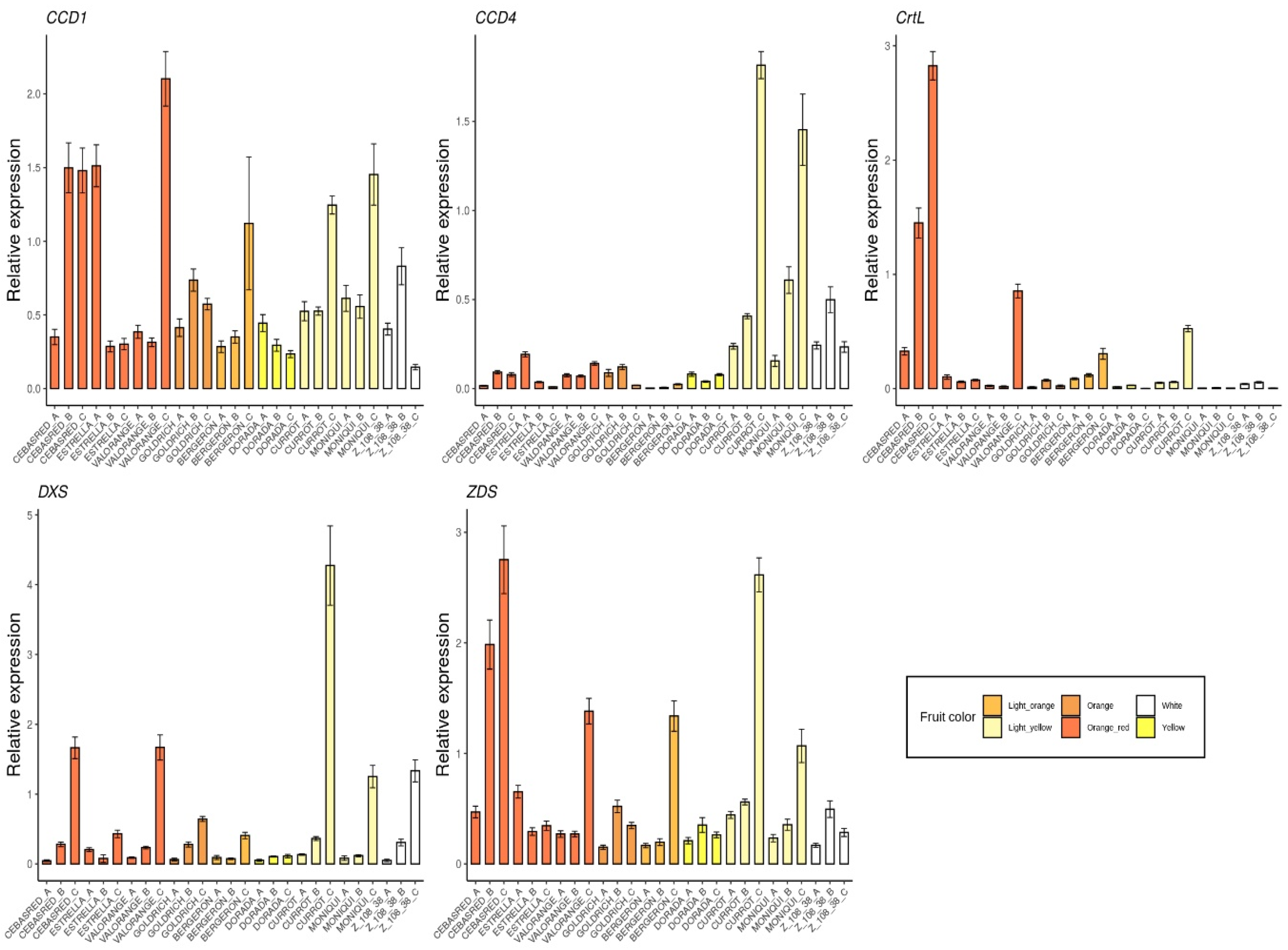
2.2. Monitoring Gene Expression of Candidate Genes
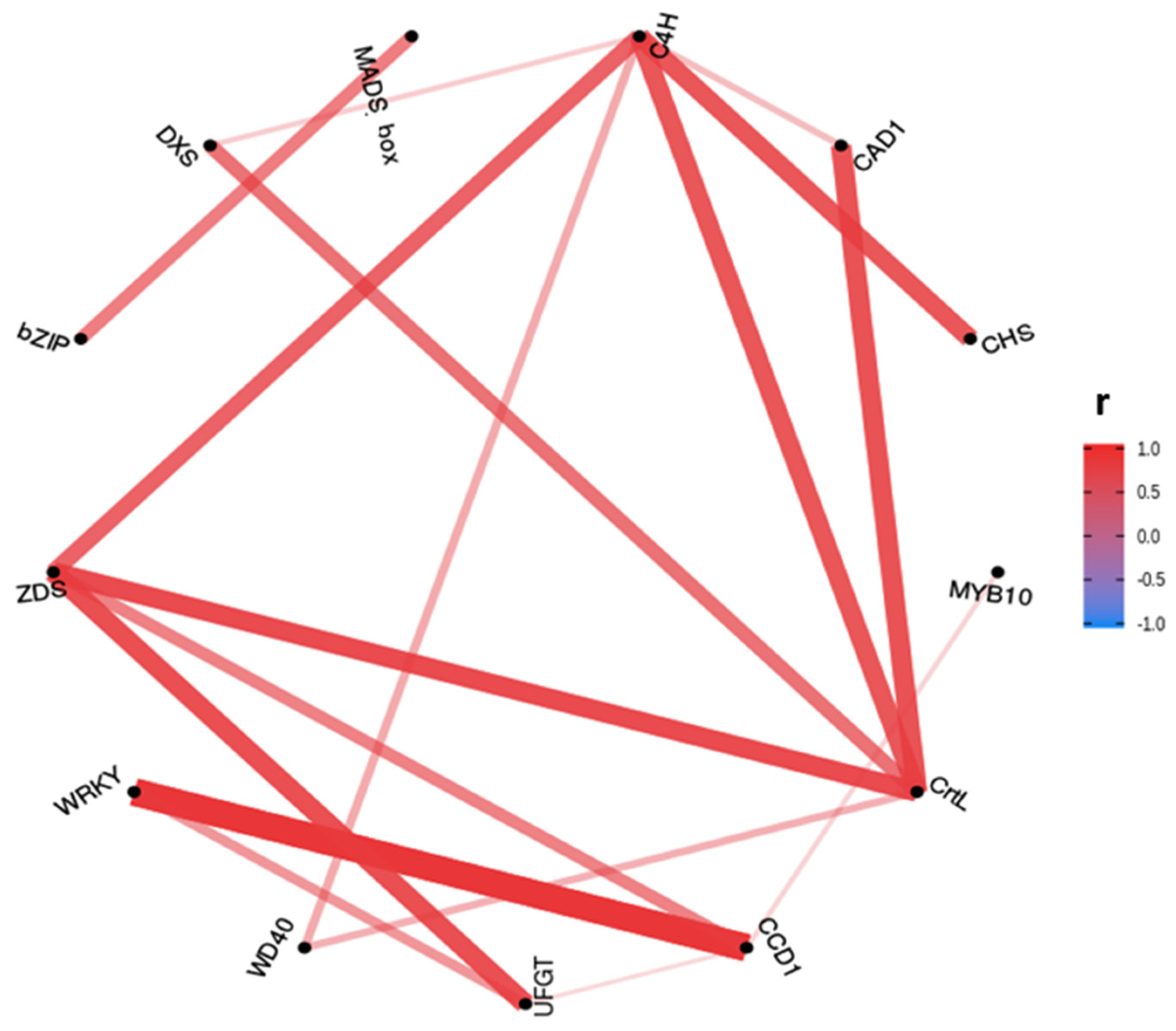
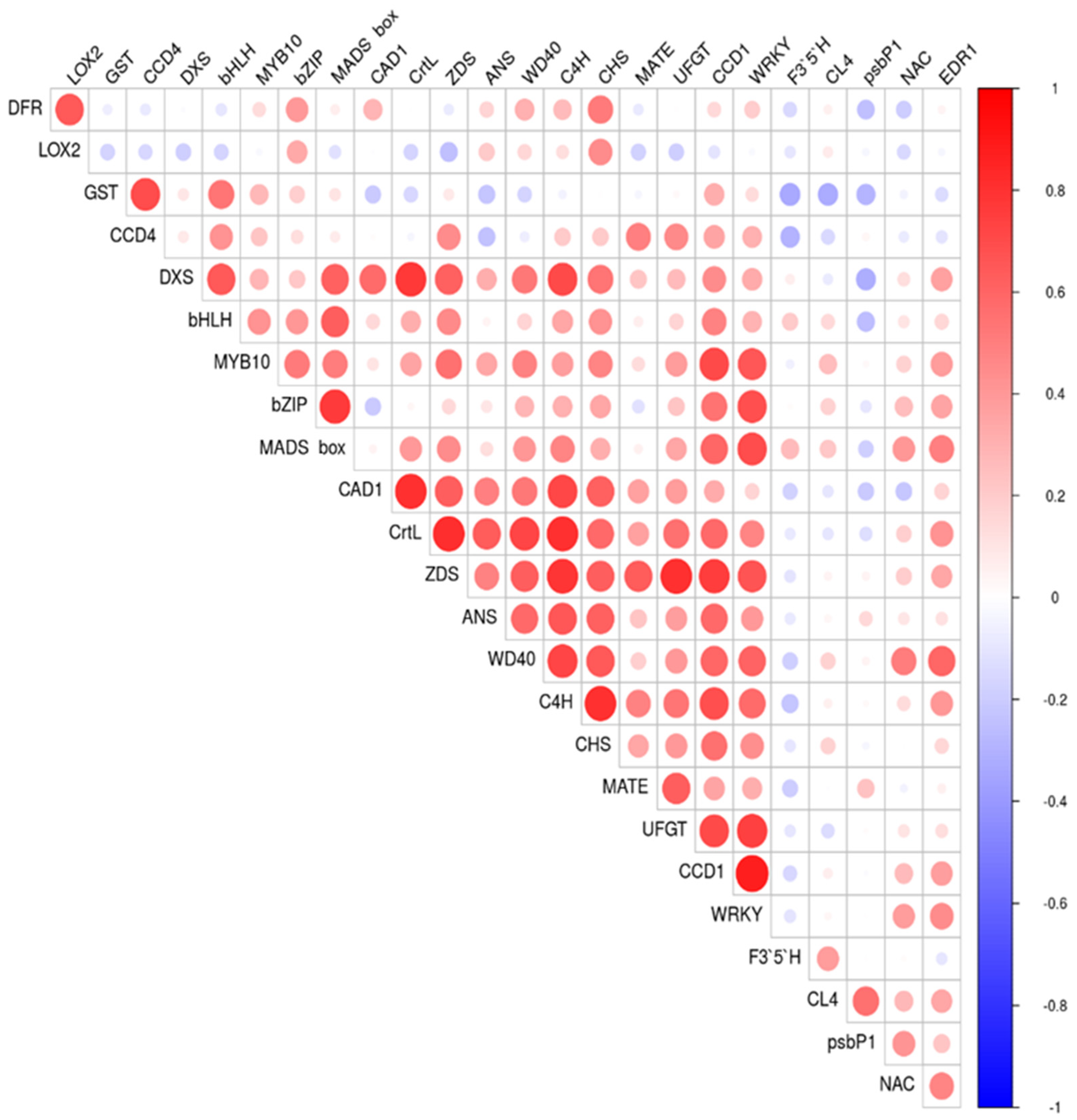
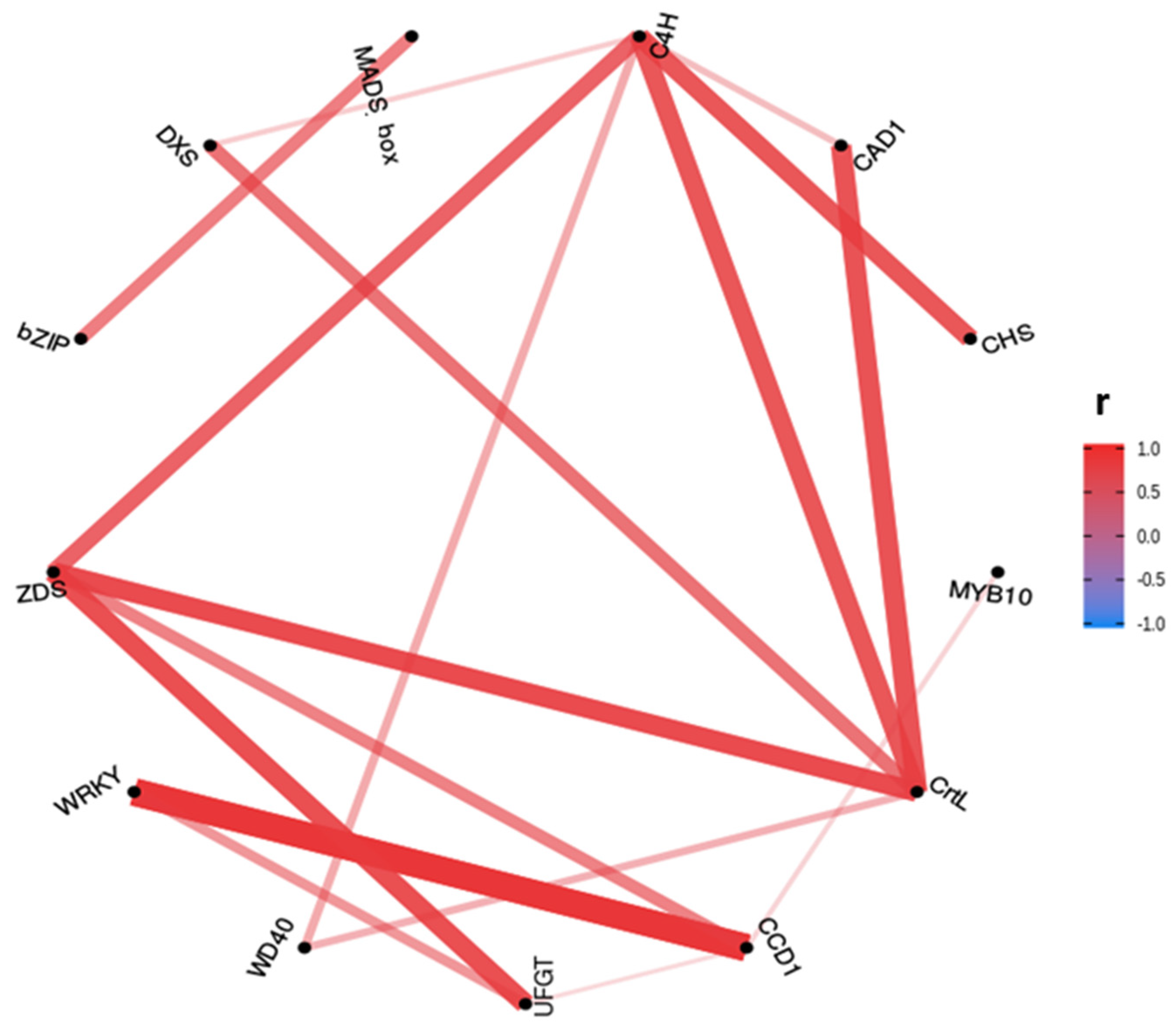
2.3. Correlation between Gene Expression of Assayed Genes
2.4. Multiple Linear Regression Model
2.4.1. Anthocyanins
2.4.2. Carotenoids
2.4.3. Anthocyanins and Carotenoids MLR Models Gene Correlation Prospection
3. Discussion
3.1. Multiple Linear Regression Model for Anthocyanins Content
3.2. Multiple Linear Regression Model for Carotenoids Content
3.3. Candidate Genes Descriptors for Ripening Stage
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Evaluation of Fruit Quality Traits
5.3. Gene Expression Analysis through RT-qPCR
5.4. Statistical Analysis and Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- García-Gómez, B.; Salazar, J.A.; Nicolás-Almansa, M.; Razi, M.; Rubio, M.; Ruiz, D.; Martínez-Gómez, P. Molecular Bases of Fruit Quality in Prunus Species: An Integrated Genomic, Transcriptomic, and Metabolic Review with a Breeding Perspective. Int. J. Mol. Sci. 2021, 22, 333. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; Ombra, M.N.; D’Acierno, A.; Cipriano, L.; Nazzaro, F. Apricots: Biochemistry and functional properties. Curr. Opin. Food Sci. 2018, 19, 23–29. [Google Scholar] [CrossRef]
- Ruiz, D.; Egea, J.; Tomás-Barberán, F.A.; Gil, M.I. Carotenoids from new apricot (Prunus armeniaca L.) varieties and their relationship with flesh and skin colour. J. Agric. Food Chem. 2005, 53, 6368–6374. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, D.; Egea, J.; Gil, M.I.; Tomás-Barberán, F.A. Characterization and quantitation of phenolic compounds in new apricot (Prunus armeniaca L.) varieties. J. Agric. Food Chem. 2005, 53, 9544–9552. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Aguero, M.; Troncoso, S.; Gudenschwager, O.; Campos-Vargas, R.; Moya-Leon, M.A.; Defilippi, B.G. Differential expression levels of aroma-related genes during ripening of apricot (Prunus armeniaca L.). Plant Physiol. Biochem. 2009, 47, 435–440. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, F.; Zhao, Y.; Shi, L.; Zhu, X. Cloning and expression analysis of polygalacturonase and pectin methylesterase genes during softening in apricot (Prunus armeniaca L.) fruit. Sci. Hort. 2019, 256, 108607. [Google Scholar] [CrossRef]
- Kovacs, E.; Nemeth-Szerdahelyi, E. β-Galactosidase activity and cell wall breakdown in apricots. J. Food Sci. 2002, 67, 2004–2008. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Rasori, A.; Bassi, D.; Geuna, F.; Ramina, A.; Tonutti, P.; Bonghi, C. Comparative transcript profiling of apricot (Prunus armeniaca L.) fruit development and on-tree ripening. Tree Genet. Gen. 2011, 7, 609–616. [Google Scholar] [CrossRef]
- Marty, I.; Bureau, S.; Sarkissian, G.; Gouble, B.; Audergon, J.M.; Albagnac, G. Ethylene regulation of carotenoid accumulation and carotenogenic gene expression in colour-contrasted apricot varieties (Prunus armeniaca). J. Exp. Bot. 2005, 56, 1877–1886. [Google Scholar] [CrossRef] [Green Version]
- Genard, M.; Bertin, N.; Borel, C.; Bussieres, P.; Gautier, H.; Habib, R.; Léchaudel, M.; Lecomte, A.; Lescourret, F.; Lobit, P.; et al. Towards a virtual fruit focusing on quality: Modelling features and potential uses. J. Exp. Bot. 2007, 58, 917–928. [Google Scholar] [CrossRef]
- Trainotti, L.; Bonghi, C.; Ziliotto, F.; Zanin, D.; Rasori, A.; Casadoro, G.; Tonutti, P. The use of microarray μPEACH1.0 to investigate transcriptome changes during transition from pre-climacteric to climacteric phase in peach fruit. Plant Sci. 2006, 170, 606–613. [Google Scholar] [CrossRef]
- Matas, A.J.; Gapper, N.E.; Chung, M.Y.; Giovannoni, J.J.; Rose, J.K. Biology and genetic engineering of fruit maturation for enhanced quality and shelf-life. Curr. Opin. Biotechnol. 2009, 20, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, J. Molecular Biology of fruit maturation and ripening. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 725–749. [Google Scholar] [CrossRef] [PubMed]
- Grimplet, J.; Romieu, C.; Audergon, J.M.; Marty, I.; Albagnac, G.; Lambert, P.; Terrier, N. Transcriptomic study of apricot fruit (Prunus armeniaca) ripening among 13.006 expressed sequence tags. Physiol. Plant. 2005, 125, 281–292. [Google Scholar] [CrossRef]
- Zhang, Q.; Feng, C.; Li, W.; Qu, Z.; Zeng, M.; Xi, W. Transcriptional regulatory networks controlling taste and aroma quality of apricot (Prunus armeniaca L.) fruit during ripening. BMC Genom. 2019, 20, 45. [Google Scholar] [CrossRef]
- Xi, W.; Zheng, H.; Zhang, Q.; Li, W. Profiling Taste and Aroma Compound Metabolism during Apricot Fruit Development and Ripening. Int. J. Mol. Sci. 2016, 17, 998. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Robredo, P.; Rubio, P.; Infante, R.; Campos-Vargas, R.; Manríquez, D.; González-Agüero, M.; Defilippi, B.G.G. Ethylene biosynthesis in apricot: Identification of a ripening-related 1-aminocyclopropane-1-carboxylic acid synthase (ACS) gene. Postharvest Biol. Technol. 2012, 63, 85–90. [Google Scholar] [CrossRef]
- Pfeiffer, P.; Pedryc, A.; Papp, N.; Abrankó, L.; Stefanovits-Bányai, É.; Hegedus, A. Molecular genetics of the flavonoid biosynthesis in two apricot genotypes. Acta Hort. 2012, 966, 107–112. [Google Scholar] [CrossRef]
- Xi, W.; Feng, J.; Liu, Y.; Zhang, S.; Zhao, G. The R2R3-MYB transcription factor PaMYB10 is involved in anthocyanin biosynthesis in apricots and determines red blushed skin. BMC Plant Biol. 2019, 19, 287. [Google Scholar] [CrossRef]
- Fan, X.; Shu, C.; Zhao, K.; Wang, X.; Cao, J.; Jiang, W. Regulation of apricot ripening and softening process during shelf life by post-storage treatments of exogenous ethylene and 1-methylcyclopropene. Sci. Hort. 2018, 232, 63–70. [Google Scholar] [CrossRef]
- Leida, C.; Ríos, G.; Soriano, J.M.; Pérez, B.; Llácer, G.; Crisosto, C.H.; Badenes, M.L. Identification and genetic characterization of an ethylene-dependent polygalacturonase from apricot fruit. Postharvest Biol. Technol. 2011, 62, 26–34. [Google Scholar] [CrossRef]
- García-Gómez, B.E.; Ruiz, D.; Salazar, J.A.; Rubio, M.; Martínez-García, P.J.; Martínez-Gómez, P. Analysis of Metabolites and Gene Expression Changes Relative to Apricot (Prunus armeniaca L.) Fruit Quality During Development and Ripening. Front. Plant Sci. 2020, 11, 1269. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.M.; Vupputuri, S.; Myers, L.; Whelton, P.K. Fruit and vegetable intake and risk of cardiovascular disease in US adults: The first National Health and Nutrition Examination Survey Epidemiologic Follow-up Study. Am. J. Clin. Nutr. 2002, 76, 93–99. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Giusti, M.M. Anthocyanins: Natural Colourants with Health-Promoting Properties. Ann. Rev. Food Sci. Tech. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M. Bioavailability, Metabolic Effects, and Safety. Ann. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Sochor, J.; Zitka, O.; Skutkova, H.; Pavlik, D.; Babula, P.; Krska, B.; Kizek, R. Content of Phenolic Compounds and Antioxidant Capacity in Fruits of Apricot Genotypes. Molecules 2010, 15, 6285–6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Derito, C.M.; Liu, M.K.; He, X.; Dong, M.; Liu, R.H. Cellular antioxidant activity of common vegetables. J. Agri. Food Chem. 2010, 58, 6621–6629. [Google Scholar] [CrossRef] [PubMed]
- Bureau, S.; Renard, C.M.G.C.; Reich, M.; Ginies, C.; Audergon, J.M. Change in anthocyanin concentrations in red apricot fruits during ripening. Food Sci. Technol. 2009, 42, 372–377. [Google Scholar] [CrossRef]
- Cheng, J.; Wei, G.; Zhou, H.; Gu, C.; Liao, L.; Han, Y. Unraveling the mechanism underlying the glycosylation and methylation of anthocyanins in peach. Plant Physiol. 2014, 166, 1044–1058. [Google Scholar] [CrossRef] [Green Version]
- Gabotti, D.; Negrini, N.; Morgutti, S.; Nocito, F.F.; Cocucci, M. Cinnamyl alcohol dehydrogenases in the mesocarp of ripening fruit of Prunus persica genotypes with different flesh characteristics: Changes in activity and protein and transcript levels. Physiol. Plant. 2015, 154, 329–348. [Google Scholar] [CrossRef]
- Zhou, H.; Lin-Wang, K.; Wang, H.; Gu, C.; Dare, A.P.; Espley, R.V.; He, H.; Allan, A.C.; Han, Y. Molecular genetics of blood-fleshed peach reveals activation of anthocyanin biosynthesis by NAC transcription factors. Plant J. 2015, 82, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.Z.; Zhou, D.R.; Ye, X.F.; Jiang, C.C.; Pan, S.L. Identification of Candidate Anthocyanin-Related Genes by Transcriptomic Analysis of “Furongli” Plum (Prunus salicina Lindl.) during Fruit Ripening Using RNA-Seq. Front. Plant Sci. 2016, 7, 1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, M.; Salazar, E.; Cabrera, S.; Olea, P.; Carrasco, B. Analysis of anthocyanin biosynthesis genes expression profiles in contrasting cultivars of Japanese plum (Prunus salicina L.) during fruit development. Gene Expr. Patterns 2016, 21, 54–62. [Google Scholar] [CrossRef]
- Usenik, V.; Stampar, F.; Veberic, R. Anthocyanins and fruit colour in plums (Prunus domestica L.) during ripening. Food Chem. 2009, 114, 529–534. [Google Scholar] [CrossRef]
- Zhang, Q.; Hao, R.J.; Xu, Z.D.; Yang, W.R.; Wang, J.; Cheng, T.R.; Pan, H.; Zhang, Q.X. Isolation and functional characterization of a R2R3-MYB regulator of Prunus mume anthocyanin biosynthetic pathway. Plant Cell Tiss. Organ Cult. 2017, 131, 417–429. [Google Scholar] [CrossRef]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takos, A.M.; Robinson, S.P.; Walker, A.R.L. Transcriptional regulation of the flavonoid pathway in the skin of dark-grown “Cripps” Red’ apples in response to sunlight. J. Hort. Sci. Biotechnol. 2006, 81, 735–744. [Google Scholar] [CrossRef]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Wang, Z.; Meng, D.; Wang, A.; Li, T.; Jiang, S.; Cong, P.; Li, T. The methylation of the PcMYB10 promoter is associated with green-skinned sport in Max Red Bartlett pear. Plant Physiol. 2013, 162, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Lin-Wang, K.; Bolitho, K.; Grafton, K.; Kortstee, A.; Karunairetnam, S.; McGhie, T.K.; Espley, R.V.; Hellens, R.P.; Allan, A.C. An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae. BMC Plant Biol. 2010, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Prinsi, B.; Negri, A.S.; Fedeli, C.; Morgutti, S.; Negrini, N.; Cocucci, M.; Espen, L. Peach fruit ripening: A proteomic comparative analysis of the mesocarp of two cultivars with different flesh firmness at two ripening stages. Phytochemistry 2011, 72, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Chen, X.; Zong, X.; Shu, H.; Gao, D.; Liu, Q. Comparative transcriptome analysis of 1944 genes involved in anthocyanin biosynthesis in the red and yellow fruits of sweet cherry 1945 (Prunus avium L.). PLoS ONE 2015, 10, e0121164. [Google Scholar]
- Gómez-Martínez, H.; Gil-Muñoz, F.; Bermejo, A.; Zuriaga, E.; Badenes, M.L. Insights of Phenolic Pathway in Fruits: Transcriptional and Metabolic Profiling in Apricot (Prunus armeniaca). Int. J. Mol. Sci. 2021, 22, 333. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar]
- Chagné, D.; Lin-Wang, K.; Espley, R.V.; Volz, R.K.; How, N.M.; Rouse, S.; Brendolise, C.; Carlisle, C.M.; Kumar, S.; De Silva, N.; et al. An ancient duplication of apple MYB transcription factors is responsible for novel red fruit-flesh phenotypes. Plant Physiol. 2013, 161, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Rahim, M.A.; Busatto, N.; Trainotti, L. Regulation of anthocyanin biosynthesis in peach fruits. Planta 2014, 240, 913–929. [Google Scholar] [CrossRef]
- Ravaglia, D.; Espley, R.V.; Henry-Kirk, R.A.; Andreotti, C.; Ziosi, V.; Hellens, R.P.; Allan, A.C. Transcriptional regulation of flavonoid biosynthesis in nectarine (Prunus persica) by a set of R2R3 MYB transcription factors. BMC Plant Biol. 2013, 13, 68. [Google Scholar] [CrossRef] [Green Version]
- Tuan, P.A.; Bai, S.; Yaegaki, H.; Tamura, T.; Hihara, S.; Moriguchi, T.; Oda, K. The crucial role of PpMYB10.1 in anthocyanin accumulation in peach and relationships between its allelic type and skin colour phenotype. BMC Plant Biol. 2015, 15, 280. [Google Scholar] [CrossRef] [Green Version]
- González, M.; Carrasco, B.; Salazar, E. Genome-wide identification and characterization of R2R3MYB family in Rosaceae. Genom. Data 2016, 9, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Fiol, A.; García-Gómez, B.E.; Jurado-Ruiz, F.; Alexiou, K.; Howad, W.; Aranzana, M.J. Characterization of Japanese Plum (Prunus salicina) PsMYB10 alleles reveals structural variation and polymorphisms correlating with fruit skin colour. Front. Plant Sci. 2021, 12, 655267. [Google Scholar] [CrossRef]
- Hernandez, J.M.; Heine, G.F.; Irani, N.G.; Feller, A.; Kim, M.G.; Matulnik, T.; Chandler, V.L.; Grotewold, E. Different mechanisms participate in the R-dependent activity of the R2R3 MYB transcription factor C1. J. Biol. Chem. 2004, 279, 48205–48213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espley, R.V.; Brendolise, C.; Chagne, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple Repeats of a Promoter Segment Causes Transcription Factor Autoregulation in Red Apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, B.; Qi, Y.; Chen, X.; Liu, C.; Liu, Z.; Ren, X. Expression differences of pigment structural genes and transcription factors explain flesh colouration in three contrasting kiwifruit cultivars. Front. Plant Sci. 2017, 8, 1057. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Zheng, D.; Han, Y.; Khan, M.A.; Soria-Guerra, R.E.; Korban, S.S. Transcriptome analysis of the exocarp of apple fruit identifies light-induced genes involved in red colour pigmentation. Gene 2014, 534, 78–87. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Sun, Y.; Qian, M.; Wu, R.; Niu, Q.; Teng, Y.; Zhang, D. Postharvest pigmentation in red Chinese sand pears (Pyrus pyrifolia Nakai) in response to optimum light and temperature. Postharvest Biol. Technol. 2014, 91, 64–71. [Google Scholar] [CrossRef]
- Feng, F.; Li, M.; Ma, F.; Cheng, L. Phenylpropanoid metabolites and expression of key genes involved in anthocyanin biosynthesis in the shaded peel of apple fruit in response to sun exposure. Plant Physiol. Biochem. 2013, 69, 54–61. [Google Scholar] [CrossRef]
- Honda, C.; Kotoda, N.; Wanda, M.; Kondo, S.; Soejima, J.; Zhang, Z.; Tsuda, T.; Moriguchi., T. Anthocyanin biosynthetic genes are coordinately expressed during red coloration in apple skin. Plant Physiol. Biochem. 2002, 11, 955–962. [Google Scholar] [CrossRef]
- Takos, A.M.; Jaffe, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [Green Version]
- Telias, A.M.; Lin-Wang, K.; Stevenson, D.E.; Cooney, J.M.; Hellens, R.P.; Allan, A.C.; Hoover, E.E.; Bradeen, J.M. Apple skin patterning is associated with differential expression of MYB10. BMC Plant Biol. 2011, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.; Park, E.; Choi, G. PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. Plant J. 2007, 49, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Choi, M.; Kim, K.; Bang, G.; Cho, M.; Choi, S.-B.; Choi, G.; Park, Y.-I. HY5 regulates anthocyanin biosynthesis by inducing the transcriptional activation of the MYB75/PAP1 transcription factor in Arabidopsis. FEBS Lett. 2013, 587, 1543–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, A.; Schrader, A.; Kokkelink, L.; Falke, C.; Welter, B.; Iniesto, E.; Rubio, V.; Uhrig, J.F.; Hülskamp, M.; Hoecker, U. Light and the E3 ubiquitin ligase COP1/SPA control the protein stability of the MYB transcription factors PAP1 and PAP2 involved in anthocyanin accumulation in Arabidopsis. Plant J. 2013, 74, 638–651. [Google Scholar] [CrossRef]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagasser, M. A. thaliana TRANSPARENT TESTA 1 is involved in seed coat development and defines the WIP subfamily of plant zinc finger proteins. Genes Dev. 2002, 16, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Nesi, N.; Debeaujon, I.; Jond, C.; Stewart, A.J.; Jenkins, G.I.; Caboche, M.; Lepiniec, L. The TRANSPARENT TESTA16 Locus Encodes the ARABIDOPSIS BSISTER MADS Domain Protein and Is Required for Proper Development and Pigmentation of the Seed Coat. Plant Cell 2002, 14, 2463. [Google Scholar] [CrossRef] [Green Version]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC transcription factor, ANAC078, regulates flavonoid biosynthesis under high-light. Plant Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/bHLH/MYB Complexes to Regulate Jasmonate-Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Gou, J.-Y.; Felippes, F.F.; Liu, C.-J.; Weigel, D.; Wang, J.-W. Negative Regulation of Anthocyanin Biosynthesis in Arabidopsis by a miR156-Targeted SPL Transcription Factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, A.; Brockman, A.; Aguirre, L.; Campbell, A.; Bean, A.; Cantero, A.; Gonzalez, A. Advances in the MYB-bHLH-WD Repeat (MBW) pigment regulatory model: Addition of a WRKY factor and co-option of an anthocyanin MYB for betalain regulation. Plant Cell Physiol. 2017, 58, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Ayour, J.; Sagar, M.; Alfeddy, M.N.; Taourirte, M.; Benichou, M. Evolution of pigments and their relationship with skin colour based on ripening in fruits of different Moroccan genotypes of apricots (Prunus armeniaca L.). Sci. Hort. 2016, 207, 168–175. [Google Scholar] [CrossRef]
- Ruiz, D.; Reich, M.; Bureau, S.; Renard, C.M.G.C.; Audergon, J.M. Application of reflectance colourimeter measurements and infrared spectroscopy methods to rapid and nondestructive evaluation of carotenoids content in apricot (Prunus armeniaca L.). J. Agric. Food Chem. 2008, 56, 4916–4922. [Google Scholar] [CrossRef] [PubMed]
- Adami, M.; De Franceschi, P.; Brandi, F.; Liverani, A.; Giovannini, D.; Rosati, C.; Dondini, L.; Tartarini, S. Identifying a Carotenoid Cleavage Dioxygenase (ccd4) Gene Controlling Yellow/White Fruit Flesh Colour of Peach. Plant Mol. Biol. Rep. 2013, 31, 1166–1175. [Google Scholar] [CrossRef]
- Cao, S.; Liang, M.; Shi, L.; Shao, J.; Song, C.; Bian, K.; Chen, W.; Yang, Z. Accumulation of carotenoids and expression of carotenogenic genes in peach fruit. Food Chem. 2017, 214, 137–146. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Kita, M.; Ban, Y.; Honda, C.; Yaegaki, H.; Moriguchi, T.; Kato, M.; Ikoma, Y. Carotenoid accumulation in Japanese apricot (Prunus mume Siebold & Zucc.): Molecular analysis of carotenogenic gene expression and ethylene regulation. J. Agric. Food Chem. 2007, 55, 3414–3420. [Google Scholar]
- Khan, A.S. Differences in fruit growth and ripening of early-, mid- and late-season maturing Japanese plum cultivars. Fruits 2016, 71, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Ohmiya, A. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotech. 2008, 19, 190–197. [Google Scholar] [CrossRef]
- Frye, C.A.; Tang, D.; Innes, R.W. Negative regulation of defense responses in plants by a conserved MAPKK kinase. Proc. Nat. Acad. Sci. USA 2001, 98, 373–378. [Google Scholar] [CrossRef]
- Bouvier, F.; D’Harlingue, A.; Backhaus, R.A.; Kumagai, M.H.; Camara, B. Identification of neoxanthin synthase as a carotenoid cyclase paralog. Eur. J. Bioch. 2000, 267, 6346–6352. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Shen, J.Y.; Wei, W.W.; Xi, W.P.; Xu, C.J.; Ferguson, I.; Chen, K.S. Expression of Genes Associated with Aroma Formation Derived from the Fatty Acid Pathway during Peach Fruit Ripening. J. Agric. Food Chem. 2010, 58, 6157–6165. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, J.; Zhao, J.; Zhou, H.; Ren, F.; Wang, L.; Han, Y. Inactivation of a Gene Encoding Carotenoid Cleavage Dioxygenase (CCD4) Leads to Carotenoid-Based Yellow Colouration of Fruit Flesh and Leaf Midvein in Peach. Plant Mol. Biol. Rep. 2014, 32, 246–257. [Google Scholar] [CrossRef]
- Falchi, R.; Vendramin, E.; Zanon, L.; Scalabrin, S.; Cipriani, G.; Verde, I.; Morgante, M. Three distinct mutational mechanisms acting on a single gene underpin the origin of yellow flesh in peach. Plant J. 2013, 76, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovisetto, A.; Guzzo, F.; Tadiello, A.; Confortin, E.; Pavanello, A.; Botton, A.; Casadoro, G. Characterization of a bZIP gene highly expressed during ripening of the peach fruit. Plant Physiol. Bioch. 2013, 70, 462–470. [Google Scholar] [CrossRef]
- Costa, F.; Alba, R.; Schouten, H.; Soglio, V.; Gianfranceschi, L.; Serra, S.; Giovannoni, J. Use of homologous and heterologous gene expression profiling tools to characterize transcription dynamics during apple fruit maturation and ripening. BMC Plant Biol. 2010, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, A.; De Vos, C.H.; Wein, M.; Sun, Z.; Greco, R.; Kroon, A.; Mol, J.N.M.; Connell, A.P.O. The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. Plant J. 2010, 28, 1–15. [Google Scholar] [CrossRef]
- Dragovic-Uzelac, V.; Levaj, B.; Mrkic, V.; Bursac, D.; Boras, M. The content of polyphenols and carotenoids in three apricot cultivars depending on stage of maturity and geographical region. Food Chem. 2007, 102, 966–975. [Google Scholar] [CrossRef]
- Pirona, R.; Eduardo, I.; Pacheco, I.; Da Silva Linge, C.; Miculan, M.; Verde, I.; Rossini, L. Fine mapping and identification of a candidate gene for a major locus controlling maturity date in peach. BMC Plant Biol. 2013, 13, 166. [Google Scholar] [CrossRef] [Green Version]
- Crifò, T.; Petrone, G.; Lo Cicero, L.; Lo Piero, A.R. Short Cold Storage Enhances the Anthocyanin Contents and Level of Transcripts Related to Their Biosynthesis in Blood Oranges. J. Agric. Food Chem. 2012, 60, 476–481. [Google Scholar] [CrossRef]
- Lü, P.; Yu, S.; Zhu, N.; Chen, Y.-R.; Zhou, B.; Pan, Y.; Tzeng, D.; Fabi, J.P.; Argyris, J.; Garcia-Mas, J.; et al. Genome encode analyses reveal the basis of convergent evolution of fleshy fruit ripening. Nat. Plants 2018, 4, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Gramzow, L.; Theissen, G. A hitchhiker’s guide to the MADS world of plants. Genome Biol. 2010, 11, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Zhang, Q.; Sun, L.; Du, D.; Cheng, T.; Pan, H.; Wang, J. Genome-wide identification, characterisation and expression analysis of the MADS-box gene family in Prunus mume. Mol. Genet. Gen. 2014, 289, 903–920. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Poole, M.; Jones, M.O.; Kamarainen-Karppinen, T.; Koskimaki, J.J.; Hohtola, A.; Seymour, G.B. A SQUAMOSA MADS Box Gene Involved in the Regulation of Anthocyanin Accumulation in Bilberry Fruits. Plant Physiol. 2010, 153, 1619–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhao, G.; Yang, Y.-N.; Le, W.-Q.; Khan, M.A.; Zhang, S.-L.; Gu, C.; Huang, W.-J. Identification of differentially expressed genes related to colouration in red/green mutant pear (Pyrus communis L.). Tree Genet. Gen. 2013, 9, 75–83. [Google Scholar] [CrossRef]
- Guo, S.; Song, Z.; Ma, R.; Yang, Y.; Yu, M. Genome-wide identification and expression analysis of the lipoxygenase gene family during peach fruit ripening under different postharvest treatments. Acta Physiol. Plant. 2017, 39, 111. [Google Scholar] [CrossRef]
- Kausch, K.D.; Handa, A.K. Molecular cloning of a ripening-specific lipoxygenase and its expression during wild-type and mutant tomato fruit development. Plant Physiol. 1997, 113, 1041–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eduardo, I.; Chietera, G.; Pirona, R.; Pacheco, I.; Troggio, M.; Banchi, E.; Bassi, D.; Rossini, L.; Vecchietti, A.; Pozzi, C.M. Genetic dissection of aroma volatile compounds from the essential oil of peach fruit: QTL analysis and identification of candidate genes using dense SNP maps. Tree Genet. Gen. 2013, 9, 189–204. [Google Scholar] [CrossRef]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- García-Gómez, B.E.; Salazar, J.A.; Dondini, L.; Martínez-Gómez, P.; Ruiz, D. Identification of QTLs linked to fruit quality traits in apricot (Prunus armeniaca L) and biological validation through gene expression analysis using qPCR. Mol. Breed. 2019, 39, 28. [Google Scholar] [CrossRef]
- García-Gómez, B.; Razi, M.; Salazar, J.A.; Prudencio, A.S.; Ruiz, D.; Dondini, L.; Martínez-Gómez, P. Comparative Analysis of SSR Markers Developed in Exon, Intron, and Intergenic Regions and Distributed in Regions Controlling Fruit Quality Traits in Prunus Species: Genetic Diversity and Association Studies. Plant Mol. Biol. Rep. 2018, 36, 23–35. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Rennes, A.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Statist. Software 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Taiyun, W.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix, version 0; Comprehensive R Archive Network (CRAN): Vienna, Austria, 2017.
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; 325p. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Trait | Mean | SD | Cultivar | Trait | Mean | SD | Cultivar | Quality Trait | Mean | SD |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ‘Bergeron’ | Fruit weight | 81.0 | 5.23 | ‘Dorada’ | Fruit weight | 78.4 | 4.95 | ‘Moniquí’ | Fruit weight | 68.6 | 7.75 |
| Skin color | 80.5 | 2.45 | Skin color | 84.0 | 6.75 | Skin color | 95.2 | 3.38 | |||
| Flesh color | 77.7 | 3.13 | Flesh color | 86.3 | 7.36 | Flesh color | 94.7 | 1.37 | |||
| SSC | 12.3 | 0.72 | SSC | 12.5 | 0.97 | SSC | 14.5 | 0.265 | |||
| pH | 3.42 | 0.09 | pH | 3.72 | 0.10 | pH | 3.49 | 0.041 | |||
| Firmness | 32.0 | 2.86 | Firmness | 32.1 | 3.44 | Firmness | 53.7 | 11.8 | |||
| Chlorophylls a | 2.58 | 0.45 | Chlorophylls a | 1.63 | 0.63 | Chlorophylls a | 1.5 | 1.32 | |||
| Chlorophylls b | 1.82 | 0.05 | Chlorophylls b | 2.36 | 0.79 | Chlorophylls b | 1.93 | 1.18 | |||
| Anthocyanins | 1.02 | 0.34 | Anthocyanins | 1.90 | 0.54 | Anthocyanins | 1.24 | 0.35 | |||
| Carotenoids | 69.9 | 4.40 | Carotenoids | 29.6 | 1.18 | Carotenoids | 3.53 | 1.62 | |||
| Ripening date | 175 | 1.73 | Ripening date | 178 | 2.52 | Ripening date | 165 | 2 | |||
| ‘Cebasred’ | Fruit weight | 50.4 | 2.03 | ‘Estrella’ | Fruit weight | 90.3 | 10.7 | ‘Valorange’ | Fruit weight | 64.6 | 8.28 |
| Skin color | 64.0 | 0.62 | Skin color | 78.3 | 4.02 | Skin color | 82.9 | 3.88 | |||
| Flesh color | 71.1 | 0.35 | Flesh color | 79.6 | 1.13 | Flesh color | 78.4 | 3.32 | |||
| SSC | 11.9 | 0.62 | SSC | 14.1 | 0.50 | SSC | 13.1 | 1.10 | |||
| pH | 3.76 | 0.05 | pH | 3.44 | 0.01 | pH | 3.49 | 0.15 | |||
| Firmness | 36.5 | 4.83 | Firmness | 31.8 | 2.16 | Firmness | 50.0 | 8.03 | |||
| Chlorophylls a | 1.21 | 0.66 | Chlorophylls a | 1.59 | 0.86 | Chlorophylls a | 1.44 | 0.10 | |||
| Chlorophylls b | 1.69 | 0.47 | Chlorophylls b | 2.03 | 1.16 | Chlorophylls b | 1.15 | 0.16 | |||
| Anthocyanins | 10.6 | 0.46 | Anthocyanins | 17.8 | 0.18 | Anthocyanins | 18.7 | 1.21 | |||
| Carotenoids | 73.6 | 0.51 | Carotenoids | 90.0 | 8.85 | Carotenoids | 100 | 4.80 | |||
| Ripening date | 134 | 3.21 | Ripening date | 149 | 2.08 | Ripening date | 157 | 0.57 | |||
| ‘Currot’ | Fruit weight | 49.2 | 6.62 | ‘Goldrich’ | Fruit weight | 91.9 | 1.94 | ‘Z108-38′ | Fruit weight | 65.5 | 2.49 |
| Skin color | 89.9 | 2.33 | Skin color | 76.6 | 3.22 | Skin color | 92.3 | 1.57 | |||
| Flesh color | 87.2 | 1.07 | Flesh color | 71.6 | 4.03 | Flesh color | 92.5 | 1.39 | |||
| SSC | 15.1 | 1.10 | SSC | 13.5 | 0.81 | SSC | 13.7 | 0.25 | |||
| pH | 3.87 | 0.13 | pH | 3.12 | 0.08 | pH | 4.04 | 0.13 | |||
| Firmness | 25.6 | 4.09 | Firmness | 42.7 | 5.21 | Firmness | 35.1 | 2.30 | |||
| Chlorophylls a | 0.66 | 0.12 | Chlorophylls a | 0.92 | 0.71 | Chlorophylls a | 2.60 | 0.03 | |||
| Chlorophylls b | 0.93 | 0.24 | Chlorophylls b | 0.97 | 0.63 | Chlorophylls b | 3.78 | 1.17 | |||
| Anthocyanins | 0.51 | 0.64 | Anthocyanins | 0.62 | 0.12 | Anthocyanins | 1.24 | 0.70 | |||
| Carotenoids | 7.58 | 0.14 | Carotenoids | 77.8 | 2.46 | Carotenoids | 11.2 | 0.50 | |||
| Ripening date | 139 | 4.16 | Ripening date | 164 | 1 | Ripening date | 15 | 2 |
| Cultivar | Pedigree | Origin | Ripening | Fruit Color | Ethylene Production |
|---|---|---|---|---|---|
| Bergeron‘ | Unknown | France | Very late | Light orange | Very low |
| ‘Cebasred‘ | ‘5–26‘ × ‘Colorado‘ | Spain | Very early | Orange–Red | Low |
| ‘Currot‘ | Unknown | Spain | Very early | Light yellow | High |
| ‘Dorada‘ | ‘Bergeron‘ × ‘Moniquí‘ | Spain | Late | Yellow | Low |
| ‘Estrella‘ | ‘Orange–Red‘ × ‘Z211-18‘ | Spain | Medium | Orange–Red | Medium |
| ‘Goldrich‘ | ‘Sunglo‘ × ‘Perfection‘ | USA | Late | Orange | Very low |
| ‘Moniquí‘ | Unknown | Spain | Medium | Light yellow | High |
| ‘Valorange‘ | ‘Orange–Red‘ × ‘Currot‘ | Spain | Medium | Orange–Red | Low |
| ‘Z108-38‘ | ‘Gitano’ × ‘Pepito del Rubio’ | Spain | Late | White | High |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Gómez, B.E.; Salazar, J.A.; Egea, J.A.; Rubio, M.; Martínez-Gómez, P.; Ruiz, D. Monitoring Apricot (Prunus armeniaca L.) Ripening Progression through Candidate Gene Expression Analysis. Int. J. Mol. Sci. 2022, 23, 4575. https://doi.org/10.3390/ijms23094575
García-Gómez BE, Salazar JA, Egea JA, Rubio M, Martínez-Gómez P, Ruiz D. Monitoring Apricot (Prunus armeniaca L.) Ripening Progression through Candidate Gene Expression Analysis. International Journal of Molecular Sciences. 2022; 23(9):4575. https://doi.org/10.3390/ijms23094575
Chicago/Turabian StyleGarcía-Gómez, Beatriz E., Juan A. Salazar, Jose A. Egea, Manuel Rubio, Pedro Martínez-Gómez, and David Ruiz. 2022. "Monitoring Apricot (Prunus armeniaca L.) Ripening Progression through Candidate Gene Expression Analysis" International Journal of Molecular Sciences 23, no. 9: 4575. https://doi.org/10.3390/ijms23094575
APA StyleGarcía-Gómez, B. E., Salazar, J. A., Egea, J. A., Rubio, M., Martínez-Gómez, P., & Ruiz, D. (2022). Monitoring Apricot (Prunus armeniaca L.) Ripening Progression through Candidate Gene Expression Analysis. International Journal of Molecular Sciences, 23(9), 4575. https://doi.org/10.3390/ijms23094575