
Two Male-Specific Antimicrobial Peptides SCY2 and Scyreprocin as Crucial Molecules Participated in the Sperm Acrosome Reaction of Mud Crab Scylla paramamosain


Abstract
:1. Introduction
2. Results
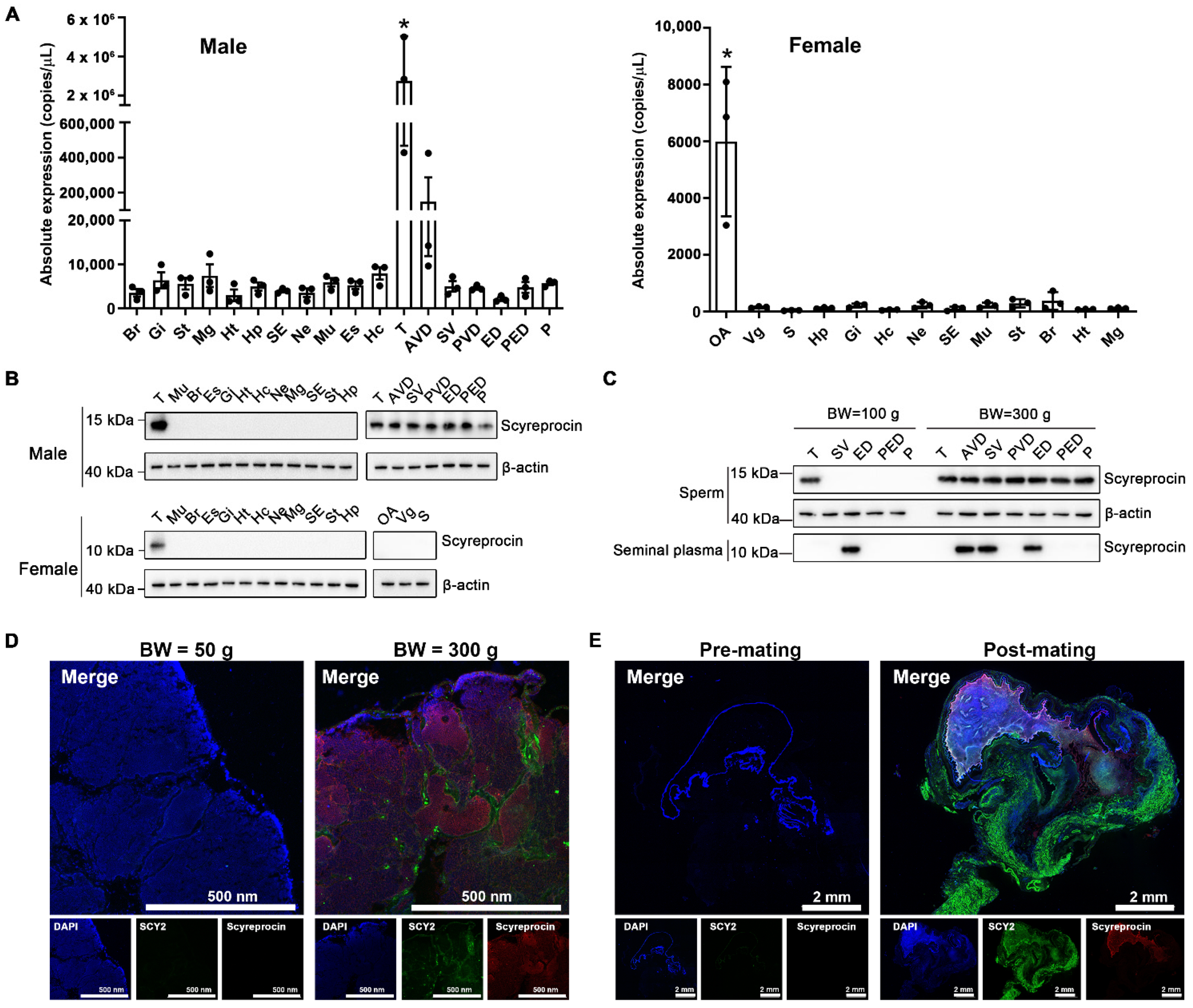
2.1. Expression Pattern of Scyreprocin and SCY2 In Vivo
2.2. Scyreprocin and SCY2 Responded to Bacterial Infection In Vivo
2.3. Subcellular Localization of Scyreprocin and SCY2 in Mud Crab Sperm
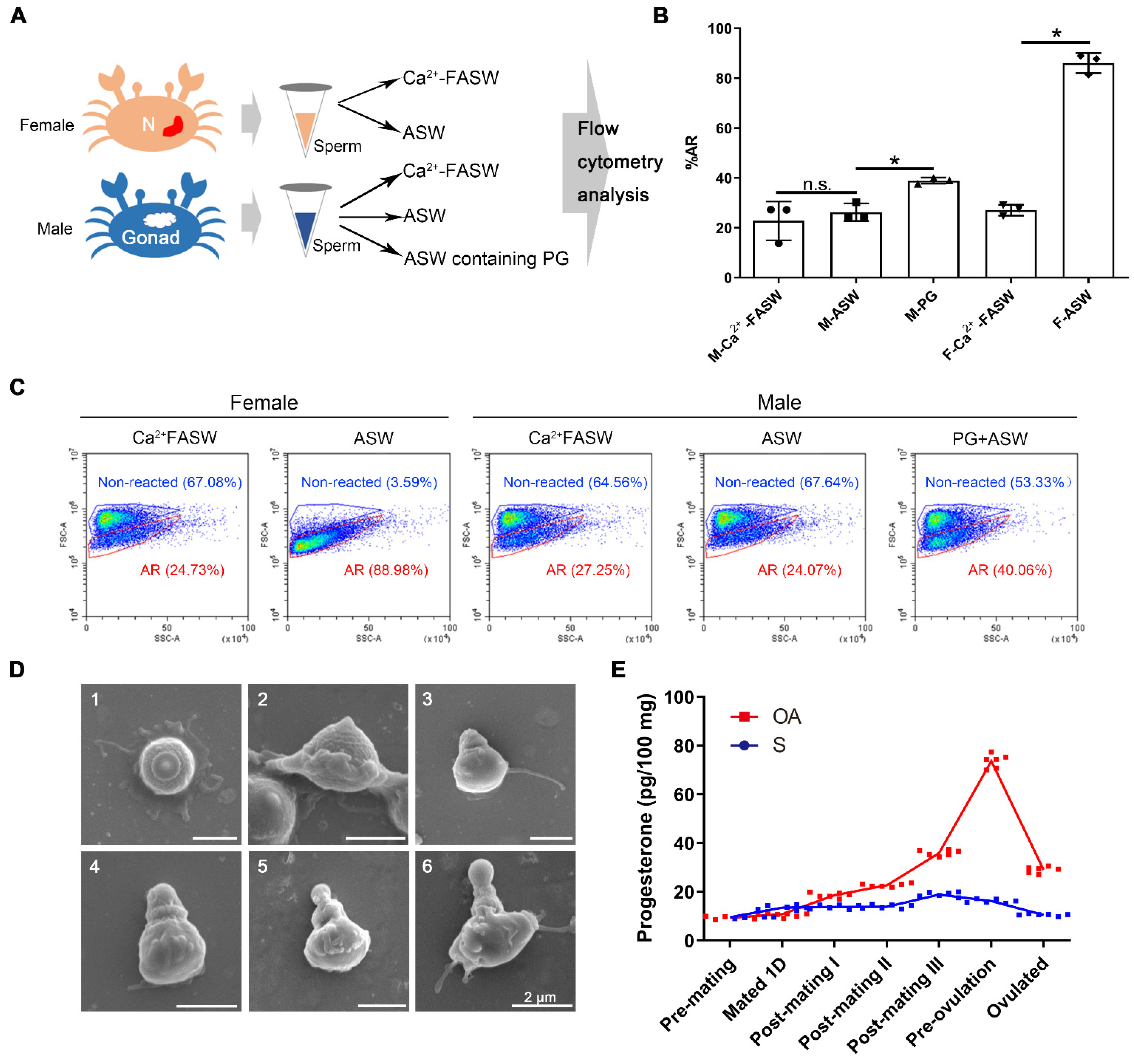
2.4. Progesterone Induced In Vitro Sperm Acrosome Reaction of S. paramamosain
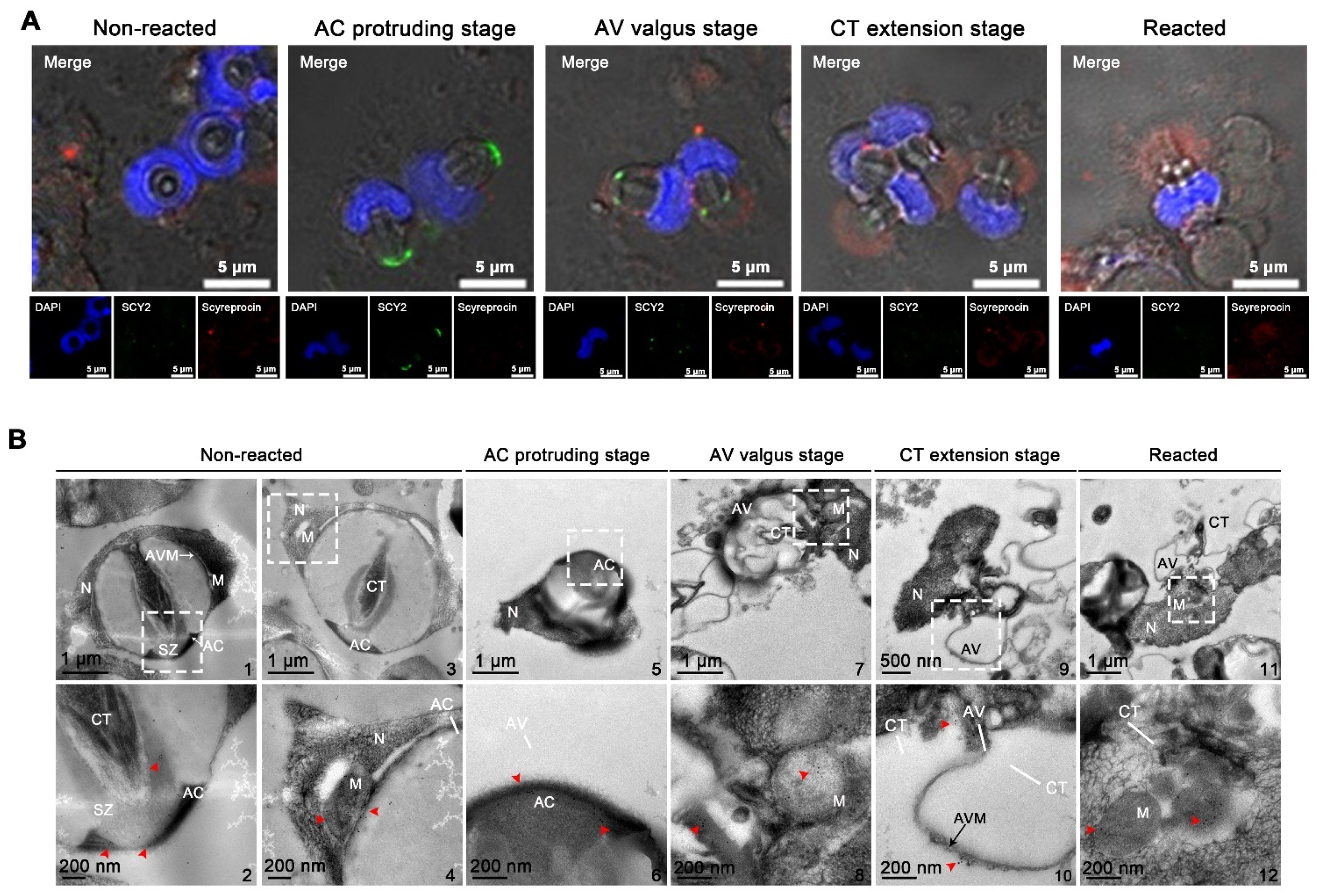
2.5. Localizations of SCY2 and Scyreprocin in Sperm during Sperm Acrosome Reaction
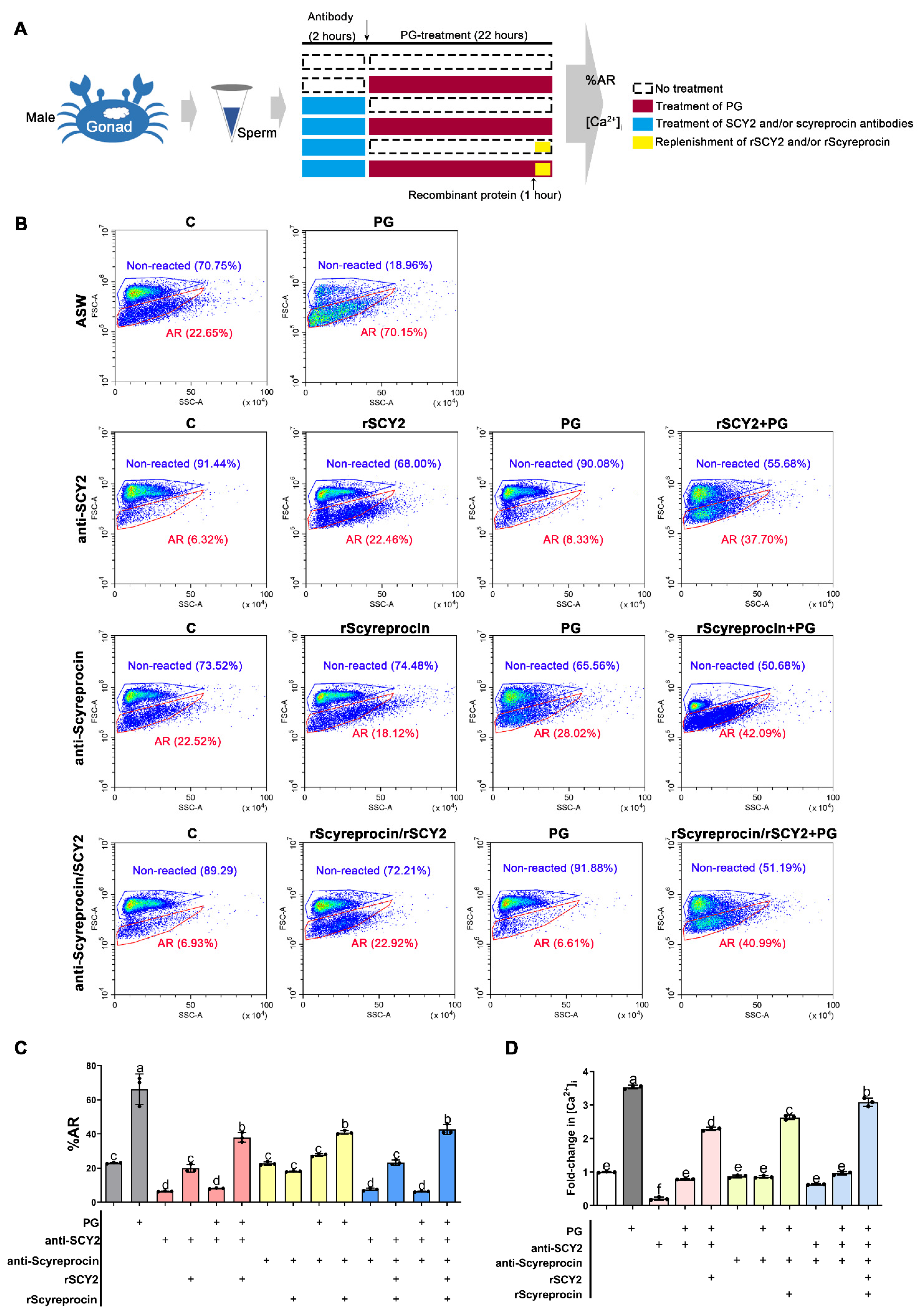
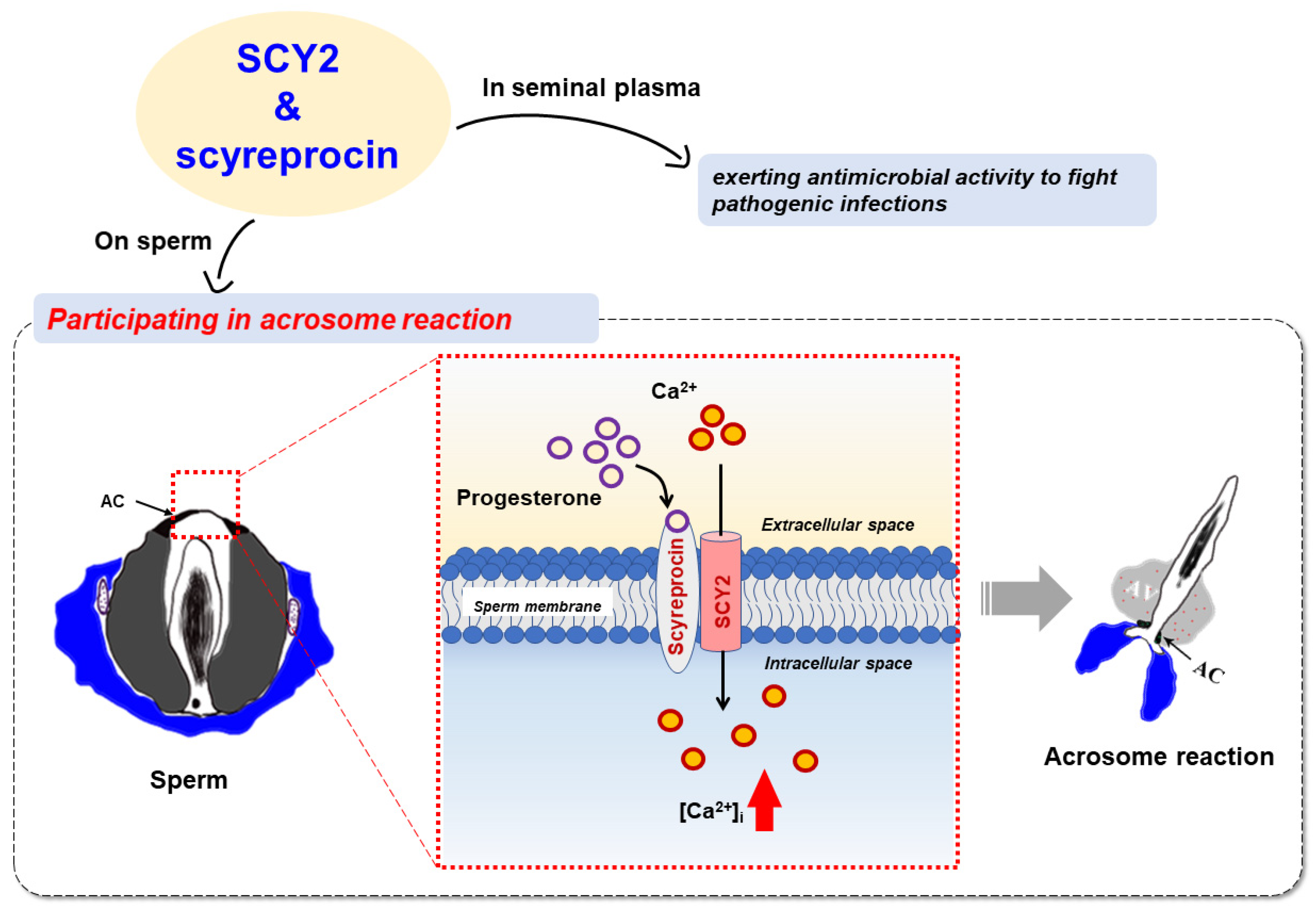
2.6. SCY2 and Scyreprocin Participated in Progesterone-Induced Acrosome Reaction
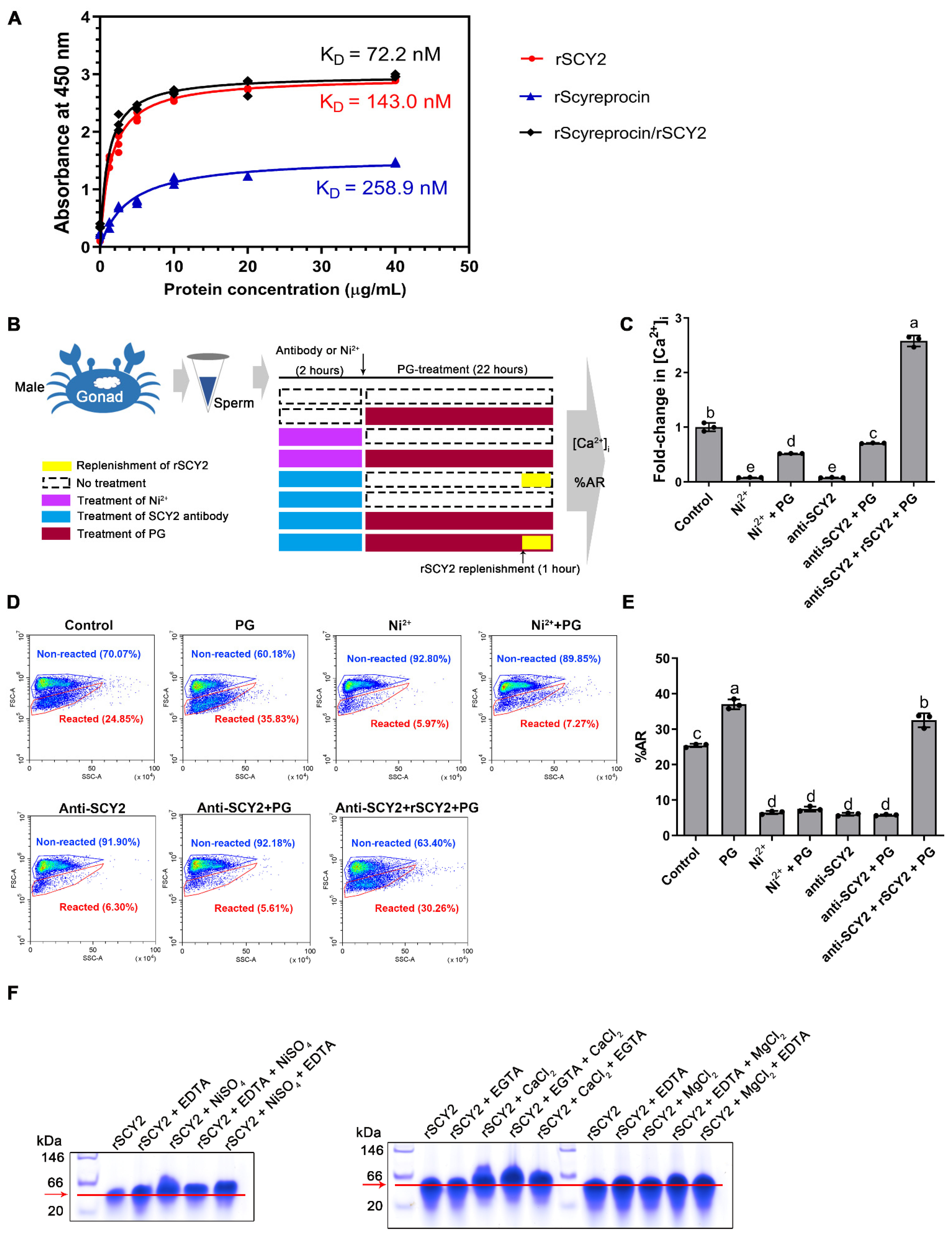
2.7. Progesterone Binding Capacity of SCY2 and Scyreprocin
2.8. SCY2 was Involved in Calcium Influx during Acrosome Reaction
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Isolation of Spermatophores and Seminal Plasma
4.3. Preparation of Recombinant Proteins and Polycolonal Antibodies
4.4. Quantitative PCR
4.5. Antimicrobial Assays
4.6. Western Blotting
4.7. Immunofluorescence Assay
4.8. Hormone Level Examination
4.9. Enzyme-Linked Immunosorbent Assay
4.10. Transmission Electron Microscopy (TEM) Observation
4.11. SCY2-Calcium Binding Property
4.12. Evaluation of Sperm Intracellular Calcium Concentration and Acrosome Reaction Ratio
4.13. In Vitro AR Induction
4.14. Antibody Blockade Assay
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stival, C.; Puga Molina, L.D.C.; Paudel, B.; Buffone, M.; Visconti, P.; Krapf, D. Sperm capacitation and acrosome reaction in mammalian sperm. Sperm Acrosome Biog. Funct. Dur. Fertil. 2016, 220, 93–106. [Google Scholar]
- Dan, J.C. Studies on the acrosome. I. Reaction to egg-water and other stimuli. Biol. Bull. 1952, 103, 54–66. [Google Scholar] [CrossRef]
- Ryuzo, Y.; Gary, C.; Takahiro, M.; Tadashi, A.; Tatsuo, H.; Carol, V.; Murali, P.; Frederick, G.; Hajime, M.; Tina, W. Sperm attractant in the micropyle region of fish and insect eggs. Biol. Reprod. 2013, 88, 47. [Google Scholar] [CrossRef]
- Wilson, K.L.; Fitch, K.R.; Bafus, B.T.; Wakimoto, B.T. Sperm plasma membrane breakdown during Drosophila fertilization requires sneaky, an acrosomal membrane protein. Development 2006, 133, 4871–4879. [Google Scholar] [CrossRef] [Green Version]
- Hirohashi, N.; Yanagimachi, R. Sperm acrosome reaction: Its site and role in fertilization. Biol. Reprod. 2018, 99, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Schackmann, R.W.; Christen, R.; Shapiro, B.M. Membrane potential depolarization and increased intracellular pH accompany the acrosome reaction of sea urchin sperm. Proc. Natl. Acad. Sci. USA 1981, 78, 6066–6070. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.C.; Berkery, D.; Schuel, R.; Laychock, S.G.; Zimmerman, A.M.; Zimmerman, S.; Schuel, H. Evidence for a cannabinoid receptor in sea urchin sperm and its role in blockade of the acrosome reaction. Mol. Reprod. Dev. 1993, 36, 507–516. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Swanson, W.J.; Hellberg, M.E. What have we learned about sea urchin sperm bindin? Dev. Growth Differ. 2003, 37, 1–10. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Moy, G.W. The fucose sulfate polymer of egg jelly binds to sperm REJ and is the inducer of the sea urchin sperm acrosome reaction. Dev. Biol. 1997, 192, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Bleil, J.D.; Wassarman, P.M. Mammalian sperm-egg interaction: Identification of a glycoprotein in mouse egg zonae pellucidae possessing receptor activity for sperm. Cell 1980, 20, 873–882. [Google Scholar] [CrossRef]
- Murphy, S.J.; Yanagimachi, R. The pH dependence of motility and the acrosome reaction of guinea pig spermatozoa. Mol. Reprod. Dev. 1984, 10, 1–8. [Google Scholar] [CrossRef]
- Primakoff, P.; Myles, D.G. Penetration, adhesion, and fusion in mammalian sperm-egg interaction. Science 2002, 296, 2183–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meizel, S.; Turner, K.O. Progesterone acts at the plasma membrane of human sperm. Mol. Cell. Endocrinol. 1991, 77, R1–R5. [Google Scholar] [CrossRef]
- Meizel, S. The sperm, a neuron with a tail: ‘neuronal’ receptors in mammalian sperm. Biol. Rev. 2004, 79, 713–732. [Google Scholar] [CrossRef]
- Dan, J.C. Studies on the Acrosome. III. Effect of Calcium Deficiency. Biol. Bull 1954, 107, 335–349. [Google Scholar] [CrossRef]
- Correia, J.; Michelangeli, F.; Publicover, S. Regulation and roles of Ca2+ stores in human sperm. Reproduction 2015, 150, R65–R76. [Google Scholar] [CrossRef] [Green Version]
- Costello, S.; Michelangeli, F.; Nash, K.; Lefievre, L.; Morris, J.; Machado-Oliveira, G.; Barratt, C.; Kirkman-Brown, J.; Publicover, S. Ca2+-stores in sperm: Their identities and functions. Reproduction 2009, 138, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.R.; Mannowetz, N.; Iavarone, A.T.; Safavi, R.; Gracheva, E.O.; Smith, J.F.; Hill, R.Z.; Bautista, D.M.; Kirichok, Y.; Lishko, P.V. Unconventional endocannabinoid signaling governs sperm activation via the sex hormone progesterone. Science 2016, 352, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Strunker, T.; Goodwin, N.; Brenker, C.; Kashikar, N.D.; Weyand, I.; Seifert, R.; Kaupp, U.B. The CatSper channel mediates progesterone-induced Ca2+ influx in human sperm. Nature 2011, 471, 382–386. [Google Scholar] [CrossRef]
- Seifert, R.; Flick, M.; Bonigk, W.; Alvarez, L.; Trotschel, C.; Poetsch, A.; Muller, A.; Goodwin, N.; Pelzer, P.; Kashikar, N.D.; et al. The CatSper channel controls chemosensation in sea urchin sperm. EMBO J. 2015, 34, 379–392. [Google Scholar] [CrossRef] [Green Version]
- Mcleskey, S.B.; Dowds, C.; Carballada, R.; White, R.R.; Saling, P.M. Molecules involved in mammalian sperm-egg interaction. Int. Rev. Cytol. A Surv. Cell Biol. 1997, 177, 57–113. [Google Scholar] [CrossRef]
- Hirohashi, N.; Kamei, N.; Kubo, H.; Sawada, H.; Matsumoto, M.; Hoshi, M. Egg and sperm recognition systems during fertilization. Dev. Growth Differ. 2008, 50, S221–S238. [Google Scholar] [CrossRef]
- Töpfer-Petersen, E. Molecules on the sperm’s route to fertilization. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2015, 285, 259–266. [Google Scholar] [CrossRef]
- Li, S.J.; Wang, G.Z. Studies on reproductive biology and artificial culture of mud crab, Scylla Serrata. J. Xiamen Univ. 2001, 40, 552–565. [Google Scholar]
- Shang-guan, B.; Li, S. Study on ultrastrcture of the sperm of Scylla serrata (Crustacea, Decapoda, Brachyura). Curr. Zool. 1994, 1, 7–11. [Google Scholar]
- Li, P.; Chan, H.C.; He, B.; So, S.C.; Chung, Y.W.; Shang, Q.; Zhang, Y.D.; Zhang, Y.L. An antimicrobial peptide gene found in the male reproductive system of rats. Science 2001, 291, 1783–1785. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.X.; Zhang, Y.L.; Xiao, L.Q.; Zheng, M.; Leung, K.M.; Chan, M.Y.; Lo, P.S.; Tsang, L.L.; Wong, H.Y.; Ho, L.S. An epididymis-specific β-defensin is important for the initiation of sperm maturation. Nat. Cell Biol. 2004, 6, 458–464. [Google Scholar] [CrossRef]
- Tollner, T.L.; Venners, S.A.; Hollox, E.J.; Yudin, A.I.; Liu, X.; Tang, G.; Xing, H.; Kays, R.J.; Lau, T.; Overstreet, J.W. A Common Mutation in the Defensin DEFB126 Causes Impaired Sperm Function and Subfertility. Sci. Transl. Med. 2011, 3, 92ra65. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Diao, H.; Ni, Z.; Hu, S.G.; Yu, H.G.; Zhang, Y.L. The epididymis-specific antimicrobial peptide β-defensin 15 is required for sperm motility and male fertility in the rat (Rattus norvegicus). Cell. Mol. Life Sci. Cmls 2010, 68, 697–708. [Google Scholar] [CrossRef]
- Qiao, K.; Xu, W.F.; Chen, H.Y.; Peng, H.; Zhang, Y.Q.; Huang, W.S.; Wang, S.P.; An, Z.; Shan, Z.G.; Chen, F.Y.; et al. A new antimicrobial peptide SCY2 identified in Scylla paramamosain exerting a potential role of reproductive immunity. Fish Shellfish Immun. 2016, 51, 251–262. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, F.Y.; Chen, H.Y.; Peng, H.; Hao, H.; Wang, K.J. A novel antimicrobial peptide scyreprocin from mud crab Scylla paramamosain showing potent antifungal and anti-biofilm activity. Front. Microbiol. 2020, 11, 1589. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Li, S.J.; Wang, G.Z. Ultrastructure of the female spermatheca and its storage spermatozoa after spawning of mud crab Scylla serrata. J. Shanghai Fish. Univ. 2000, 9, 69–71. [Google Scholar]
- Ma, X.W.; Hou, L.; Chen, B.; Fan, D.Q.; Chen, Y.C.; Yang, Y.; Wang, K.J. A truncated Sph12–38 with potent antimicrobial activity showing resistance against bacterial challenge in Oryzias melastigma. Fish Shellfish Immun. 2017, 67, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Zhu, K.; Peng, H.; Chen, B.; Liu, J.; Chen, F.; Ma, X.; Wang, S.; Qiao, K.; Wang, K. The new antimicrobial peptide SpHyastatin from the mud crab Scylla paramamosain with multiple antimicrobial mechanisms and high effect on bacterial infection. Front. Microbiol. 2016, 7, 1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brokordt, K.; Defranchi, Y.; Espósito, I.; Cárcamo, C.; Schmitt, P.; Mercado, L.; Erwin, F.O.; Rivera-Ingraham, G.A. Reproduction immunity trade-off in a mollusk: Hemocyte energy metabolism underlies cellular and molecular immune responses. Front. Physiol. 2019, 10, 77. [Google Scholar] [CrossRef] [PubMed]
- Dorin, J.R.; Barratt, C.L.R. Importance of β-defensins in sperm function. Mol. Hum. Reprod. 2014, 20, 821–826. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, O.E.; Gram, L.; Johnsen, A.H.; Andersson, E.; Bangsbøll, S.; Tjabringa, G.S.; Hiemstra, P.S.; Malm, J.; Egesten, A.; Borregaard, N. Processing of seminal plasma hCAP-18 to ALL-38 by gastricsin: A novel mechanism of generating antimicrobial peptides in vagina. J. Biol. Chem. 2003, 278, 28540–28546. [Google Scholar] [CrossRef] [Green Version]
- Eisenbach, M.; Giojalas, L.C. Sperm guidance in mammals-an unpaved road to the egg. Nat. Rev. Mol. Cell Biol. 2006, 7, 276–285. [Google Scholar] [CrossRef]
- Kaupp, U.B.; Kashikar, N.D.; Weyand, I. Mechanisms of sperm chemotaxis. Annu. Rev. Physiol. 2008, 70, 93–117. [Google Scholar] [CrossRef]
- Kekalainen, J.; Evans, J.P. Female-induced remote regulation of sperm physiology may provide opportunities for gamete-level mate choice. Evolution 2017, 71, 238–248. [Google Scholar] [CrossRef]
- Teves, M.E.; Guidobaldi, H.A.; Unates, D.R.; Sanchez, R.; Miska, W.; Publicover, S.J.; Garcia, A.A.M.; Giojalas, L.C. Molecular mechanism for human sperm chemotaxis mediated by progesterone. PLoS ONE 2009, 4, e8211. [Google Scholar] [CrossRef] [Green Version]
- Foresta, C.; Rossato, M.; Mioni, R.; Zorzi, M. Progesterone induces capacitation in human spermatozoa. Andrologia 1992, 24, 33–35. [Google Scholar] [CrossRef]
- Talbot, P.; Summers, R.G.; Hylander, B.L.; Keough, E.M.; Franklin, L.E. The role of calcium in the acrosome reaction: An analysis using ionophore A23187. J. Exp. Zool. 1976, 198, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Du, N.H.; Lai, W.; Xue, L.H. Acrosome reaction of the sperm in the Chinese mitten-handed crab, Eriocheir sinensis (Crustacea, Decapoda). Acta Zool. Sin. 1987, 33, 9–13. [Google Scholar] [CrossRef]
- Clark, W.H.; Griffin, F.J. The morphology and physiology of the acrosome reaction in the sperm of the Decapod, Sicyonia ingentis. Dev. Growth Differ. 1988, 30, 451–462. [Google Scholar] [CrossRef]
- Medina, A.; Rodriguez, A. Structural-changes in sperm from the fiddler crab, Uca tangeri (Crustacea, Brachyura), during the acrosome reaction. Mol. Reprod. Dev. 1992, 33, 195–201. [Google Scholar] [CrossRef]
- Du, N.S.; Lai, W.; An, Y.; Jiang, H.W. Studies on the cytology of fertilization in the Chinese mitten-handed crab, Eriocheir sinensis (Crustacea, Decapoda). Sci. China Ser. B 1993, 36, 288–296. [Google Scholar]
- Ye, H.H.; Song, P.; Ma, J.; Huang, H.Y.; Wang, G.Z. Changes in progesterone levels and distribution of progesterone receptor during vitellogenesis in the female mud crab (Scylla paramamosain). Mar. Freshw. Behav. Physiol. 2010, 43, 25–35. [Google Scholar] [CrossRef]
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nature 2011, 471, 387–391. [Google Scholar] [CrossRef]
- Smith, J.F.; Kirichok, Y.; Lishko, P.V. Calcium channel CatSper is a non-genomic progesterone receptor of human sperm. Biophys. J. 2012, 102, 338a. [Google Scholar] [CrossRef] [Green Version]
- Ralt, D.; Goldenberg, M.; Fetterolf, P.; Thompson, D.; Dor, J.; Mashiach, S.; Garbers, D.L.; Eisenbach, M. Sperm attraction to a follicular factor(s) correlates with human egg fertilizability. Proc. Natl. Acad. Sci. USA 1991, 88, 2840–2844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.M.; Wolfner, M.F. Seminal proteins but not sperm induce morphological changes in the Drosophila melanogaster female reproductive tract during sperm storage. J. Insect Physiol. 2007, 53, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Senarai, T.; Vanichviriyakit, R.; Miyata, S.; Sato, C.; Sretarugsa, P.; Weerachatyanukul, W.; Kitajima, K. Alpha-2 macroglobulin as a region-specific secretory protein in male reproductive tract, and its dynamics during sperm transit towards the female spermatheca in the blue crab. Mol. Reprod. Dev. 2017, 84, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Liu, H.P.; Chen, B.; Hao, H.; Wang, K.J. Optimized production of scygonadin in Pichia pastoris and analysis of its antimicrobial and antiviral activities. Protein Expr. Purif. 2012, 82, 37–44. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford method for protein quantitation. In Basic Protein and Peptide Protocols. Methods in Molecular Biology; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 1994; Volume 32, pp. 9–15. [Google Scholar]
- Xu, W.F.; Qiao, K.; Huang, S.P.; Peng, H.; Huang, W.S.; Chen, B.; Chen, F.Y.; Bo, J.; Wang, K.J. Quantitative gene expression and in situ localization of scygonadin potentially associated with reproductive immunity in tissues of male and female mud crabs, Scylla paramamosain. Fish Shellfish. Immun. 2011, 31, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Arocho, A.; Chen, B.Y.; Ladanyi, M.; Pan, Q.L. Validation of the 2-DDCt calculation as an alternate method of data analysis for quantitative PCR of BCR-ABL P210 transcripts. Diagn. Mol. Pathol. 2006, 15, 56–61. [Google Scholar] [CrossRef]
- Lin, W.; Yang, J.; He, X.; Mo, G.; Jing, H.; Yan, X.; Lin, D.; Ren, L. Structure and function of a potent lipopolysaccharide-binding antimicrobial and anti-inflammatory peptide. J. Med. Chem. 2013, 9, 3546–3556. [Google Scholar] [CrossRef]
- Chen, H.M.; Chan, S.C.; Lee, J.C.; Chang, C.C.; Jack, R.W. Transmission electron microscopic observations of membrane effects of antibiotic Cecropin B on Escherichia coli. Microsc. Res. Tech. 2003, 62, 423–430. [Google Scholar] [CrossRef]
- Lawniczak, M.K.N.; Barnes, A.I.; Linklater, J.R.; Boone, J.M.; Wigby, S.; Chapman, T. Mating and immunity in invertebrates. Trends Ecol. Evol. 2007, 22, 48–55. [Google Scholar] [CrossRef]
- Harshman, L.G.; Zera, A.J. The cost of reproduction: The devil in the details. Trends Ecol. Evol. 2007, 22, 80–86. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | rScyreprocin | rSCY2 | rScyreprocin/rSCY2 | |||
|---|---|---|---|---|---|---|
| MIC a (μM) | MBC a (μM) | MIC (μM) | MBC (μM) | MIC (μM) | MBC (μM) | |
| Pseudomonas putida isolate X1 | <0.5 | 2–4 | 6.25–12.5 | >50 | 0.5–1 | 2–4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Chen, F.; Qiao, K.; Zhang, H.; Chen, H.-Y.; Wang, K.-J. Two Male-Specific Antimicrobial Peptides SCY2 and Scyreprocin as Crucial Molecules Participated in the Sperm Acrosome Reaction of Mud Crab Scylla paramamosain. Int. J. Mol. Sci. 2022, 23, 3373. https://doi.org/10.3390/ijms23063373
Yang Y, Chen F, Qiao K, Zhang H, Chen H-Y, Wang K-J. Two Male-Specific Antimicrobial Peptides SCY2 and Scyreprocin as Crucial Molecules Participated in the Sperm Acrosome Reaction of Mud Crab Scylla paramamosain. International Journal of Molecular Sciences. 2022; 23(6):3373. https://doi.org/10.3390/ijms23063373
Chicago/Turabian StyleYang, Ying, Fangyi Chen, Kun Qiao, Hua Zhang, Hui-Yun Chen, and Ke-Jian Wang. 2022. "Two Male-Specific Antimicrobial Peptides SCY2 and Scyreprocin as Crucial Molecules Participated in the Sperm Acrosome Reaction of Mud Crab Scylla paramamosain" International Journal of Molecular Sciences 23, no. 6: 3373. https://doi.org/10.3390/ijms23063373
APA StyleYang, Y., Chen, F., Qiao, K., Zhang, H., Chen, H.-Y., & Wang, K.-J. (2022). Two Male-Specific Antimicrobial Peptides SCY2 and Scyreprocin as Crucial Molecules Participated in the Sperm Acrosome Reaction of Mud Crab Scylla paramamosain. International Journal of Molecular Sciences, 23(6), 3373. https://doi.org/10.3390/ijms23063373