
Anti-Eryptotic Activity of Food-Derived Phytochemicals and Natural Compounds

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
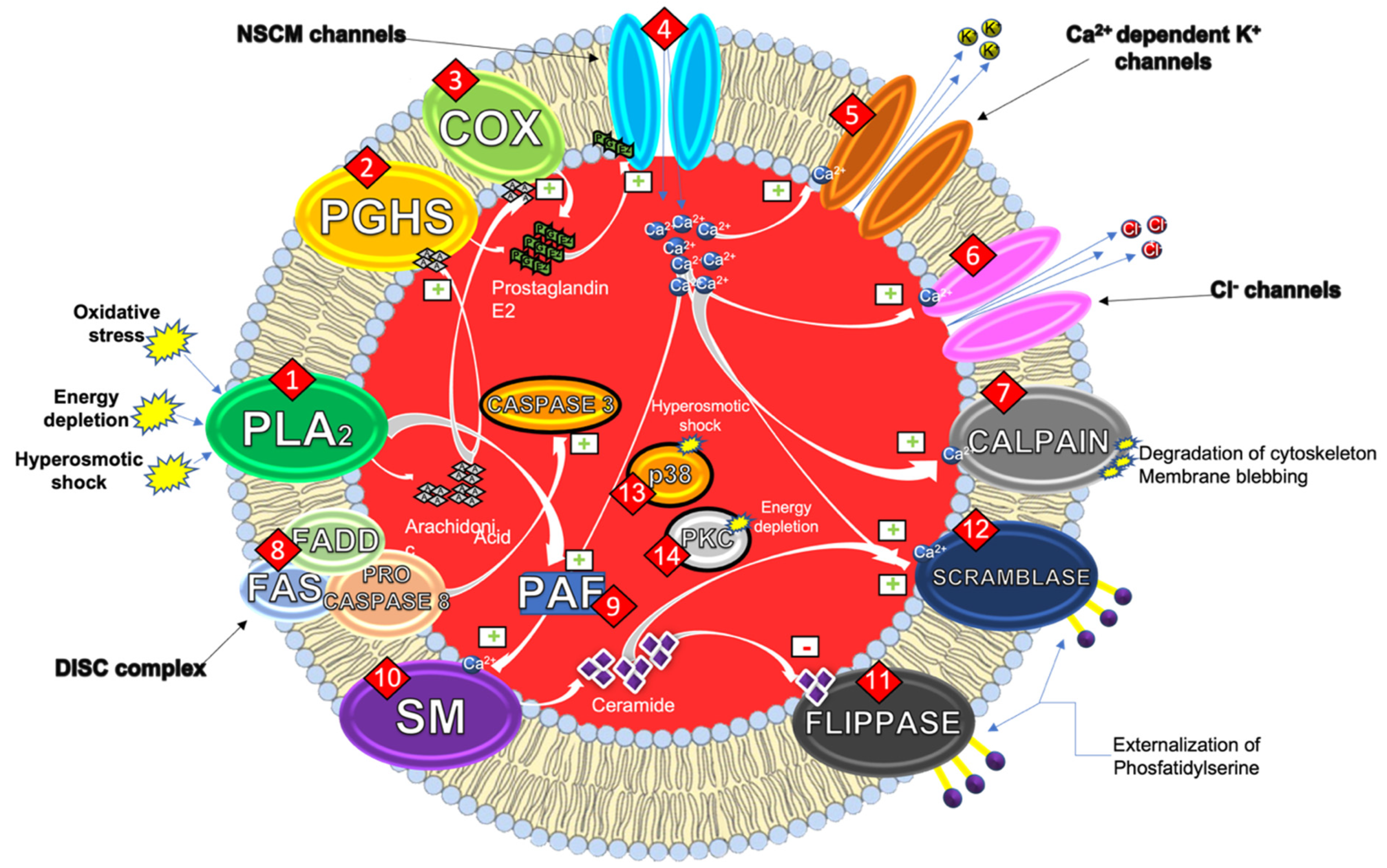
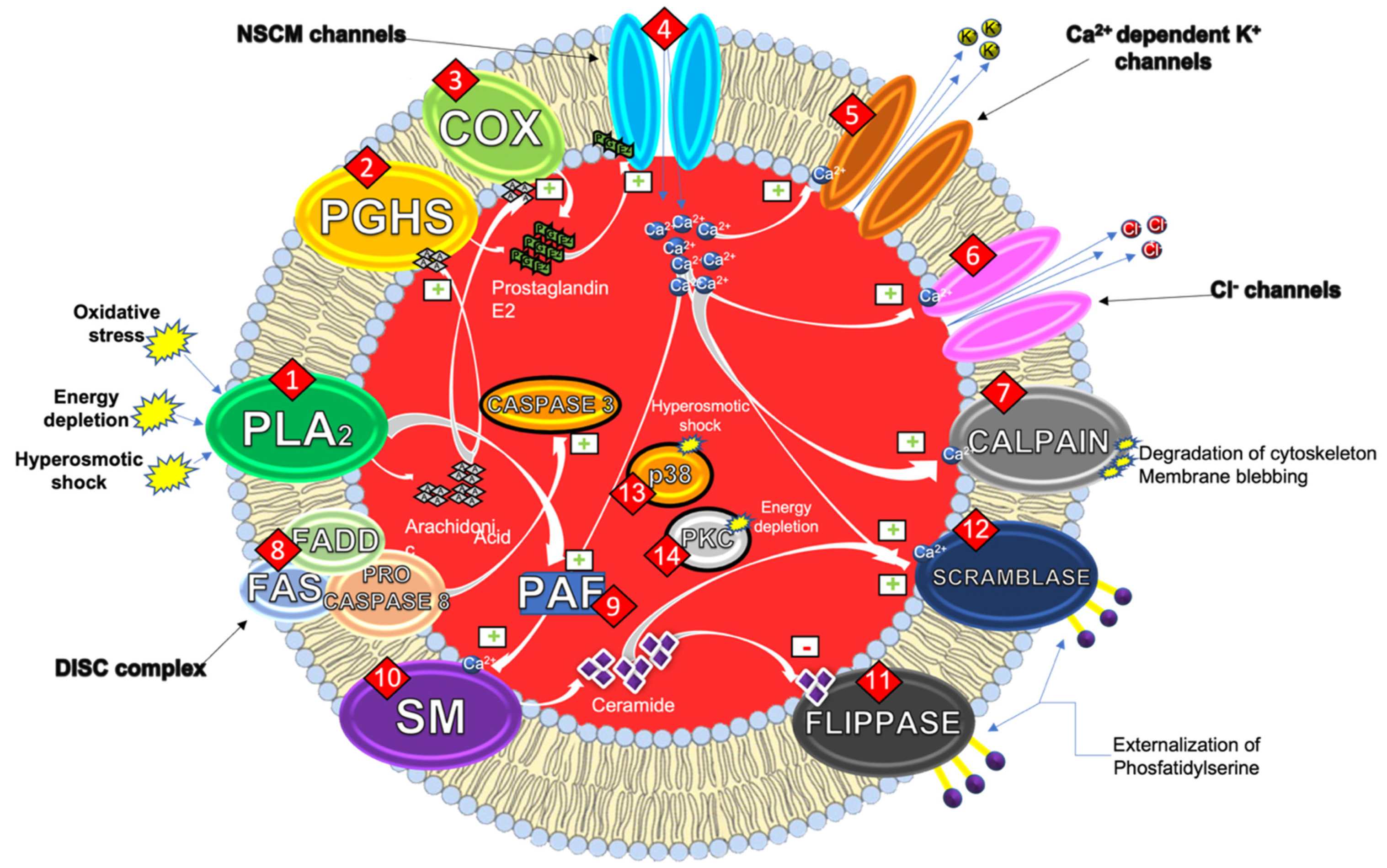
:1. Eryptosis
1.1. Mechanisms of the Eryptotic Machinery
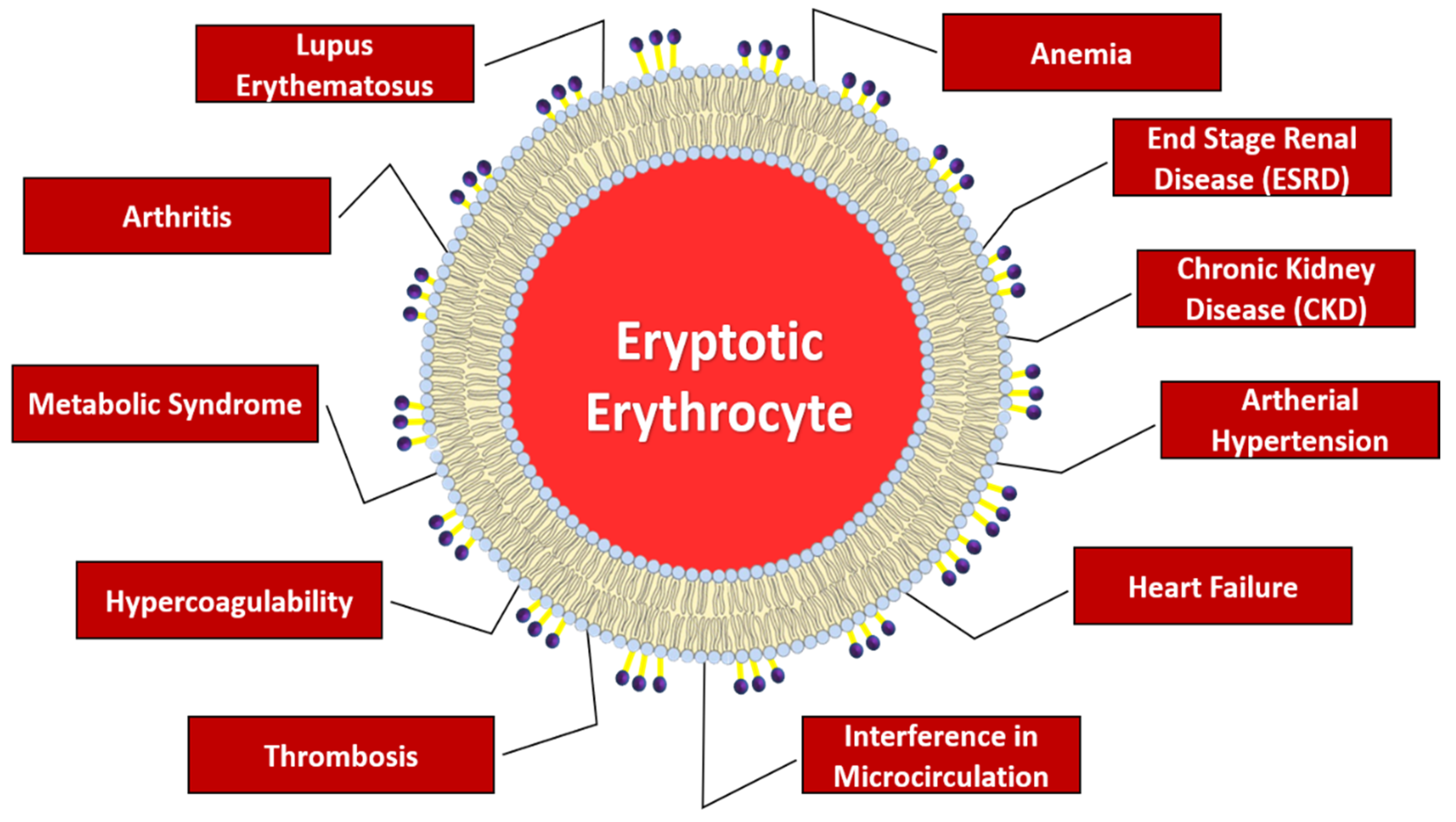
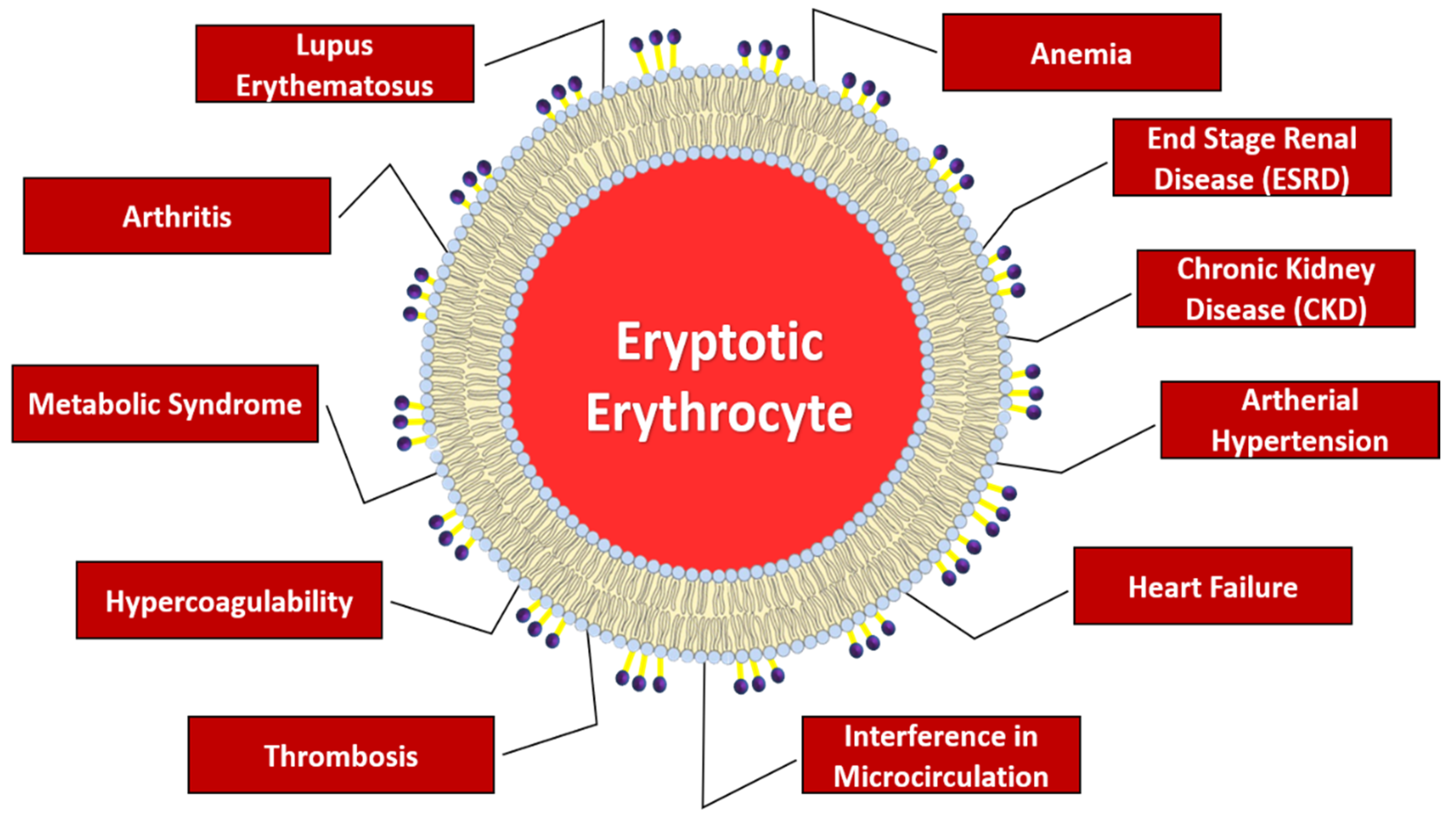
1.2. Pathophysiological Implications
2. Natural Substances with Anti-Eryptotic Activity
2.1. Phenolic Compounds
2.2. Alkaloids Compounds
2.3. Other Natural Compounds
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Arese, P.; Turrini, F.; Schwarzer, E. Band 3/complement-mediated recognition and removal of normally senescent and pathological human erythrocytes. Cell. Physiol. Biochem. 2005, 16, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.; Qadri, S.M.; Lang, F. Killing me softly—Suicidal erythrocyte death. Int. J. Biochem. Cell. Biol. 2012, 44, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Föller, M.; Huber, S.M.; Lang, F. Erythrocyte programmed cell death. IUBMB Life 2008, 60, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Qadri, S.M. Mechanisms and significance of eryptosis, the suicidal death of erythrocytes. Blood Purif. 2012, 33, 125–130. [Google Scholar] [CrossRef]
- Elliott, S.; Pham, E.; Macdougall, I.C. Erythropoietins: A common mechanism of action. Exp. Hematol. 2008, 36, 1573–1584. [Google Scholar] [CrossRef]
- Restivo, I.; Attanzio, A.; Tesoriere, L.; Allegra, M. Suicidal erythrocyte death in metabolic Syndrome. Antioxidants 2021, 10, 154. [Google Scholar] [CrossRef]
- Repsold, L.; Joubert, A.M. Eryptosis: An erythrocyte’s suicidal type of cell death. Biomed. Res. Int. 2018, 2018, 9405617. [Google Scholar] [CrossRef]
- Maher, A.D.; Kuchel, P.W. The Gárdos channel: A review of the Ca2+-activated K+ channel in human erythrocytes. Int. J. Biochem. Cell Biol. 2003, 35, 1182–1197. [Google Scholar] [CrossRef]
- Velásquez, F.C.; Maté, S.; Bakás, L.; Herlax, V. Induction of eryptosis by low concentrations of E. coli alpha-hemolysin. Biochim. Biophys. Acta 2015, 1848, 2779–2788. [Google Scholar] [CrossRef] [Green Version]
- Berg, C.P.; Engels, I.H.; Rothbart, A.; Lauber, K.; Renz, A.; Schlosser, S.F.; Schulze-Osthoff, K.; Wesselborg, S. Human mature red blood cells express caspase-3 and caspase-8, but are devoid of mitochondrial regulators of apoptosis. Cell Death Differ. 2001, 8, 1197–1206. [Google Scholar] [CrossRef]
- Mandal, D.; Mazumder, A.; Das, P.; Kundu, M.; Basu, J. Fas-, caspase 8-, and caspase 3-dependent signaling regulates the activity of the aminophospholipid translocase and phosphatidylserine externalization in human erythrocytes. J. Biol. Chem. 2005, 280, 39460–39467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, P.A.; Kempe, D.S.; Tanneur, V.; Eisele, K.; Klarl, B.A.; Myssina, S.; Jendrossek, V.; Ishii, S.; Shimizu, T.; Waidmann, M.; et al. Stimulation of erythrocyte ceramide formation by platelet-activating factor. J. Cell Sci. 2005, 118, 1233–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, F.; Gulbins, E.; Lang, P.A.; Zappulla, D.; Föller, M. Ceramide in suicidal death of erythrocytes. Cell. Physiol. Biochem. 2010, 26, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Gatidis, S.; Zelenak, C.; Fajol, A.; Lang, E.; Jilani, K.; Michael, D.; Qadri, S.M.; Lang, F. p38 MAPK activation and function following osmotic shock of erythrocytes. Cell. Physiol. Biochem. 2011, 28, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Abed, M.; Lang, E.; Foller, M. Oxidative stress and suicidal erythrocyte death. Antioxid. Redox Signal. 2014, 21, 138–153. [Google Scholar] [CrossRef]
- Klarl, B.A.; Lang, P.A.; Kempe, D.S.; Niemoeller, O.M.; Akel, A.; Sobiesiak, M.; Eisele, K.; Podolski, M.; Huber, S.M.; Wieder, T.; et al. Protein kinase C mediates erythrocyte “programmed cell death” following glucose depletion. Am. J. Physiol. Cell Physiol. 2006, 290, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Boas, F.E.; Forman, L.; Beutler, E. Phosphatidylserine exposure and red cell viability in red cell aging and in hemolytic anemia. Proc. Natl. Acad. Sci. USA 1998, 95, 3077–3081. [Google Scholar] [CrossRef] [Green Version]
- Qadri, S.M.; Bissinger, R.; Solh, Z.; Oldenborg, P.A. Eryptosis in health and disease: A paradigm shift towards understanding the (patho)physiological Implications of programmed cell death of erythrocytes. Blood Rev. 2017, 31, 349–361. [Google Scholar] [CrossRef]
- Artunc, F.; Risler, T. Serum erythropoietin concentrations and responses to anemia in patients with or without chronic kidney disease. Nephrol. Dial. Transpl. 2007, 22, 2900–2908. [Google Scholar] [CrossRef] [Green Version]
- Attanasio, P.; Ronco, C.; Anker, S.D.; Cicoira, M.; von Haehling, S. Role of iron deficiency and anemia in cardiorenal syndromes. Semin. Nephrol. 2012, 32, 57–62. [Google Scholar] [CrossRef]
- Lang, F.; Bissinger, R.; Abed, M.; Artunc, F. Eryptosis—The neglected cause of anemia in end stage renal disease. Kidney Blood Press Res. 2017, 42, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Abed, M.; Artunc, F.; Alzoubi, K.; Honisch, S.; Baumann, D.; Foller, M.; Lang, F. Suicidal erythrocyte death in end stage renal disease. J. Mol. Med. 2014, 92, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, M.; Del Vecchio, L.; Sirolli, V.; Locatelli, F. New treatment approaches for the anemia of CKD. Am. J. Kidney Dis. 2016, 67, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Marks, P.W. Hematologic manifestations of liver disease. Semin. Hematol. 2013, 50, 216–221. [Google Scholar] [CrossRef]
- Maheshwari, A.; Mishra, R.; Thuluvath, P.J. Post-liver-transplant anemia: Etiology and management. Liver Transpl. 2004, 10, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.; Gatidis, S.; Freise, N.F.; Bock, H.; Kubitz, R.; Lauermann, C.; Orth, H.M.; Klindt, C.; Schuier, M.; Keitel, V.; et al. Conjugated bilirubin triggers anemia by inducing erythrocyte death. Hepatology 2015, 61, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Deray, G.; Heurtier, A.; Grimaldi, A.; Launay Vacher, V.; Isnard Bagnis, C. Anemia and diabetes. Am. J. Nephrol. 2004, 24, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Gauci, R.; Hunter, M.; Bruce, D.G.; Davis, W.A.; Davis, T.M.E. Anemia complicating type 2 diabetes: Prevalence, risk factors and prognosis. J. Diabetes Complic. 2017, 31, 1169–1174. [Google Scholar] [CrossRef]
- Singh, D.K.; Winocour, P.; Farrington, K. Erythropoietic stress and anemia in diabetes mellitus. Nat. Rev. Endocrinol. 2009, 5, 204–210. [Google Scholar] [CrossRef]
- Calderón-Salinas, J.V.; Muñoz-Reyes, E.G.; Guerrero-Romero, J.F.; Rodríguez-Morán, M.; Bracho-Riquelme, R.L.; Carrera-Gracia, M.A.; Quintanar-Escorza, M.A. Eryptosis and oxidative damage in type 2 diabetic mellitus patients with chronic kidney disease. Mol. Cell. Biochem. 2011, 357, 171–179. [Google Scholar] [CrossRef]
- Maellaro, E.; Leoncini, S.; Moretti, D.; Del Bello, B.; Tanganelli, I.; De Felice, C.; Ciccoli, L. Erythrocyte caspase-3 activation and oxidative imbalance in erythrocytes and in plasma of type 2 diabetic patients. Acta Diabetol. 2013, 50, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Vlassara, H.; Uribarri, J. Advanced glycation end products (AGE) and diabetes: Cause, effect, or both? Curr. Diabetes Rep. 2014, 14, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinzón-Díaz, C.E.; Calderón-Salinas, J.V.; Rosas-Flores, M.M.; Hernández, G.; López-Betancourt, A.; Quintanar-Escorza, M.A. Eryptosis and oxidative damage in hypertensive and dyslipidemic patients. Mol. Cell. Biochem. 2018, 440, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Attanasio, P.; Bissinger, R.; Haverkamp, W.; Pieske, B.; Wutzler, A.; Lang, F. Enhanced suicidal erythrocyte death in acute cardiac failure. Eur. J. Clin. Investig. 2015, 45, 1316–1324. [Google Scholar] [CrossRef]
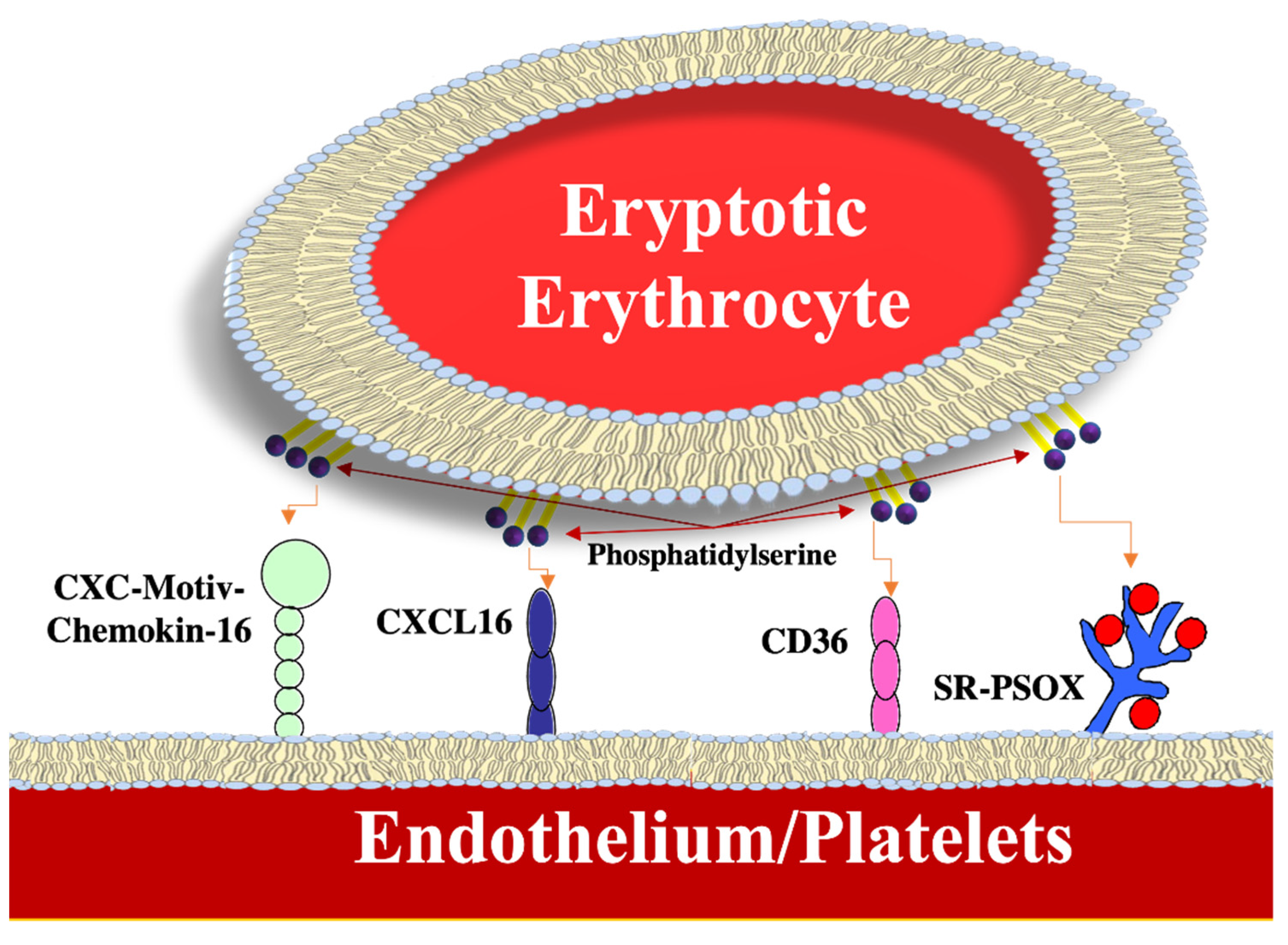
- Borst, O.; Abed, M.; Alesutan, I.; Towhid, S.T.; Qadri, S.M.; Föller, M.; Gawaz, M.; Lang, F. Dynamic adhesion of eryptotic erythrocytes to endothelial cells via CXCL16/SR-PSOX. Am. J. Physiol. Cell Physiol. 2012, 302, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.; Towhid, S.T.; Schmid, E.; Hoffmann, S.M.; Abed, M.; Münzer, P.; Vogel, S.; Neis, F.; Brucker, S.; Gawaz, M.; et al. Dynamic adhesion of eryptotic erythrocytes to immobilized platelets via platelet phosphatidylserine receptors. Am. J. Physiol. Cell Physiol. 2014, 306, 291–297. [Google Scholar] [CrossRef]
- Barr, J.D.; Chauhan, A.K.; Schaeffer, G.V.; Hansen, J.K.; Motto, D.G. Red blood cells mediate the onset of thrombosis in the ferric chloride murine model. Blood 2013, 121, 3733–3741. [Google Scholar] [CrossRef]
- Qadri, S.M.; Donkor, D.A.; Bhakta, V.; Eltringham-Smith, L.J.; Dwivedi, D.J.; Moore, J.C.; Pepler, L.; Ivetic, N.; Nazi, I.; Fox-Robichaud, A.E.; et al. Phosphatidylserine externalization and procoagulant activation of erythrocytes induced by Pseudomonas aeruginosa virulence factor pyocyanin. J. Cell. Mol. Med. 2016, 20, 710–720. [Google Scholar] [CrossRef] [Green Version]
- Attanzio, A.; Frazzitta, A.; Vasto, S.; Tesoriere, L.; Pintaudi, A.M.; Livrea, M.A.; Cilla, A.; Allegra, M. Increased eryptosis in smokers is associated with the antioxidant status and C-reactive protein levels. Toxicology 2019, 411, 43–48. [Google Scholar] [CrossRef]
- Solá, E.; Vayá, A.; Martínez, M.; Moscardó, A.; Corella, D.; Santaolaria, M.L.; España, F.; Hernández-Mijares, A. Erythrocyte membrane phosphatidylserine exposure in obesity. Obesity 2009, 17, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; López-García, G.; Collado-Díaz, V.; Amparo Blanch-Ruiz, M.; Garcia-Llatas, G.; Barberá, R.; Martinez-Cuesta, M.A.; Real, J.T.; Álvarez, Á.; Martínez-Hervás, S. Hypercholesterolemic patients have higher eryptosis and erythrocyte adhesion to human endothelium independently of statin therapy. Int. J. Clin. Pract. 2021, 75, e14771. [Google Scholar] [CrossRef] [PubMed]
- Alfhili, M.A.; Aljuraiban, G.S. Lauric acid, a dietary saturated medium-chain fatty acid, elicits calcium-dependent eryptosis. Cells 2021, 10, 3388. [Google Scholar] [CrossRef] [PubMed]
- Olumuyiwa-Akeredolu, O.O.; Pretorius, E. Platelet and red blood cell interactions and their role in rheumatoid arthritis. Rheumatol. Int. 2015, 35, 1955–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, P.; Bian, M.; Ma, W.; Liu, C.; Yang, P.; Zhu, B.; Xu, Y.; Zheng, M.; Qiao, J.; Shuai, Z.; et al. Eryptosis as an underlying mechanism in systemic lupus erythematosus-related anemia. Cell. Physiol. Biochem. 2016, 40, 1391–1400. [Google Scholar] [CrossRef]
- Lang, E.; Bissinger, R.; Qadri, S.M.; Lang, F. Suicidal death of erythrocytes in cancer and its chemotherapy: A potential target in the treatment of tumor-associated anemia. Int. J. Cancer 2017, 141, 1522. [Google Scholar] [CrossRef] [Green Version]
- Cilla, A.; Alegría, A.; Attanzio, A.; Garcia-Llatas, G.; Tesoriere, L.; Livrea, M.A. Dietary phytochemicals in the protection against oxysterol-induced damage. Chem. Phys. Lipids 2017, 207 Pt B, 192–205. [Google Scholar] [CrossRef]
- Yu, M.; Gouvinhas, I.; Rocha, J.; Barros, A. Phytochemical and antioxidant analysis of medicinal and food plants towards bioactive food and pharmaceutical resources. Sci. Rep. 2021, 11, 10041. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the Phenol-Explorer database. Eur. J. Clin. Nutr. 2010, 64 (Suppl. 3), S112–S120. [Google Scholar] [CrossRef]
- Albuquerque, B.R.; Heleno, S.A.; Oliveira, M.; Barros, L.; Ferreira, I. Phenolic compounds: Current industrial applications, limitations and future challenges. Food. Funct. 2021, 12, 14–29. [Google Scholar] [CrossRef]
- Colizzi, C. The protective effects of polyphenols on Alzheimer’s disease. Alzheimers Dement. 2018, 15, 184–196. [Google Scholar]
- Alfhili, M.A.; Basudan, A.M.; Alsughayyir, J. Antiproliferative Wnt inhibitor wogonin prevents eryptosis following ionophoric challenge, hyperosmotic shock, oxidative stress, and metabolic deprivation. J. Food Biochem. 2021, 45, e13977. [Google Scholar] [CrossRef] [PubMed]
- Delveaux, J.; Turpin, C.; Veeren, B.; Diotel, N.; Bravo, S.B.; Begue, F.; Álvarez, E.; Meilhac, O.; Bourdon, E.; Rondeau, P. Antirhea borbonica aqueous extract protects albumin and erythrocytes from glycoxidative damages. Antioxidants 2020, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Tortora, F.; Notariale, R.; Lang, F.; Manna, C. Hydroxytyrosol decreases phosphatidylserine exposure and inhibits suicidal death induced by lysophosphatidic acid in human erythrocytes. Cell Physiol. Biochem. 2019, 53, 921–932. [Google Scholar]
- Officioso, A.; Alzoubi, K.; Lang, F.; Manna, C. Hydroxytyrosol inhibits phosphatidylserine exposure and suicidal death induced by mercury in human erythrocytes: Possible involvement of the glutathione pathway. Food Chem. Toxicol. 2016, 89, 47–53. [Google Scholar] [CrossRef]
- Liu, J.; Bhuyan, A.; Ma, K.; Zhang, S.; Cheng, A.; Lang, F. Inhibition of suicidal erythrocyte death by pyrogallol. Mol. Bio. Rep. 2020, 47, 5025–5032. [Google Scholar] [CrossRef]
- Singh, S.; Garg, G.; Singh, A.K.; Bissoyi, A.; Rizvi, S.I. Fisetin, a potential caloric restriction mimetic, attenuates senescence biomarkers in rat erythrocytes. Biochem. Cell Biol. 2019, 97, 480–487. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M.; Tufarelli, V. In vitro antioxidant activities of resveratrol, cinnamaldehyde and their synergistic effect against cyadox-induced cytotoxicity in rabbit erythrocytes. Drug. Chem. Toxicol. 2017, 40, 196–205. [Google Scholar] [CrossRef]
- Qadri, S.M.; Föller, M.; Lang, F. Inhibition of suicidal erythrocyte death by resveratrol. Life Sci. 2009, 85, 33–38. [Google Scholar] [CrossRef]
- Shaik, N.; Zbidah, M.; Lang, F. Inhibition of Ca2+ entry and suicidal erythrocyte death by naringin. Cell. Physiol. Biochem. 2009, 30, 678–686. [Google Scholar] [CrossRef]
- Gatidis, S.; Meier, A.; Jilani, K.; Lang, E.; Zelenak, C.; Qadri, S.M.; Lang, F. Phlorhizin protects against erythrocyte cell membrane scrambling. J. Agric. Food Chem. 2011, 59, 8524–8530. [Google Scholar] [CrossRef] [PubMed]
- Qadri, S.M.; Mahmud, H.; Föller, M.; Lang, F. Inhibition of suicidal erythrocyte death by xanthohumol. J. Agric. Food Chem. 2009, 57, 7591–7595. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, H.; Mauro, D.; Foller, M.; Lang, F. Inhibitory effect of thymol on suicidal erythrocyte death. Cell. Physiol. Biochem. 2009, 24, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; du Plooy, J.N.; Bester, J. A Comprehensive Review on Eryptosis. Cell. Physiol. Biochem. 2016, 39, 1977–2000. [Google Scholar] [CrossRef]
- Lang, E.; Lang, F. Triggers, inhibitors, mechanisms, and significance of eryptosis: The suicidal erythrocyte death. Biomed. Res. Int. 2015, 2015, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.; Sun, H.; Zhang, A.H.; Xu, H.Y.; Yan, G.L.; Han, Y.; Wang, X.J. Natural alkaloids: Basic aspects, biological roles, and future perspectives. Chin. J. Nat. Med. 2014, 12, 401–406. [Google Scholar] [CrossRef]
- Dang, T.T.; Onoyovwi, A.; Farrow, S.C.; Facchini, P.J. Biochemical genomics for gene discovery in benzylisoquinoline alkaloid biosynthesis in opium poppy and related species. Methods Enzymol. 2012, 515, 231–266. [Google Scholar]
- Floride, E.; Föller, M.; Ritter, M.; Lang, F. Caffeine inhibits suicidal erythrocyte death. Cell. Physiol. Biochem. 2008, 22, 253–260. [Google Scholar] [CrossRef]
- Ghashghaeinia, M.; Koralkova, P.; Giustarini, D.; Mojzikova, R.; Fehrenbacher, B.; Dreischer, P.; Schaller, M.; Mrowietz, U.; Martínez-Ruiz, A.; Wieder, T.; et al. The specific PKC-α inhibitor chelerythrine blunts costunolide-induced eryptosis. Apoptosis 2020, 25, 674–685. [Google Scholar] [CrossRef]
- Tesoriere, L.; Attanzio, A.; Allegra, M.; Livrea, M.A. Dietary indicaxanthin from cactus pear (Opuntia ficusindica L. Mill) fruit prevents eryptosis induced by oxysterols in a hypercholesterolaemia-relevant proportion and adhesion of human erythrocytes to endothelial cell layers. Br. J. Nutr. 2015, 114, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Allegra, M.; Restivo, I.; Fucarino, A.; Pitruzzella, A.; Vasto, S.; Livrea, M.A.; Tesoriere, L.; Attanzio, A. Proeryptotic activity of 4-hydroxynonenal: A new potential physiopathological role for lipid peroxidation products. Biomolecules 2020, 10, 770. [Google Scholar] [CrossRef] [PubMed]
- Kengaiah, J.; Nandish, S.; Ramachandraiah, C.; Chandramma; Shivaiah, A.; Vishalakshi, G.J.; Paul, M.; Santhosh, M.S.; Shankar, R.L.; Sannaningaiah, D. Protective Effect of Tamarind Seed Coat Ethanol Extract on Eryptosis Induced by Oxidative Stress. Biochemistry 2020, 85, 119–129. [Google Scholar] [PubMed]
- Alvarez-Sala, A.; López-García, G.; Attanzio, A.; Tesoriere, L.; Cilla, A.; Barberá, R.; Alegría, A. Effects of plant sterols or β-cryptoxanthin at physiological serum concentrations on suicidal erythrocyte death. J. Agric. Food. Chem. 2018, 66, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, G.; Li, X.; Shi, Y.; Guan, G. L-Carnitine inhibits eryptosis induced by uremic serum and the related mechanisms. Ren. Fail. 2015, 37, 1050–1056. [Google Scholar] [CrossRef] [Green Version]
- Qian, E.W.; Ge, D.T.; Kong, S.K. Salidroside protects human erythrocytes against hydrogen peroxide-induced apoptosis. J. Nat. Prod. 2012, 75, 531–537. [Google Scholar] [CrossRef]
- Shan, F.; Yang, R.; Ji, T.; Jiao, F. Vitamin C inhibits aggravated eryptosis by hydrogen peroxide in glucose-6-phosphated dehydrogenase deficiency. Cell. Physiol. Biochem. 2016, 39, 1453–1462. [Google Scholar] [CrossRef]
- Mahmud, H.; Qadri, S.M.; Föller, M.; Lang, F. Inhibition of suicidal erythrocyte death by vitamin C. Nutrition 2010, 26, 671–676. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Phytochemical Compound | Concentration | Treatment Time | Inductor | References |
|---|---|---|---|---|
| Wogonin (WGN) | 100 µM | 24–48 h | Energy depletion—48 h Cell shrinkage—24 h Ionomycin 2 µM—24 h PEITC 134 mM—24 h | [52] |
| Cellular mechanism At 48 h with WGN, in presence of energy depletion, eryptosis inhibited with PS↓, Ca2+↓ and FS↑. At 24 h in presence of oxidative stress inducted by PEITC and hyperosmotic conditions, eryptosis inhibited with PS↓ and in presence of ionophore ionomycin for 24 h eryptosis inhibited with PS↓ and Ca2+↓ | ||||
| Antirhea Borbonica Extract (ABE) | 500 mg /L | 24–48 h in zebrafish | MGO0–10 mM 24 h | [53] |
| Cellular mechanism At 48 h with ABE eryptosis inhibited with PS↓ and ROS↓. Antiglycative and antioxidant action on albumin with 24 h of treatment | ||||
| Hydroxytyrosol (HT) | 0.1–1 µM [54] 1–5 µM [55] | 4 h [54]–24 h [55] | LPA 0.5–2.5 µM—24 h [54] HgCl2 2.5 µM—4 h [55] | [54] |
| [55] | ||||
| Cellular mechanism At 24 h with HT 0.1 µM, eryptosis inhibited with PS↓, Ca2+↓ and GSH↑. Moreover, FS↑ and ATP↑ with HT 0.5 µM and MV↓ with HT 1 µM [55]. At 4 h with HT 1 µM, eryptosis inhibited with PS↓, GSH↑ and ATP↑ [54] | ||||
| Pyrogallol (PYR) | 2–8 µM | 30 min–48 h | Energy depletion—48 h | [56] |
| Cell shrinkage—6 h | ||||
| Ionomycin 1 µM—1 h | ||||
| tBOOH 0.3 mM—30 min | ||||
| Cellular mechanism Eryptosis inhibited with PS↓ with PYR 2 µM at 48 h in presence of energy depletion, at 6 h in presence of hyperosmotic environment, at 1 h in presence of ionophore ionomycin, and also after a treatment for 30 min in presence of oxidative stress inducted by tBOOH | ||||
| Fisetin (FIS) | 15 mg/kg | 6 weeks in rats | D-galactose 500 mg/kg 4 months | [57] |
| Cellular mechanism Eryptosis inhibited with PS↓, ROS↓, GSH↑ | ||||
| Resveratrol (RES) | 1–40 µM | In rabbit 3 h [58] | Cyadox 40 µg/mL—3 h [58] | [58] |
| In human 30 min– 48 h [59] | tBOOH 0.3 mM 30 min [59] | [59] | ||
| Energy depletion—48 h [59] | ||||
| Cell shrinkage—48 h [59] | ||||
| Cellular mechanism In rabbit’s RBCs, at 3 h with RES 40 µM, eryptosis inhibited with oxidative stress↓, activity of active caspases 3 and 8↓ and LDH↓ [58]. Furthermore, in human’s RBCs at 30 min with RES 10 µM, in presence of oxidative stress inducted by tBOOH, PS↓; at 48 h in presence of inducted cell shrinkage with RES 1 µM PS↓ and at 48 h energy depletion PS↓ and Ca2+↓ with RES 5 µM and FS↑ with RES 10 µM [59] | ||||
| Naringin (NAR) | 10–40 µM | 30 min–48 h | Energy depletion—48 h | [60] |
| tBOOH 0.3 mM—30 min | ||||
| Cellular mechanism At 48 h with NAR, in presence of energy depletion, from 10 µM eryptosis inhibited with PS↓, from 20 µM Ca2+↓, and from 40 µM FS↑. At 30 min with NAR, in presence of oxidative stress inducted by tBOOH, eryptosis from 40 µM PS↓ and FS↑ | ||||
| Phlorizin (PHL) | 10–100 µM | 30 min–48 h | Energy depletion—48 h | [61] |
| tBOOH 0.3 mM—30 min | ||||
| Cellular mechanism At 48 h with PHL 50 µM, in presence of energy depletion, eryptosis inhibited with PS↓ and FS↑. At 30 min with PHL 10 µM, in presence of oxidative stress inducted by tBOOH, eryptosis inhibited with PS↓ and with 50 µM FS↑ and Ca2+↓ | ||||
| Xanthohumol (XAN) | 0.25–1 µM | 30 min–48 h | Energy depletion—48 h | [62] |
| tBOOH 0.3 mM—30 min | ||||
| Cellular mechanism At 48 h with XAN, in presence of energy depletion, eryptosis inhibited with PS↓ with XAN 0.25 µM and Ca2+↓ and FS↑ with XAN 0.5 µM. At 30 min, in presence of oxidative stress inducted by tBOOH, eryptosis inhibited with PS↓ with XAN 0.5 µM and Ca2+↓ with XAN 1 µM | ||||
| Thymol (THY) | 2.5–20 µg/mL | 30 min–48 h | Energy depletion—48 h | [63] |
| Cell shrinkage—48 h | ||||
| tBOOH 0.3 mM—30 min | ||||
| Cellular mechanism At 48 h with THY 20 µg/mL, in presence of energy depletion, eryptosis inhibited with PS↓ and FS↑; with the same conditions but in presence of cell shrinkage THY inhibit eryptosis only with PS↓. At 30 min, in presence of oxidative stress inducted by tBOOH in presence or absence of Ca2+ in the ringer, eryptosis inhibited with PS↓ and Ca2+↓ with THY 2.5 µg/mL | ||||
| Phytochemical Compound | Concentration | Treatment Time | Inductor | References |
|---|---|---|---|---|
| Caffeine (CAF) | 50–500 µM | 48 h | Energy depletion—48 h | [68] |
| Cell shrinkage—48 h | ||||
| Cellular mechanism After 48 h of treatment with CAF 500 µM, in presence of energy depletion, eryptosis inhibited with PS↓, Ca2+↓ and FS↑. After 48 h of treatment with CAF 50 µM, in presence of cell shrinkage, eryptosis inhibited only with PS↓ | ||||
| Chelerythrine (CHE) | 1–10 µM | 24 h | Custonolide 1–80 µM—24 h | [69] |
| Cellular mechanism At 24 h with CHE 10 µM, in presence of custonolide 80 µM, eryptosis inhibited with PS↓ and FS↑ | ||||
| Indicaxanthin (IND) | 1–5 µM | 48 h | Mixture of oxysterols | [70] |
| 20 mM—48 h | ||||
| Cellular mechanism At 48 h with IND 1 µM, in presence of mixture of oxysterols 20 mM, eryptosis inhibited with PS↓, Ca2+↓, FS↑, ROS↓ and PGE2↓. Moreover, at 48 h of with IND 2.5 µM GSH↑ and endothelial adherence↓ with IND 5 µM. | ||||
| Phytochemical Compound | Concentration | Treatment Time | Inductor | References |
|---|---|---|---|---|
| Acetylsalycilic acid (ASA) | 50 µM | 24 h | 4-Hydroxynonenal | [71] |
| 25–50 µM—24 h | ||||
| Cellular mechanism Eryptosis inhibited with PS↓ and Ca2+↓ | ||||
| Tamarind seed | 50–200 µg/mL | 24 h | AAPH 100–1000 µM—24 h | [72] |
| Extract (TSCEE) | ||||
| Cellular mechanism In presence of oxidative stress inducted by AAPH, eryptosis inhibited from TSCEE 50 µg/mL with PS↓, Ca2+↓, ROS↓ and GSH↑ | ||||
| Plant sterols (MPS) | 22 µM | 48 h | tBOOH 0.075 or 0.3 mM 30 min | [73] |
| Cellular mechanism Eryptosis inhibited with Ca2+↓, Hemolysis↓, ROS↓ and GSH↑ in presence of tBOOH 75 µM. | ||||
| L-Carnitine (LCar) | 200 umol/L | 24–48 h | 30% uremic serum (v/v) | [74] |
| 24–48 h | ||||
| Cellular mechanism Eryptosis inhibited with PS↓, ROS↓ and GSH↑ | ||||
| Salidroside (SAL) | 100–300 µM | 24 h | H2O2 1 mM—24 h | [75] |
| Cellular mechanism Eryptosis inhibited from SAL 100 µM with PS↓, Ca2+↓, ROS↓, eemolysis↓ and activity of active Caspase 3↓ | ||||
| Vitamin C (VitC) | 60–280 µM | 30 min [77] | H2O2 0.5% (v/v)—30 min [76] | [76] |
| 30 min–48 h [76] | G6PD deficiency [76] | [77] | ||
| Energy depletion—48 h [77] | ||||
| Cell shrinkage—48 h [77] | ||||
| tBOOH 0.3 mM—30 min [77] | ||||
| Cellular mechanism In patients with G6PD deficiency, at 30 min with VitC 60 µM in presence or absence of H2O2 0.5% (v/v), eryptosis inhibited with PS↓ and activity of active caspases 3↓ [76]. At 48 h with VitC, in presence of energy depletion, eryptosis inhibited with PS↓, Ca2+↓, and FS↑ from VitC 110 µM; with the same treatment but with cell shrinkage, VitC 60 µM inhibit eryptosis with PS↓ and at 30 min, in presence of oxidative stress inducted by tBOOH, eryptosis inhibited with PS↓ [77] | ||||
| Cinnamaldehyde (CIN) | 40 µM | 3 h | Cyadox 40 µg/mL—3 h | [58] |
| Cellular mechanism Eryptosis inhibited with oxidative stress↓, activity of active caspases 3 and 8↓ and LDH↓ | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Restivo, I.; Attanzio, A.; Tesoriere, L.; Allegra, M.; Garcia-Llatas, G.; Cilla, A. Anti-Eryptotic Activity of Food-Derived Phytochemicals and Natural Compounds. Int. J. Mol. Sci. 2022, 23, 3019. https://doi.org/10.3390/ijms23063019
Restivo I, Attanzio A, Tesoriere L, Allegra M, Garcia-Llatas G, Cilla A. Anti-Eryptotic Activity of Food-Derived Phytochemicals and Natural Compounds. International Journal of Molecular Sciences. 2022; 23(6):3019. https://doi.org/10.3390/ijms23063019
Chicago/Turabian StyleRestivo, Ignazio, Alessandro Attanzio, Luisa Tesoriere, Mario Allegra, Guadalupe Garcia-Llatas, and Antonio Cilla. 2022. "Anti-Eryptotic Activity of Food-Derived Phytochemicals and Natural Compounds" International Journal of Molecular Sciences 23, no. 6: 3019. https://doi.org/10.3390/ijms23063019
APA StyleRestivo, I., Attanzio, A., Tesoriere, L., Allegra, M., Garcia-Llatas, G., & Cilla, A. (2022). Anti-Eryptotic Activity of Food-Derived Phytochemicals and Natural Compounds. International Journal of Molecular Sciences, 23(6), 3019. https://doi.org/10.3390/ijms23063019